Serum and Muscle 1H NMR-Based Metabolomics Profiles Reveal Metabolic Changes Influenced by a Maternal Leucine-Rich Diet in Tumor-Bearing Adult Offspring Rats

 , , , and
, , , and 
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Diets
2.2. Experimental Procedure
2.3. Sample Preparation and 1H NMR Data Acquisition
2.4. Spectral Data Processing and Multivariable Statistical Analysis
3. Results
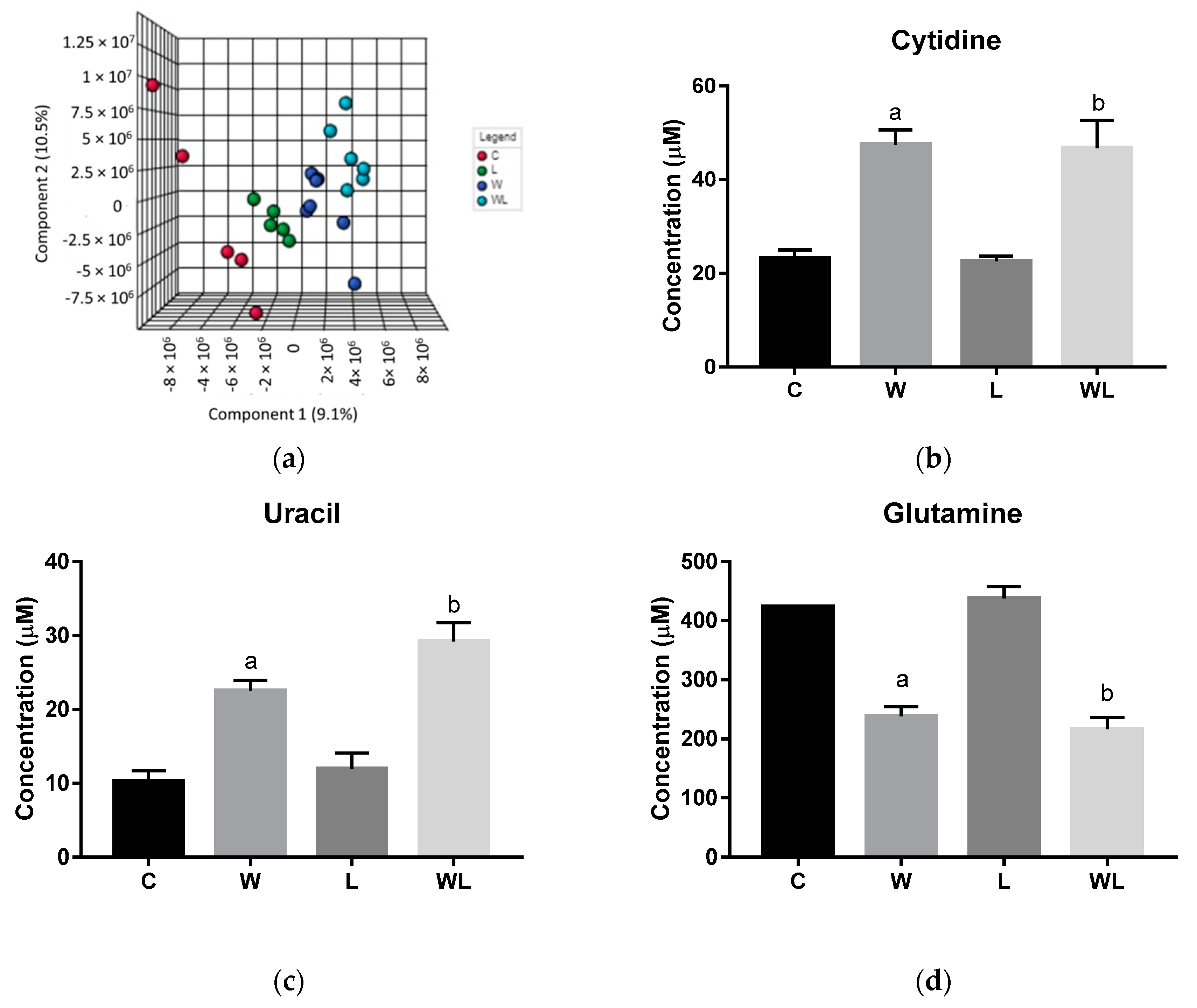
3.1. Initial Spectral Analysis Showed Differences Related to the Walker-256 Tumour Growth and Maternal Leucine-Rich Diet in Serum Samples
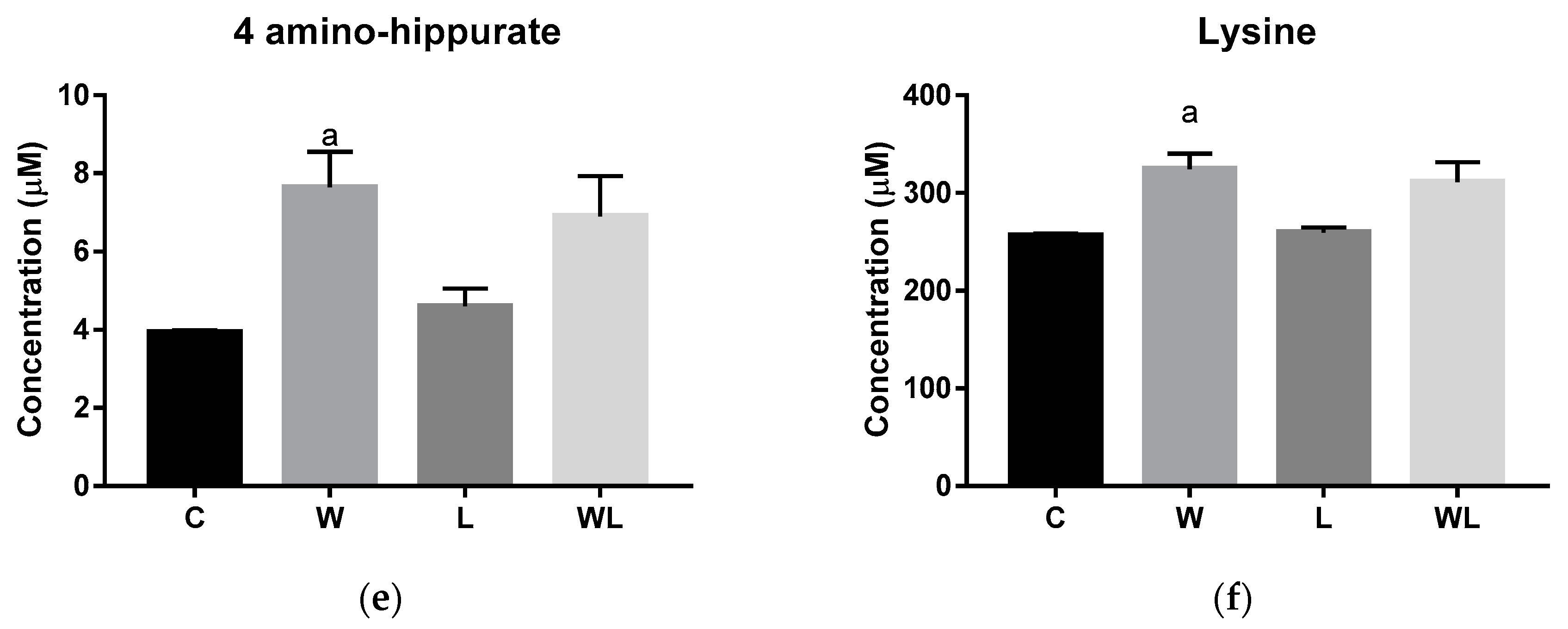
3.2. Serum Chemometrics Analysis Reveals Metabolic Separation between the Groups and the Metabolites Responsible for this Differentiation
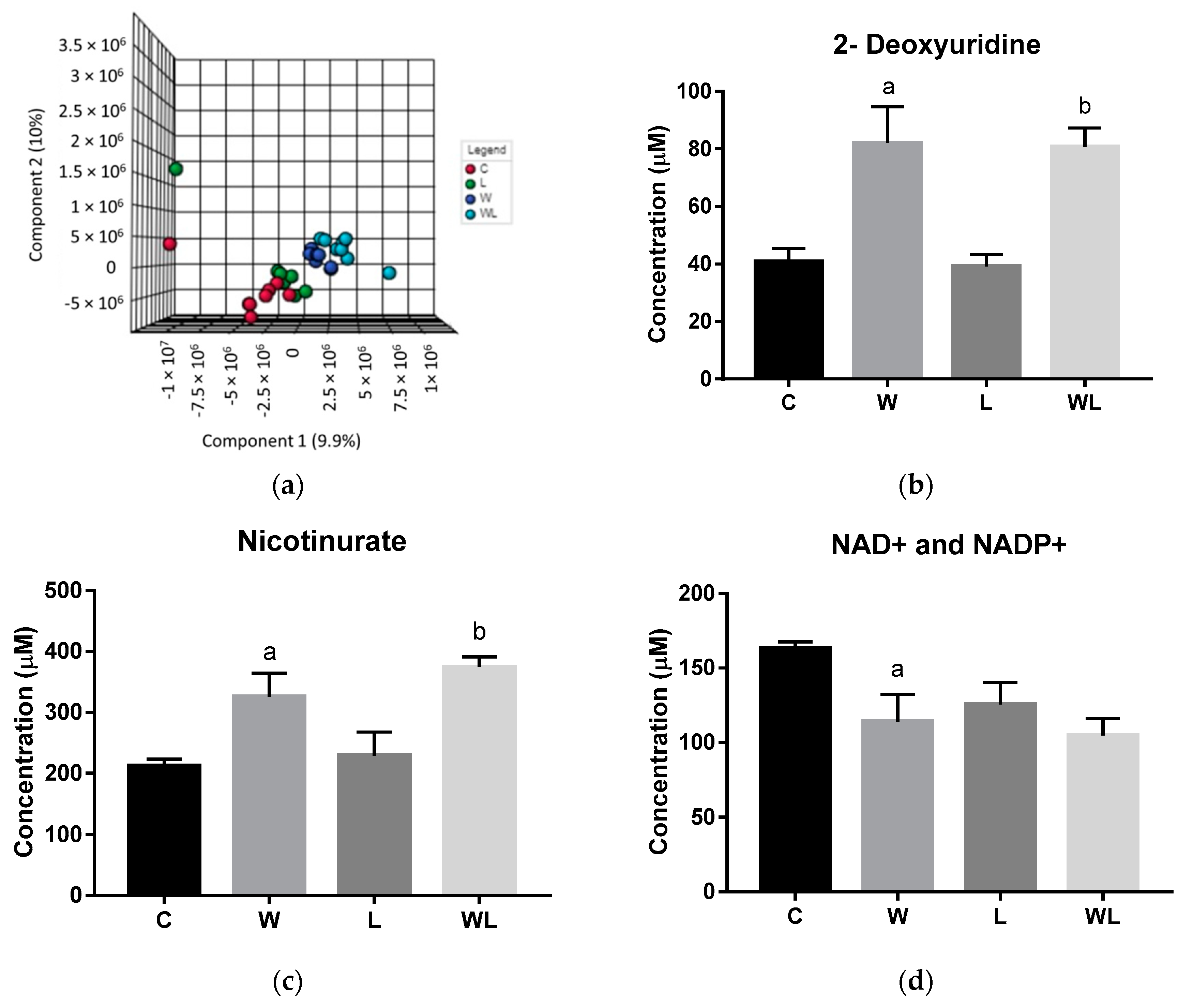
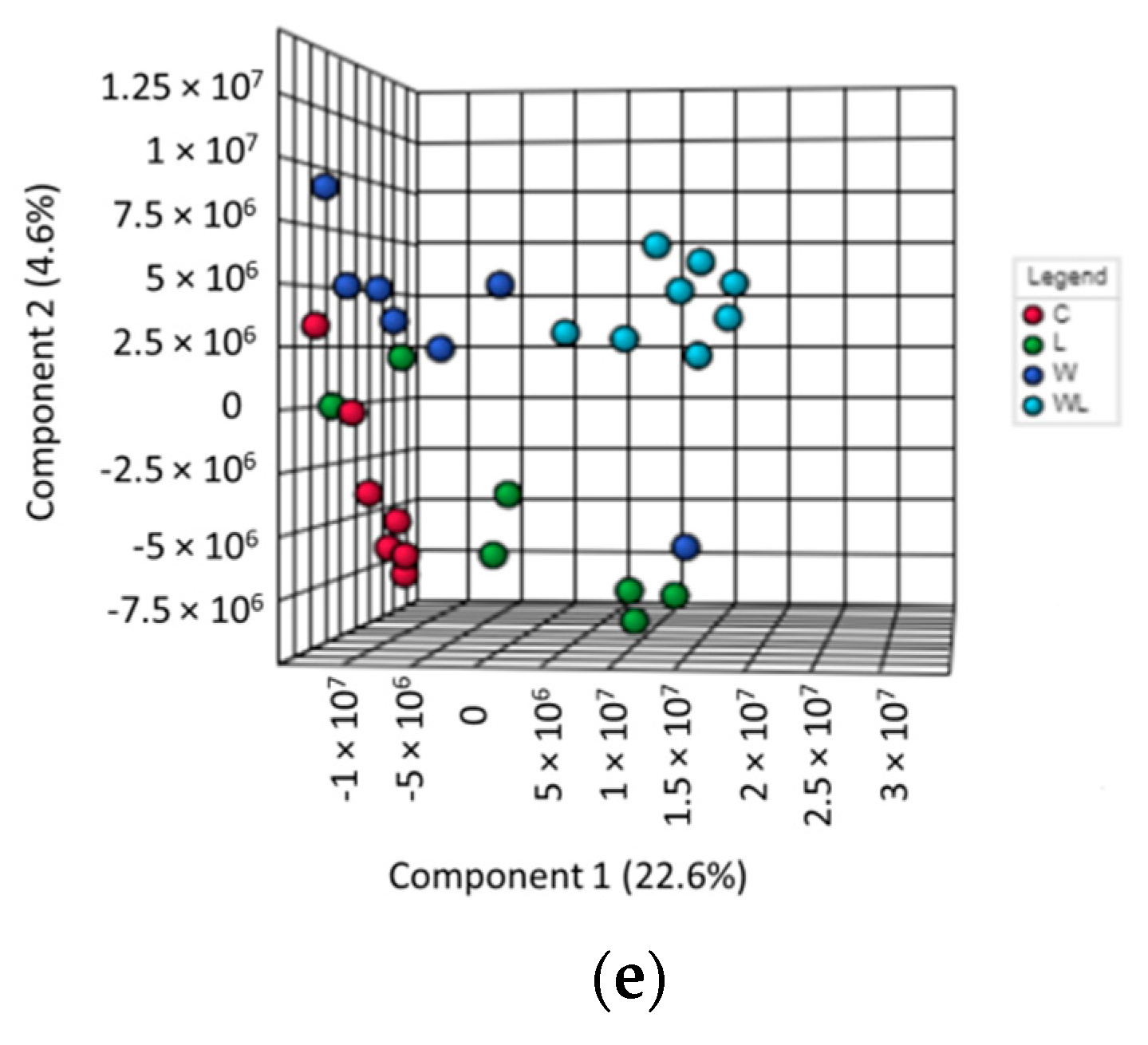
3.3. Initial Spectral Analysis in Gastrocnemius Muscle Showed Differences Related to the Walker-256 Tumour Growth and Maternal Leucine-Rich Diet
3.4. Gastrocnemius Muscle Chemometrics Analysis Reveals the Metabolites Responsible for the Metabolic Segregation between the Tumour-Bearing Groups
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Argilés, J.M.; Stemmler, B.; López-Soriano, F.J.; Busquets, S. Inter-tissue communication in cancer cachexia. Nat. Rev. Endocrinol. 2018. [Google Scholar] [CrossRef]
- Fearon, K.; Strasser, F.; Anker, S.D.; Bosaeus, I.; Bruera, E.; Fainsinger, R.L.; Jatoi, A.; Loprinzi, C.; MacDonald, N.; Mantovani, G.; et al. Definition and classification of cancer cachexia: An international consensus. Lancet Oncol. 2011, 12, 489–495. [Google Scholar] [CrossRef]
- da Silva Miyaguti, N.A.; de Oliveira, S.C.P.; Gomes-Marcondes, M.C.C. Maternal Leucine-Rich Diet Minimises Muscle Mass Loss in Tumour-bearing Adult Rat Offspring by Improving the Balance of Muscle Protein Synthesis and Degradation. Biomolecules 2019, 9, 229. [Google Scholar] [CrossRef] [PubMed]
- Regnault, T.R.H.; de Vrijer, B.; Battaglia, F.C. Transport and Metabolism of Amino Acids in Placenta. Endocrine 2002, 19, 23–41. [Google Scholar] [CrossRef]
- Viana, L.R.; Gomes-Marcondes, M.C.C. Leucine-Rich Diet Improves the Serum Amino Acid Profile and Body Composition of Fetuses from Tumor-Bearing Pregnant Mice1. Biol. Reprod. 2013, 88, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Melnik, B.C. Milk—A Nutrient System of Mammalian Evolution Promoting mTORC1-Dependent Translation. Int. J. Mol. Sci. 2015, 16, 17048–17087. [Google Scholar] [CrossRef]
- Porporato, P.E. Understanding cachexia as a cancer metabolism syndrome. Oncogenesis 2016, 5, e200. [Google Scholar] [CrossRef]
- Saxton, R.A.; Sabatini, D.M. mTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef]
- Costelli, P.; Reffo, P.; Penna, F.; Autelli, R.; Bonelli, G.; Baccino, F.M. Ca(2+)-dependent proteolysis in muscle wasting. Int. J. Biochem. Cell Biol. 2005, 37, 2134–2146. [Google Scholar] [CrossRef]
- Fearon, K.C.H.; Glass, D.J.; Guttridge, D.C. Cancer cachexia: Mediators, signaling, and metabolic pathways. Cell Metab. 2012, 16, 153–166. [Google Scholar] [CrossRef]
- Mathias, P.C.F.; Elmhiri, G.; De Oliveira, J.C.; Delayre-Orthez, C.; Barella, L.F.; Tófolo, L.P.; Fabricio, G.S.; Chango, A.; Abdennebi-Najar, L. Maternal diet, bioactive molecules, and exercising as reprogramming tools of metabolic programming. Eur. J. Nutr. 2014, 53, 711–722. [Google Scholar] [CrossRef] [PubMed]
- Chango, A.; Pogribny, I.P. Considering maternal dietary modulators for epigenetic regulation and programming of the fetal epigenome. Nutrients 2015, 7, 2748–2770. [Google Scholar] [CrossRef] [PubMed]
- Kosmides, A.K.; Kamisoglu, K.; Calvano, S.E.; Corbett, S.A.; Androulakis, I.P. Metabolomic Fingerprinting: Challenges and Opportunities. Crit Rev. Biomed. Eng. 2013, 41, 205–221. [Google Scholar] [CrossRef]
- López, N.; Sánchez, J.; Picó, C.; Palou, A.; Serra, F. Dietary l-leucine supplementation of lactating rats results in a tendency to increase lean/fat ratio associated to lower orexigenic neuropeptide expression in hypothalamus. Peptides 2010, 31, 1361–1367. [Google Scholar] [CrossRef] [PubMed]
- López, N.; Sánchez, J.; Palou, A.; Serra, F. Gender-associated impact of early leucine supplementation on adult predisposition to obesity in rats. Nutrients 2018, 10, 76. [Google Scholar] [CrossRef]
- Wishart, D.S. Emerging applications of metabolomics in drug discovery and precision medicine. Nat. Rev. Drug Discov. 2016, 15, 473–484. [Google Scholar] [CrossRef] [PubMed]
- Gallagher, I.J.; Jacobi, C.; Tardif, N.; Rooyackers, O.; Fearon, K. Omics/systems biology and cancer cachexia. Semin. Cell Dev. Biol. 2016, 54, 92–103. [Google Scholar] [CrossRef] [PubMed]
- Twelkmeyer, B.; Tardif, N.; Rooyackers, O. Omics and cachexia. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 181–185. [Google Scholar] [CrossRef]
- Paul, H.A.; Bomhof, M.R.; Vogel, H.J.; Reimer, R.A. Diet-induced changes in maternal gut microbiota and metabolomic profiles influence programming of offspring obesity risk in rats. Sci. Rep. 2016, 6, 1–14. [Google Scholar] [CrossRef]
- Vale, C.; Stewart, L.; Tierney, J. Trends in UK cancer trials: Results from the UK Coordinating Committee for Cancer Research National Register of Cancer Trials. Br. J. Cancer 2005, 92, 811–814. [Google Scholar] [CrossRef][Green Version]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 Purified Diets for Laboratory Rodents: Final Report of the American Institute of Nutrition Ad Hoc Writing Committee on the Reformulation of the AIN-76A Rodent Diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- da Silva Miyaguti, N.A.; de Oliveira, S.C.P.; Gomes-Marcondes, M.C.C. Maternal nutritional supplementation with fish oil and/or leucine improves hepatic function and antioxidant defenses, and minimizes cachexia indexes in Walker-256 tumor-bearing rats offspring. Nutr. Res. 2018, 51, 29–39. [Google Scholar] [CrossRef] [PubMed]
- Baker, D.E.J. Reproduction and breeding. In The Laboratory Rat; Baker, H.J., Lindsey, J.R., Weisbroth, S.H., Eds.; Academic Press: New York, NY, USA, 1979; Volume 1, pp. 153–168. [Google Scholar]
- Gomes-Marcondes, M.C.C.; Curi, R.; Cury, L. Consequences of Walker 256 tumour growth for the placental/foetal development in rats. J. Cancer Res. Ther. 1998, 5, 277–283. [Google Scholar]
- Viana, L.R.; Canevarolo, R.; Luiz, A.C.P.; Soares, R.F.; Lubaczeuski, C.; de Matto Zeri, A.C.; Gomes-Marcondes, M.C.C. Leucine-rich diet alters the 1H-NMR based metabolomic profile without changing the Walker-256 tumour mass in rats. BMC Cancer 2016, 16. [Google Scholar] [CrossRef] [PubMed]
- Le Belle, J.E.; Harris, N.G.; Williams, S.R.; Bhakoo, K.K. A comparison of cell and tissue extraction techniques using high-resolution 1H-NMR spectroscopy. NMR Biomed. 2002, 15, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Dieterle, F.; Ross, A.; Schlotterbeck, G.; Senn, H. Probabilistic quotient normalization as robust method to account for dilution of complex biological mixtures. Application in 1H NMR metabonomics. Anal. Chem. 2006, 78, 4281–4290. [Google Scholar] [CrossRef]
- Trygg, J.; Holmes, E.; Lundstedt, T. Chemometrics in metabonomics. J. Proteome Res. 2007, 6, 469–479. [Google Scholar] [CrossRef] [PubMed]
- Gad, S.C.; Weil, C.S. Statistics for toxicologists. In Principles and Methods of Toxicology, 2nd ed.; Wallace, H., Ed.; Raven Press Ltd.: New York, NY, USA, 1989; pp. 221–274. [Google Scholar]
- Quan-Jun, Y.; Jun, B.; Li-Li, W.; Yong-Long, H.; Bin, L.; Qi, Y.; Yan, L.; Cheng, G.; Gen-Jin, Y. NMR-based metabolomics reveals distinct pathways mediated by curcumin in cachexia mice bearing CT26 tumor. RSC Adv. 2015, 5, 11766–11775. [Google Scholar] [CrossRef]
- Quanjun, Y.; Genjin, Y.; Lili, W.; Bin, L.; Jin, L.; Qi, Y.; Yan, L.; Yonglong, H.; Cheng, G.; Junping, Z. Serum metabolic profiles reveal the effect of formoterol on cachexia in tumor-bearing mice. Mol. Biosyst. 2013, 9, 3015–3025. [Google Scholar] [CrossRef]
- Vaitheesvaran, B.; Xu, J.; Yee, J.; Lu, Q.-Y.; Go, V.L.; Xiao, G.G.; Lee, W.-N. The Warburg effect: A balance of flux analysis. Metabolomics 2015, 11, 787–796. [Google Scholar] [CrossRef]
- Yu, K.; Matzapetakis, M.; Valent, D.; Saco, Y.; De Almeida, A.M.; Terré, M.; Bassols, A. Skeletal muscle metabolomics and blood biochemistry analysis reveal metabolic changes associated with dietary amino acid supplementation in dairy calves. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Rambla, C.; Puchades-Carrasco, L.; García-Flores, M.; Rubio-Briones, J.; López-Guerrero, J.A.; Pineda-Lucena, A. Non-invasive urinary metabolomic profiling discriminates prostate cancer from benign prostatic hyperplasia. Metabolomics 2017, 13, 1–12. [Google Scholar] [CrossRef]
- Cui, P.; Shao, W.; Huang, C.; Wu, C.; Jiang, B.; Lin, D. Metabolic derangements of skeletal muscle from a murine model of glioma cachexia. Skelet. Muscle 2019, 9, 3. [Google Scholar] [CrossRef]
- Der-Torossian, H.; Wysong, A.; Shadfar, S.; Willis, M.S.; McDunn, J.; Couch, M.E. Metabolic derangements in the gastrocnemius and the effect of Compound A therapy in a murine model of cancer cachexia. J. Cachexia. Sarcopenia Muscle 2013, 4, 145–155. [Google Scholar] [CrossRef]
- Migliavacca, E.; Tay, S.K.H.; Patel, H.P.; Sonntag, T.; Civiletto, G.; McFarlane, C.; Forrester, T.; Barton, S.J.; Leow, M.K.; Antoun, E.; et al. Mitochondrial oxidative capacity and NAD+ biosynthesis are reduced in human sarcopenia across ethnicities. Nat. Commun. 2019, 10. [Google Scholar] [CrossRef]
- Peters, S.J.; van Helvoort, A.; Kegler, D.; Argilès, J.M.; Luiking, Y.C.; Laviano, A.; van Bergenhenegouwen, J.; Deutz, N.E.P.; Haagsman, H.P.; Gorselink, M.; et al. Dose-dependent effects of leucine supplementation on preservation of muscle mass in cancer cachectic mice. Oncol. Rep. 2011, 26, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Frickenschimidt, A.; Frohlich, H.; Bullinger, D.; Zell, A.; Laufer, S.; Gleiter, C.H.; Liebich, H.; Kammaerer, B. Metabonomics in cancer diagnosis: Mass spectrometry-based profiling of urinary nucleosides from breast cancer patients. Biomarkers 2008, 13, 435–449. [Google Scholar] [CrossRef] [PubMed]
- Lo, W.; Jeng, L.; Lai, C.; Tsai, F.; Lin, C.; Chen, W.T. Urinary cytidine as an adjunct biomarker to improve the diagnostic ratio for gastric cancer in Taiwanese patients. Clin. Chim. Acta 2014, 428, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Wu, G. Amino acids: Metabolism, functions, and nutrition. Amino Acids 2009, 37, 1–17. [Google Scholar] [CrossRef]
- Pin, F.; Barreto, R.; Couch, M.E.; Bonetto, A.; O’Connell, T.M. Cachexia induced by cancer and chemotherapy yield distinct perturbations to energy metabolism. J. Cachexia. Sarcopenia Muscle 2019, 10, 140–154. [Google Scholar] [CrossRef] [PubMed]
- O’Connell, T.M.; Ardeshirpour, F.; Asher, S.A.; Winnike, J.H.; Yin, X.; George, J.; Guttridge, D.C.; He, W.; Wysong, A.; Willis, M.S.; et al. Metabolomic analysis of cancer cachexia reveals distinct lipid and glucose alterations. Metabolomics 2008, 4, 216–225. [Google Scholar] [CrossRef]
- Der-Torossian, H.; Asher, S.A.; Winnike, J.H.; Wysong, A.; Yin, X.; Willis, M.S.; O’Connell, T.M.; Couch, M.E. Cancer cachexia’s metabolic signature in a murine model confirms a distinct entity. Metabolomics 2013, 9, 730–739. [Google Scholar] [CrossRef]
- Cala, M.P.; Agulló-Ortuño, M.T.; Prieto-García, E.; González-Riano, C.; Parrilla-Rubio, L.; Barbas, C.; Díaz-García, C.V.; García, A.; Pernaut, C.; Adeva, J.; et al. Multiplatform plasma fingerprinting in cancer cachexia: A pilot observational and translational study. J. Cachexia Sarcopenia Muscle 2018, 9, 348–357. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VIP Score | Metabolites | Chemical Shifts (ppm) |
|---|---|---|
| Serum | ||
 | 4-aminohippurate | 7.63 and 7.66 |
| AMP | 8.25 | |
| Cytidine | 6.01, 6.03 and 6.04 | |
| Glutamine | 2.14 | |
| Lysine | 1.73 and 1.77 | |
| Uracil | 7.52 | |
| Gastrocnemius Polar Extract | ||
 | 2-deoxyuridine | 6.24 and 6.26 |
| Adenosine | 8.26 | |
| Adenosine monophosphate | 8.62 | |
| Anserine | 7.03 | |
| Histidine and Histamine | 7.90 | |
| Isoleucine | 1.08 | |
| NAD+ and NADP+ | 6.03 | |
| Nicotinurate | 8.95 and 8.97 | |
| O-acetylcarnitine | 5.64 | |
| Gastrocnemius Non-Polar Extract | ||
 | Glycerophospholipids | 2.66 and 3.56 |
| Phosphatidylcholine | 4.46 | |
| Polyunsaturated fatty acids | 2.94 and 4.97 | |
| Sphingomyelin | 3.19 | |
| Triacylglycerols | 2.57 | |
| Unsaturated octadecanoic acid |
| Comparisons | Serum Metabolites | Muscle Polar Metabolites | Implications and Future Perspectives | |
|---|---|---|---|---|
| W vs. C | ↑W | Lactate Tyrosine Histidine Phenylalanine Tryptophan Cytidine Uracil 4-aminohippurate Lysine | Creatine 2-deoxyuridine Nicotinurate |
|
| ↓W | Glutamine | Lactate Anserine NAD+ and NADP+ | ||
| L vs. C | ↑L | Lactate Sugar regions Tyrosine Histidine Phenylalanine |
| |
| ↓L | Lactate Creatine | |||
| WL vs. L | ↑WL | Lactate Alanine Tyrosine Cytidine Uracil | Creatine Anserine 2-deoxyuridine Nicotinurate |
|
| ↓WL | Glutamine | Lactate | ||
| =L | 4-aminohippurate Lysine | NAD+ and NADP+ | ||
| WL vs. W | ↑WL | Lactate Sugar regions Tyrosine Histidine Phenylalanine | Lactate Creatine |
|
| ↓WL | Anserine | |||
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyaguti, N.A.d.S.; Stanisic, D.; Oliveira, S.C.P.d.; dos Santos, G.S.; Manhe, B.S.; Tasic, L.; Gomes-Marcondes, M.C.C. Serum and Muscle 1H NMR-Based Metabolomics Profiles Reveal Metabolic Changes Influenced by a Maternal Leucine-Rich Diet in Tumor-Bearing Adult Offspring Rats. Nutrients 2020, 12, 2106. https://doi.org/10.3390/nu12072106
Miyaguti NAdS, Stanisic D, Oliveira SCPd, dos Santos GS, Manhe BS, Tasic L, Gomes-Marcondes MCC. Serum and Muscle 1H NMR-Based Metabolomics Profiles Reveal Metabolic Changes Influenced by a Maternal Leucine-Rich Diet in Tumor-Bearing Adult Offspring Rats. Nutrients. 2020; 12(7):2106. https://doi.org/10.3390/nu12072106
Chicago/Turabian StyleMiyaguti, Natália Angelo da Silva, Danijela Stanisic, Sarah Christine Pereira de Oliveira, Gabriela Sales dos Santos, Beatriz Schincariol Manhe, Ljubica Tasic, and Maria Cristina Cintra Gomes-Marcondes. 2020. "Serum and Muscle 1H NMR-Based Metabolomics Profiles Reveal Metabolic Changes Influenced by a Maternal Leucine-Rich Diet in Tumor-Bearing Adult Offspring Rats" Nutrients 12, no. 7: 2106. https://doi.org/10.3390/nu12072106
APA StyleMiyaguti, N. A. d. S., Stanisic, D., Oliveira, S. C. P. d., dos Santos, G. S., Manhe, B. S., Tasic, L., & Gomes-Marcondes, M. C. C. (2020). Serum and Muscle 1H NMR-Based Metabolomics Profiles Reveal Metabolic Changes Influenced by a Maternal Leucine-Rich Diet in Tumor-Bearing Adult Offspring Rats. Nutrients, 12(7), 2106. https://doi.org/10.3390/nu12072106