Structure and Function of Bovine Whey Derived Oligosaccharides Showing Synbiotic Epithelial Barrier Protective Properties

 and
and
Abstract
1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Bacterial Strains and Growth Conditions
2.3. Preparation of Bovine Milk Oligosaccharide-Enriched Fractions
2.4. Purification of Oligosaccharide Species for Component Isolation and Characterization
2.5. Oligosaccharide Characterization
2.6. Monitoring of Oligosaccharide Profiles
2.7. Cell Culture
2.8. Transepithelial Electrical Resistance Measurements and Cell Integrity
2.9. Statistics
3. Results
3.1. Structural Characterization of Oligosaccharides in MOS and GMOS Preparations
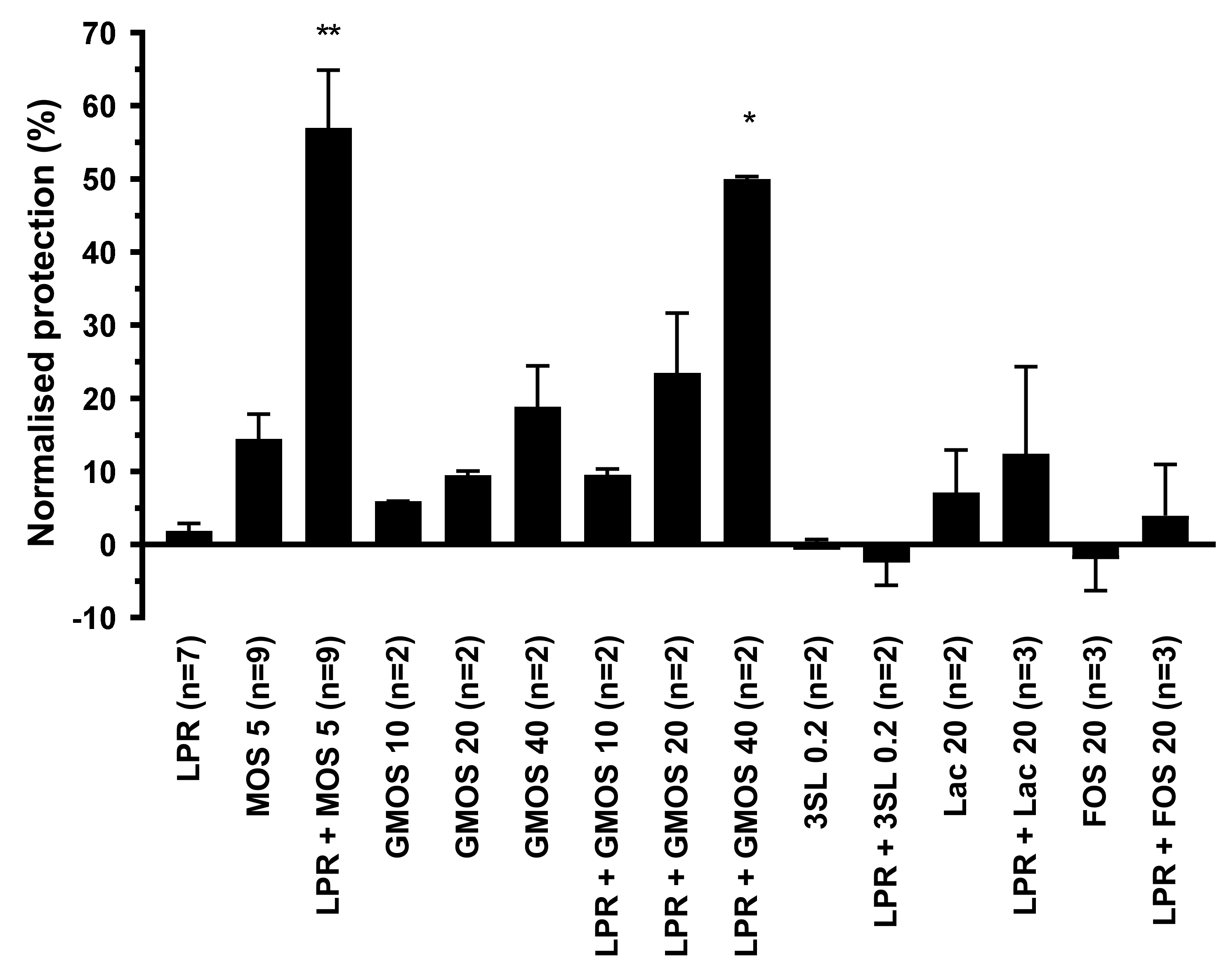
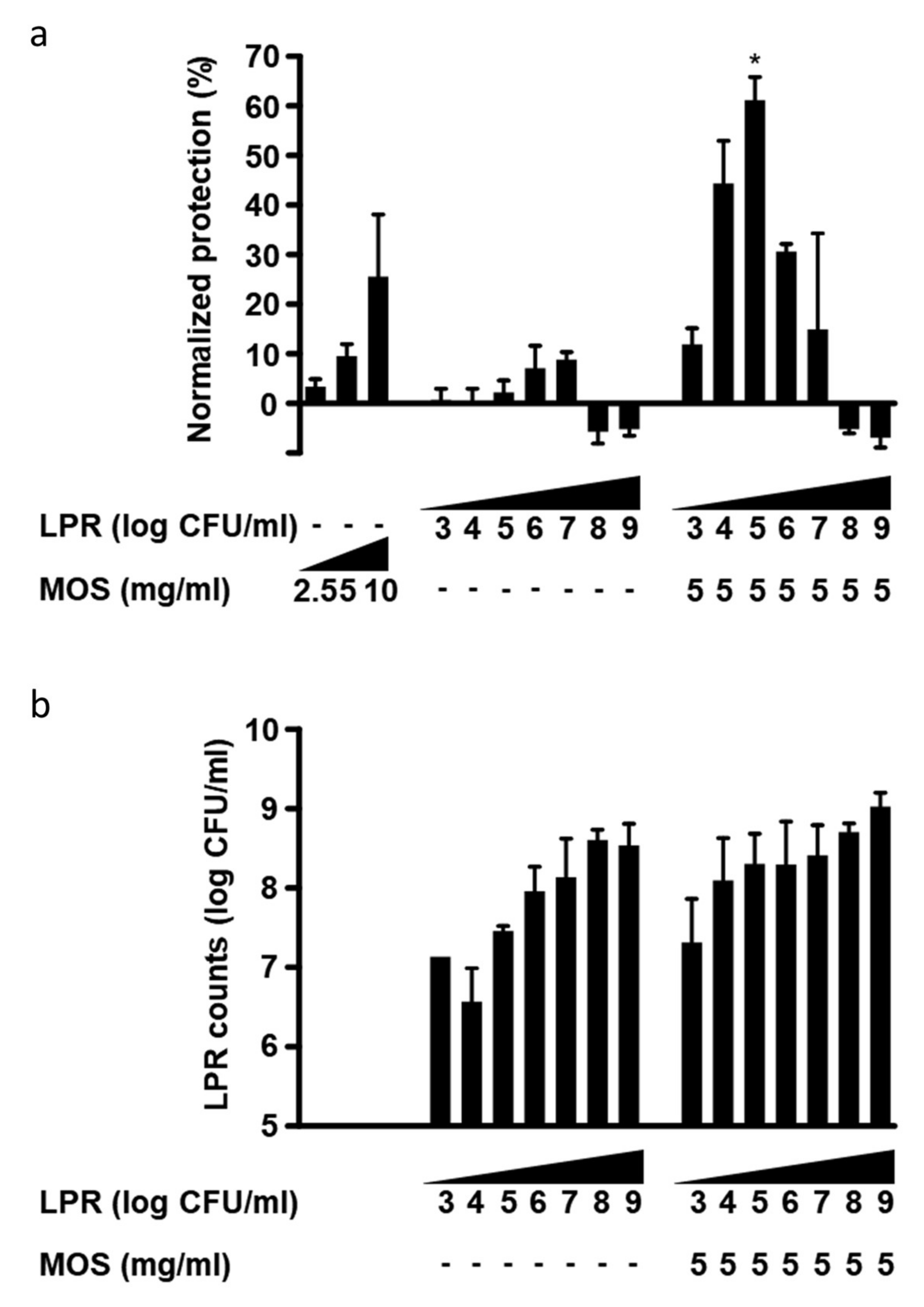
3.2. Both MOS and GMOS Synergize with LPR
3.3. LPR and MOS Synergize to Protect Against Toxin A
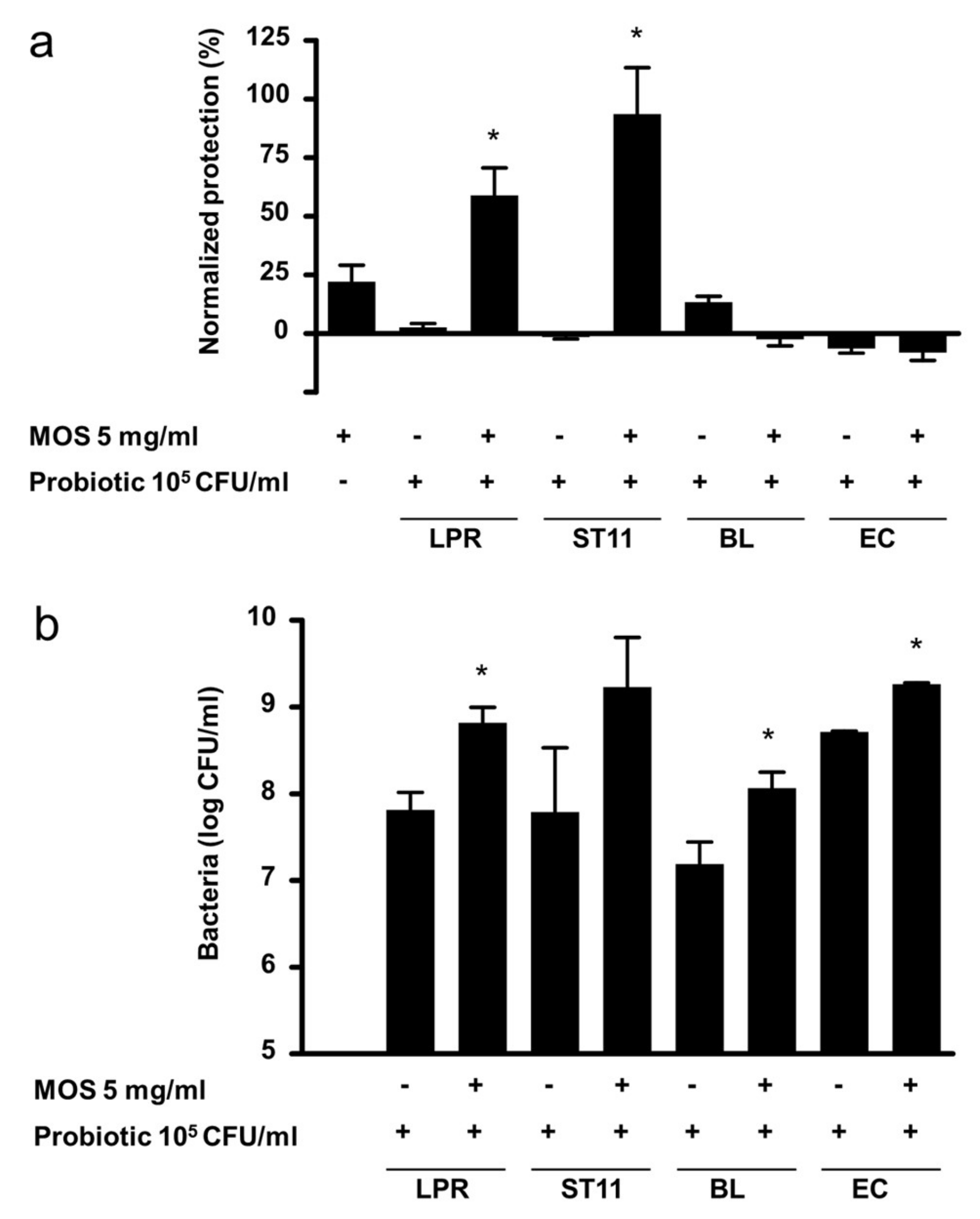
3.4. Specificity of Probiotics
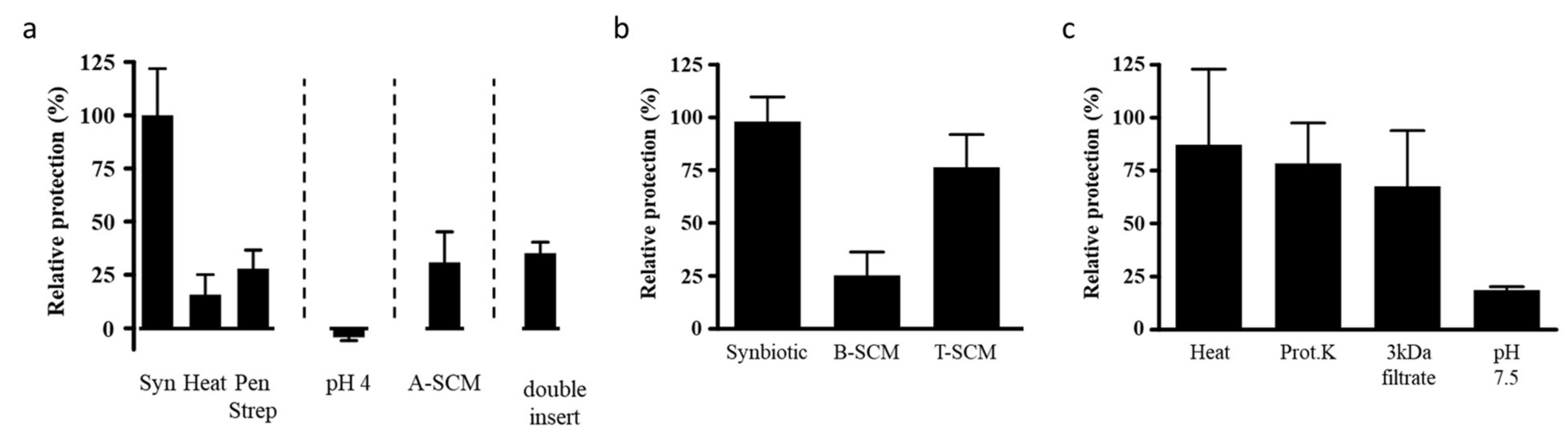
3.5. Characteristics of the Requirements for Protection
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A. Supplementary Description of Structure Identification for Each Fraction
References
- Backhed, F.; Ley, R.E.; Sonnenburg, J.L.; Peterson, D.A.; Gordon, J.I. Host-bacterial mutualism in the human intestine. Science 2005, 307, 1915–1920. [Google Scholar] [CrossRef]
- Geuking, M.B.; Koller, Y.; Rupp, S.; McCoy, K.D. The interplay between the gut microbiota and the immune system. Gut Microbes 2014, 5, 411–418. [Google Scholar] [CrossRef] [PubMed]
- Glendinning, L.; Free, A. Supra-organismal interactions in the human intestine. Front. Cell. Infect. Microbiol. 2014, 4, 47. [Google Scholar] [CrossRef]
- Illiano, P.; Brambilla, R.; Parolini, C. The mutual interplay of gut microbiota, diet and human disease. FEBS J. 2020, 287, 833–855. [Google Scholar] [CrossRef] [PubMed]
- Rooks, M.G.; Garrett, W.S. Gut microbiota, metabolites and host immunity. Nat. Rev. Immunol. 2016, 16, 341–352. [Google Scholar] [CrossRef]
- Soderholm, A.T.; Pedicord, V.A. Intestinal epithelial cells: At the interface of the microbiota and mucosal immunity. Immunology 2019, 158, 267–280. [Google Scholar] [CrossRef]
- Chu, H.; Mazmanian, S.K. Innate immune recognition of the microbiota promotes host-microbial symbiosis. Nat. Immunol. 2013, 14, 668–675. [Google Scholar] [CrossRef]
- Sellge, G.; Kufer, T.A. PRR-signaling pathways: Learning from microbial tactics. Semin. Immunol. 2015, 27, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Blacher, E.; Levy, M.; Tatirovsky, E.; Elinav, E. Microbiome-modulated metabolites at the interface of host immunity. J. Immunol. 2017, 198, 572–580. [Google Scholar] [CrossRef] [PubMed]
- Haase, S.; Haghikia, A.; Wilck, N.; Muller, D.N.; Linker, R.A. Impacts of microbiome metabolites on immune regulation and autoimmunity. Immunology 2018, 154, 230–238. [Google Scholar] [CrossRef] [PubMed]
- Hills, R.D., Jr.; Pontefract, B.A.; Mishcon, H.R.; Black, C.A.; Sutton, S.C.; Theberge, C.R. Gut microbiome: Profound implications for diet and disease. Nutrients 2019, 11, 1613. [Google Scholar] [CrossRef] [PubMed]
- Stinson, L.F. Establishment of the early-life microbiome: A dohad perspective. J. Dev. Orig. Health Dis. 2019, 1–10, 201–210. [Google Scholar] [CrossRef] [PubMed]
- Floch, M.H.; Ringel, Y.; Walker, W.A. The Microbiota in Gastrointestinal Pathophysiology: Implications for Human Health, Prebiotics, Probiotics, and Dysbiosis; Academic Press: Cambridge, MA, USA, 2016. [Google Scholar]
- Sanders, M.E.; Merenstein, D.J.; Reid, G.; Gibson, G.R.; Rastall, R.A. Probiotics and prebiotics in intestinal health and disease: From biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 2019, 16, 605–616. [Google Scholar] [CrossRef] [PubMed]
- Asahara, T.; Nomoto, K.; Shimizu, K.; Watanuki, M.; Tanaka, R. Increased resistance of mice to salmonella enterica serovar typhimurium infection by synbiotic administration of bifidobacteria and transgalactosylated oligosaccharides. J. Appl. Microbiol. 2001, 91, 985–996. [Google Scholar] [CrossRef]
- Childs, C.E.; Roytio, H.; Alhoniemi, E.; Fekete, A.A.; Forssten, S.D.; Hudjec, N.; Lim, Y.N.; Steger, C.J.; Yaqoob, P.; Tuohy, K.M.; et al. Xylo-oligosaccharides alone or in synbiotic combination with bifidobacterium animalis subsp. Lactis induce bifidogenesis and modulate markers of immune function in healthy adults: A double-blind, placebo-controlled, randomised, factorial cross-over study. Br. J. Nutr. 2014, 111, 1945–1956. [Google Scholar] [CrossRef]
- Le Leu, R.K.; Hu, Y.; Brown, I.L.; Woodman, R.J.; Young, G.P. Synbiotic intervention of bifidobacterium lactis and resistant starch protects against colorectal cancer development in rats. Carcinogenesis 2010, 31, 246–251. [Google Scholar] [CrossRef]
- Roller, M.; Pietro Femia, A.; Caderni, G.; Rechkemmer, G.; Watzl, B. Intestinal immunity of rats with colon cancer is modulated by oligofructose-enriched inulin combined with lactobacillus rhamnosus and bifidobacterium lactis. Br. J. Nutr. 2004, 92, 931–938. [Google Scholar] [CrossRef]
- Umeki, M.; Oue, K.; Mochizuki, S.; Shirai, Y.; Sakai, K. Effect of lactobacillus rhamnosus ky-3 and cellobiose as synbiotics on lipid metabolism in rats. J. Nutr. Sci. Vitam. 2004, 50, 330–334. [Google Scholar] [CrossRef]
- Dilli, D.; Aydin, B.; Fettah, N.D.; Ozyazici, E.; Beken, S.; Zenciroglu, A.; Okumus, N.; Ozyurt, B.M.; Ipek, M.S.; Akdag, A.; et al. The propre-save study: Effects of probiotics and prebiotics alone or combined on necrotizing enterocolitis in very low birth weight infants. J. Pediatr. 2015, 166, 545–551.e541. [Google Scholar] [CrossRef]
- Roller, M.; Rechkemmer, G.; Watzl, B. Prebiotic inulin enriched with oligofructose in combination with the probiotics lactobacillus rhamnosus and bifidobacterium lactis modulates intestinal immune functions in rats. J. Nutr. 2004, 134, 153–156. [Google Scholar] [CrossRef]
- Pothoulakis, C.; Lamont, J.T. Microbes and microbial toxins: Paradigms for microbial-mucosal interactions II. The integrated response of the intestine to clostridium difficile toxins. Am. J. Physiol. Gastrointest. Liver Physiol. 2001, 280, G178–G183. [Google Scholar] [CrossRef] [PubMed]
- Padua, D.; Pothoulakis, C. Novel approaches to treating clostridium difficile-associated colitis. Expert Rev. Gastroenterol. Hepatol. 2016, 10, 193–204. [Google Scholar] [CrossRef] [PubMed]
- Vincent, Y.; Manji, A.; Gregory-Miller, K.; Lee, C. A review of management of clostridium difficile infection: Primary and recurrence. Antibiotics 2015, 4, 411–423. [Google Scholar] [CrossRef]
- Goldenberg, J.Z.; Yap, C.; Lytvyn, L.; Lo, C.K.; Beardsley, J.; Mertz, D.; Johnston, B.C. Probiotics for the prevention of clostridium difficile-associated diarrhea in adults and children. Cochrane Database Syst. Rev. 2017, 12, CD006095. [Google Scholar] [CrossRef] [PubMed]
- Madoff, S.E.; Urquiaga, M.; Alonso, C.D.; Kelly, C.P. Prevention of recurrent clostridioides difficile infection: A systematic review of randomized controlled trials. Anaerobe 2020, 61, 102098. [Google Scholar] [CrossRef]
- McFarland, L.V. Probiotics for the primary and secondary prevention of C. Difficile infections: A meta-analysis and systematic review. Antibiotics 2015, 4, 160–178. [Google Scholar] [CrossRef]
- Szajewska, H.; Kolodziej, M. Systematic review with meta-analysis: Saccharomyces boulardii in the prevention of antibiotic-associated diarrhoea. Aliment. Pharmacol. Ther. 2015, 42, 793–801. [Google Scholar] [CrossRef]
- Vernaya, M.; McAdam, J.; Hampton, M.D. Effectiveness of probiotics in reducing the incidence of clostridium difficile-associated diarrhea in elderly patients: A systematic review. JBI Database Syst. Rev. Implement. Rep. 2017, 15, 140–164. [Google Scholar] [CrossRef]
- Euler, A.R.; Mitchell, D.K.; Kline, R.; Pickering, L.K. Prebiotic effect of fructo-oligosaccharide supplemented term infant formula at two concentrations compared with unsupplemented formula and human milk. J. Pediatr. Gastroenterol. Nutr. 2005, 40, 157–164. [Google Scholar] [CrossRef]
- Lewis, S.; Burmeister, S.; Cohen, S.; Brazier, J.; Awasthi, A. Failure of dietary oligofructose to prevent antibiotic-associated diarrhoea. Aliment. Pharmacol. Ther. 2005, 21, 469–477. [Google Scholar] [CrossRef]
- Smilowitz, J.T.; Lemay, D.G.; Kalanetra, K.M.; Chin, E.L.; Zivkovic, A.M.; Breck, M.A.; German, J.B.; Mills, D.A.; Slupsky, C.; Barile, D. Tolerability and safety of the intake of bovine milk oligosaccharides extracted from cheese whey in healthy human adults. J. Nutr. Sci. 2017, 6, 6. [Google Scholar] [CrossRef] [PubMed]
- Urashima, T.; Saito, T.; Nakamura, T.; Messer, M. Oligosaccharides of milk and colostrum in non-human mammals. Glycoconj. J. 2001, 18, 357–371. [Google Scholar] [CrossRef] [PubMed]
- Zivkovic, A.M.; Barile, D. Bovine milk as a source of functional oligosaccharides for improving human health. Adv. Nutr. 2011, 2, 284–289. [Google Scholar] [CrossRef] [PubMed]
- Rastall, R.A. Functional oligosaccharides: Application and manufacture. Annu. Rev. Food Sci. Technol. 2010, 1, 305–339. [Google Scholar] [CrossRef] [PubMed]
- Sako, T.; Matsumoto, K.; Tanaka, R. Recent progress on research and applications of non-digestible galacto-oligosaccharides. Int. Dairy J. 1999, 9, 69–80. [Google Scholar] [CrossRef]
- Coulier, L.; Timmermans, J.; Bas, R.; Van Den Dool, R.; Haaksman, I.; Klarenbeek, B.; Slaghek, T.; Van Dongen, W. In-depth characterization of prebiotic galacto-oligosaccharides by a Combination of Analytical Techniques. J. Agric. Food Chem. 2009, 57, 8488–8495. [Google Scholar] [CrossRef]
- Yanahira, S.; Kobayashi, T.; Suguri, T.; Nakakoshi, M.; Miura, S.; Ishikawa, H.; Nakajima, I. Formation of oligosaccharides from lactose by bacillus circulans beta-galactosidase. Biosci. Biotechnol. Biochem. 1995, 59, 1021–1026. [Google Scholar] [CrossRef]
- Van Leeuwen, S.S.; Kuipers, B.J.H.; Dijkhuizen, L.; Kamerling, J.P. (1)H NMR Analysis of the lactose/β-galactosidase-derived Galacto-Oligosaccharide Components of Vivinal® GOS Up to DP5. Carbohydr. Res. 2014, 400, 59–73. [Google Scholar] [CrossRef]
- Austin, S.; Benet, T.; Michaud, J.; Cuany, D.; Rohfritsch, P. Determination of beta -galactooligosaccharides by liquid chromatography. Int. J. Anal. Chem. 2014, 2014, 768406. [Google Scholar] [CrossRef]
- Dumortier, V.; Montreuil, J.; Bouquelet, S. Primary structure of ten galactosides formed by transglycosylation during lactose hydrolysis by bifidobacterium bifidum. Carbohydr. Res. 1990, 201, 115–123. [Google Scholar] [CrossRef]
- Chen, S.; Wei, D.; Hu, Z. Synthesis of galacto-oligosaccharides by immobilized Bacillus stearothermophilus. Wei Sheng Wu Xue Bao 2001, 41, 357–362. [Google Scholar]
- Drakoularakou, A.; Tzortzis, G.; Rastall, R.A.; Gibson, G.R. A double-blind, placebo-controlled, randomized human study assessing the capacity of a novel galacto-oligosaccharide mixture in reducing travellers’ diarrhoea. Eur. J. Clin. Nutr. 2010, 64, 146–152. [Google Scholar] [CrossRef] [PubMed]
- Krumbeck, J.A.; Maldonado-Gomez, M.X.; Martinez, I.; Frese, S.A.; Burkey, T.E.; Rasineni, K.; Ramer-Tait, A.E.; Harris, E.N.; Hutkins, R.W.; Walter, J. In Vivo selection to identify bacterial strains with enhanced ecological performance in synbiotic applications. Appl. Environ. Microbiol. 2015, 81, 2455–2465. [Google Scholar] [CrossRef]
- Krumbeck, J.A.; Rasmussen, H.E.; Hutkins, R.W.; Clarke, J.; Shawron, K.; Keshavarzian, A.; Walter, J. Probiotic bifidobacterium strains and galactooligosaccharides improve intestinal barrier function in obese adults but show no synergism when used together as synbiotics. Microbiome 2018, 6, 121. [Google Scholar] [CrossRef] [PubMed]
- Simeoni, U.; Berger, B.; Junick, J.; Blaut, M.; Pecquet, S.; Rezzonico, E.; Grathwohl, D.; Sprenger, N.; Brussow, H.; Szajewska, H.; et al. Gut microbiota analysis reveals a marked shift to bifidobacteria by a starter infant formula containing a synbiotic of bovine milk-derived oligosaccharides and bifidobacterium animalis subsp. Lactis CNCM I-3446. Environ. Microbiol. 2016, 18, 2185–2195. [Google Scholar] [CrossRef]
- So, D.; Whelan, K.; Rossi, M.; Morrison, M.; Holtmann, G.; Kelly, J.T.; Shanahan, E.R.; Staudacher, H.M.; Campbell, K.L. Dietary fiber intervention on gut microbiota composition in healthy adults: A systematic review and meta-analysis. Am. J. Clin. Nutr. 2018, 107, 965–983. [Google Scholar] [CrossRef] [PubMed]
- Swanson, K.S.; De Vos, W.M.; Martens, E.C.; Gilbert, J.A.; Menon, R.S.; Soto-Vaca, A.; Hautvast, J.; Meyer, P.D.; Borewicz, K.; Vaughan, E.E.; et al. Effect of fructans, prebiotics and fibres on the human gut microbiome assessed by 16s rrna-based approaches: A review. Benef. Microbes 2020, 11, 101–129. [Google Scholar] [CrossRef]
- Barboza, M.; Sela, D.A.; Pirim, C.; Locascio, R.G.; Freeman, S.L.; German, J.B.; Mills, D.A.; Lebrilla, C.B. Glycoprofiling bifidobacterial consumption of galacto-oligosaccharides by mass spectrometry reveals strain-specific, preferential consumption of glycans. Appl. Environ. Microbiol. 2009, 75, 7319–7325. [Google Scholar] [CrossRef] [PubMed]
- Tzortzis, G.; Goulas, A.K.; Gibson, G.R. Synthesis of prebiotic galactooligosaccharides using whole cells of a novel strain, bifidobacterium bifidum ncimb 41171. Appl. Microbiol. Biotechnol. 2005, 68, 412–416. [Google Scholar] [CrossRef] [PubMed]
- Tzortzis, G.; Goulas, A.K.; Gee, J.M.; Gibson, G.R. A novel galactooligosaccharide mixture increases the bifidobacterial population numbers in a continuous in vitro fermentation system and in the proximal colonic contents of pigs in vivo. J. Nutr. 2005, 135, 1726–1731. [Google Scholar] [CrossRef] [PubMed]
- Di Bella, S.; Ascenzi, P.; Siarakas, S.; Petrosillo, N.; Di Masi, A. Clostridium difficile toxins a and b: Insights into pathogenic properties and extraintestinal effects. Toxins 2016, 8, 134. [Google Scholar] [CrossRef] [PubMed]
- Tao, Y.; Drabik, K.A.; Waypa, T.S.; Musch, M.W.; Alverdy, J.C.; Schneewind, O.; Chang, E.B.; Petrof, E.O. Soluble factors from lactobacillus gg activate mapks and induce cytoprotective heat shock proteins in intestinal epithelial cells. Am. J. Physiol. Cell Physiol. 2006, 290, C1018–C1030. [Google Scholar] [CrossRef]
- Liu, T.S.; Musch, M.W.; Sugi, K.; Walsh-Reitz, M.M.; Ropeleski, M.J.; Hendrickson, B.A.; Pothoulakis, C.; Lamont, J.T.; Chang, E.B. Protective role of hsp72 against clostridium difficile toxin a-induced intestinal epithelial cell dysfunction. Am. J. Physiol. Cell Physiol. 2003, 284, C1073–C1082. [Google Scholar] [CrossRef]
- Musch, M.W.; Kaplan, B.; Chang, E.B. Role of increased basal expression of heat shock protein 72 in colonic epithelial c2BBE adenocarcinoma cells. Cell Growth Differ. 2001, 12, 419–426. [Google Scholar] [PubMed]
- Yan, F.; Polk, D.B. Probiotic bacterium prevents cytokine-induced apoptosis in intestinal epithelial cells. J. Biol. Chem. 2002, 277, 50959–50965. [Google Scholar] [CrossRef] [PubMed]
- Yan, F.; Polk, D.B. Characterization of a probiotic-derived soluble protein which reveals a mechanism of preventive and treatment effects of probiotics on intestinal inflammatory diseases. Gut Microbes 2012, 3, 25–28. [Google Scholar] [CrossRef]
- Yan, F.; Polk, D.B. Lactobacillus Rhamnosus GG: An updated strategy to use microbial products to promote health. Funct. Food Rev. (Print) 2012, 4, 77–84. [Google Scholar]
- Chen, X.; Kokkotou, E.G.; Mustafa, N.; Bhaskar, K.R.; Sougioultzis, S.; O’Brien, M.; Pothoulakis, C.; Kelly, C.P. Saccharomyces boulardii inhibits ERK1/2 mitogen-activated protein kinase activation both in vitro and in vivo and protects against clostridium difficile toxin A-induced enteritis. J. Biol. Chem. 2006, 281, 24449–24454. [Google Scholar] [CrossRef]
- Duncan, P.I.; Fotopoulos, G.; Pasche, E.; Porta, N.; Masserey Elmelegy, I.; Sanchez-Garcia, J.L.; Bergonzelli, G.E.; Corthesy-Theulaz, I. Yeast, beef and pork extracts counteract clostridium difficile toxin a enterotoxicity. FEMS Microbiol. Lett. 2009, 295, 218–225. [Google Scholar] [CrossRef]
- Na, X.; Kim, H.; Moyer, M.P.; Pothoulakis, C.; LaMont, J.T. Gp96 is a human colonocyte plasma membrane binding protein for clostridium difficile toxin a. Infect. Immun. 2008, 76, 2862–2871. [Google Scholar] [CrossRef]
- Akashi-Takamura, S.; Miyake, K. TLR accessory molecules. Curr. Opin. Immunol. 2008, 20, 420–425. [Google Scholar] [CrossRef] [PubMed]
- Yang, F.; Wang, J.; Li, X.; Ying, T.; Qiao, S.; Li, D.; Wu, G. 2-DE and MS analysis of interactions between Lactobacillus fermentum I5007 and intestinal epithelial cells. Electrophoresis 2007, 28, 4330–4339. [Google Scholar] [CrossRef] [PubMed]
- Segers, M.E.; Lebeer, S. Towards a better understanding of lactobacillus rhamnosus GG—Host interactions. Microb. Cell Factories 2014, 13, S7. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Collected Fraction | Identified Structure |
|---|---|
| IV-2 | Galβ4Glc |
| IV-1 | Galβ6Gal |
| IV-3/III-1 | Galβ6Galβ4Glc |
| IV-4/III-4 | Galβ3Galβ3Glc |
| IV-5/III-5/II-3 | Galβ3Galβ4Glc |
| III-1b | GalNAcα3Galβ4Glc |
| III-3/II-2 | Galβ6Galβ6Galβ4Glc |
| II-4 | Galβ3Galβ6Galβ4Glc |
| II-5 | Galβ6Galβ3Galβ4Glc |
| II-6 | Galβ3Galβ3Galβ4Glc |
| II-1 | Galβ6GalNAcα3Galβ4Glc |
| III-2 | Galβ6Galβ6Galβ6Glc |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duncan, P.I.; Aitio, O.; Heiskanen, A.; Niemelä, R.; Saarinen, J.; Helin, J.; Porta, N.; Fiaux, M.; Moënnoz, D.; Golliard, M.; et al. Structure and Function of Bovine Whey Derived Oligosaccharides Showing Synbiotic Epithelial Barrier Protective Properties. Nutrients 2020, 12, 2007. https://doi.org/10.3390/nu12072007
Duncan PI, Aitio O, Heiskanen A, Niemelä R, Saarinen J, Helin J, Porta N, Fiaux M, Moënnoz D, Golliard M, et al. Structure and Function of Bovine Whey Derived Oligosaccharides Showing Synbiotic Epithelial Barrier Protective Properties. Nutrients. 2020; 12(7):2007. https://doi.org/10.3390/nu12072007
Chicago/Turabian StyleDuncan, Peter I., Olli Aitio, Annamari Heiskanen, Ritva Niemelä, Juhani Saarinen, Jari Helin, Nadine Porta, Muriel Fiaux, Denis Moënnoz, Mireille Golliard, and et al. 2020. "Structure and Function of Bovine Whey Derived Oligosaccharides Showing Synbiotic Epithelial Barrier Protective Properties" Nutrients 12, no. 7: 2007. https://doi.org/10.3390/nu12072007
APA StyleDuncan, P. I., Aitio, O., Heiskanen, A., Niemelä, R., Saarinen, J., Helin, J., Porta, N., Fiaux, M., Moënnoz, D., Golliard, M., Cherbut, C., Berrocal, R., Austin, S., & Sprenger, N. (2020). Structure and Function of Bovine Whey Derived Oligosaccharides Showing Synbiotic Epithelial Barrier Protective Properties. Nutrients, 12(7), 2007. https://doi.org/10.3390/nu12072007