The Effect of a 2-Week Red Ginseng Supplementation on Food Efficiency and Energy Metabolism in Mice

Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Treatment
2.2. RG Extract
2.3. Energy Expenditure
2.4. Blood Analysis
2.5. Glycogen Analysis
2.6. RT-PCR (Reverse Transcriptase PCR) Analysis
2.7. Statistical Analysis
3. Results
3.1. Body Weight, Food Intake, and Abdominal Tissue Weight
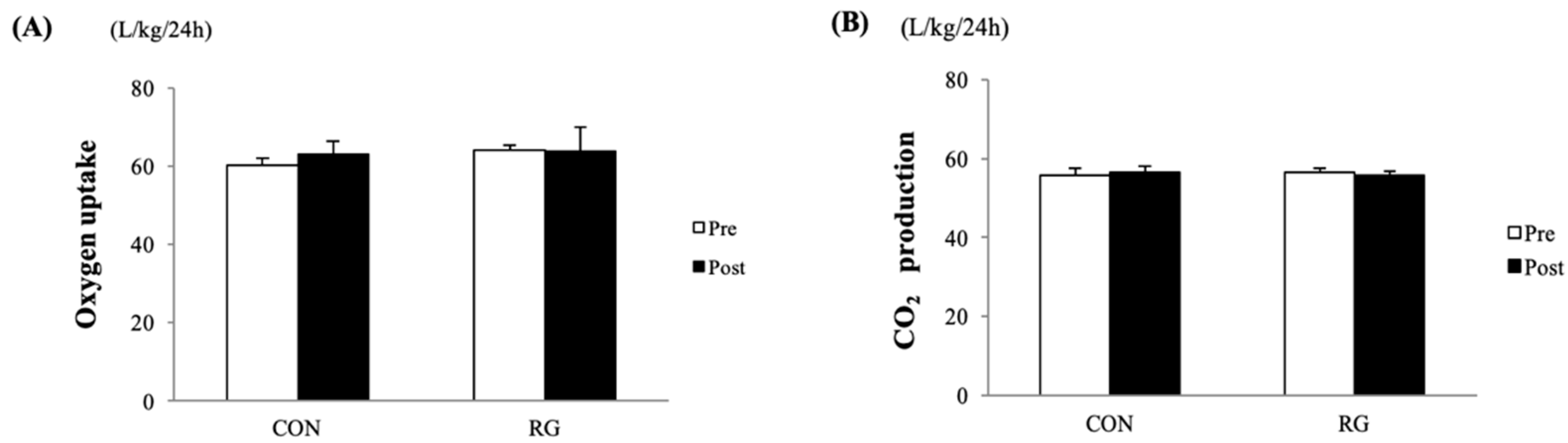
3.2. Oxygen Uptake, CO2 Production, and RMR
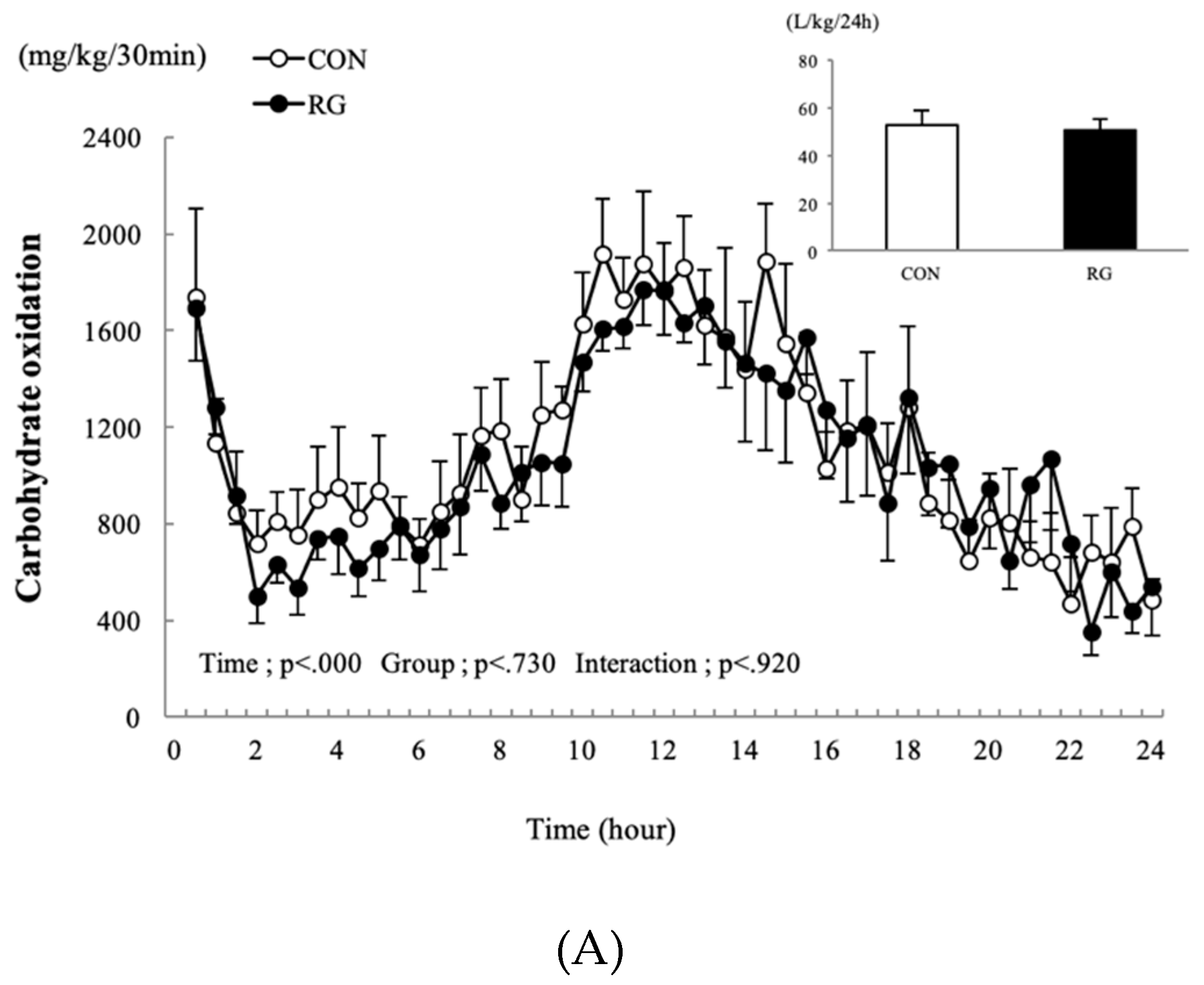
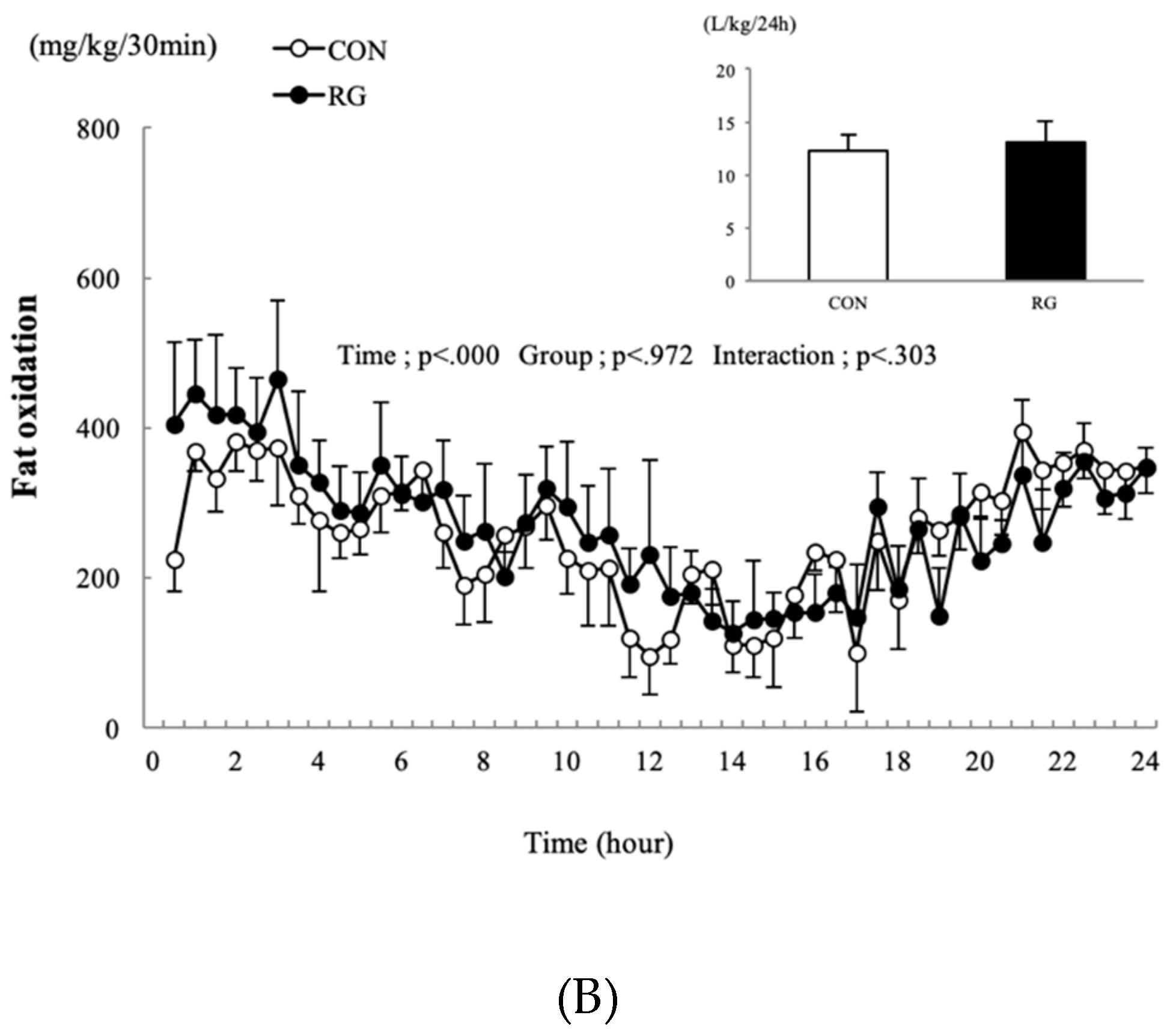
3.3. Carbohydrate and Fat Oxidation over 24 h
3.4. Plasma Parameters
3.5. Glycogen Concentrations
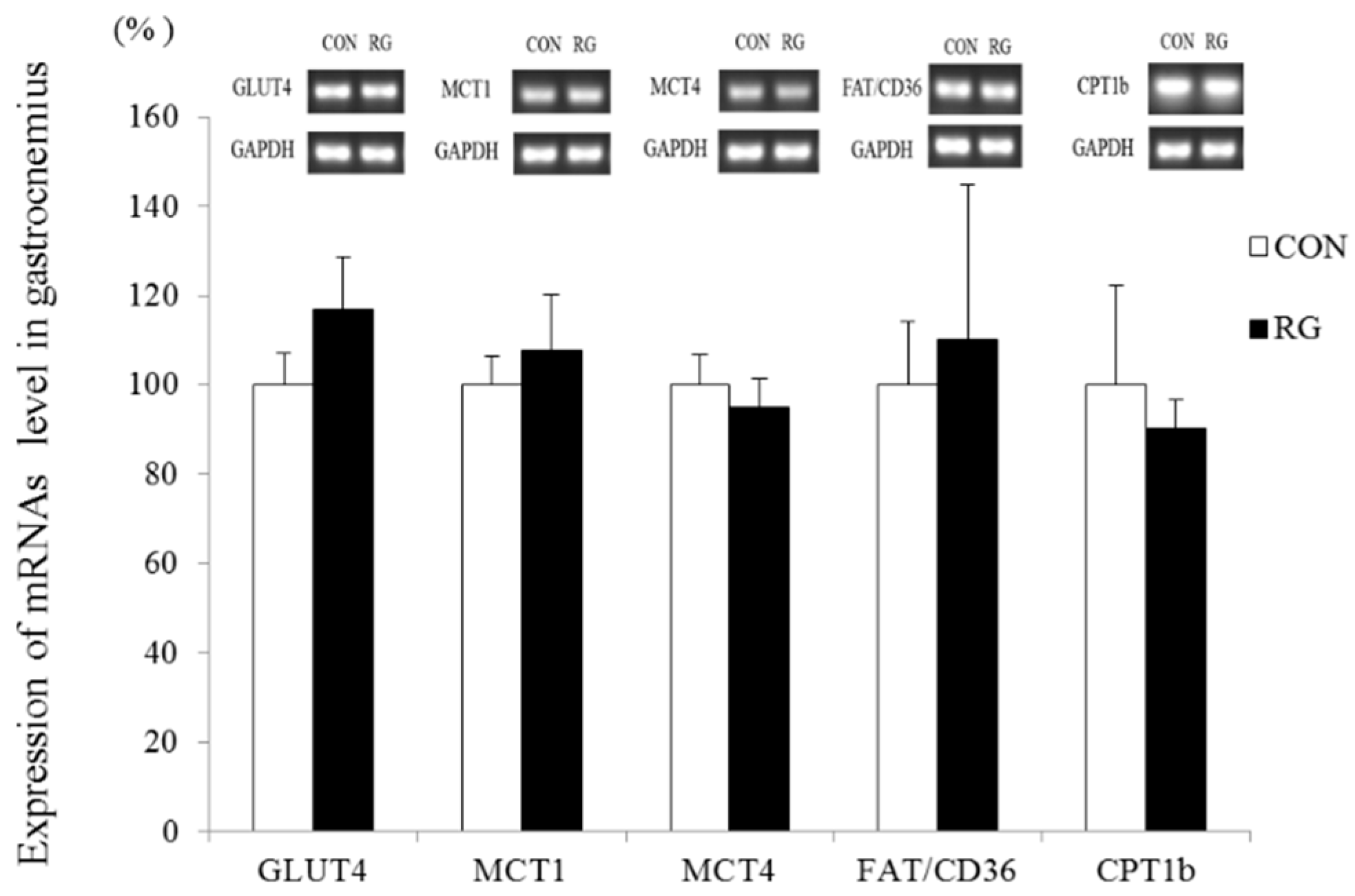
3.6. Expression of Skeletal Muscle mRNAs Related to Energy Metabolism
4. Discussion
4.1. Effect of RG on Total Food Intake and Mean Body Weight
4.2. Effect on Oxygen Uptake and Oxidation of Carbohydrates and Fats
4.3. Effect on Expression of mRNAs Related to Energy Metabolism
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Shin, Y.; Choi, H.; Kim, D.; Park, J.; Kim, N. Effect of heat processed ginseng on anti-fatigue. Korean J. Pharmacogn. 2006, 37, 246–252. [Google Scholar]
- Nam, K.-Y. The comparative understanding between red ginseng and white ginsengs, processed ginsengs (Panax ginseng CA Meyer). J. Ginseng Res. 2005, 29, 1–18. [Google Scholar]
- Tark, K.-M.; Son, M.-H.; Chae, H.-J. Optimal Analytical Conditions for Panax Ginseng Ginsenosides using HPLC and Ginsenosides Content Analysis of Red Ginseng Products and their Raw Materials. J. Korea Acad. Coop. Soc. 2009, 10, 418–424. [Google Scholar] [CrossRef]
- Abdel-Wahhab, M.A.; Hassan, N.S.; El-Kady, A.A.; Khadrawy, Y.A.; El-Nekeety, A.; Mohamed, S.R.; Sharaf, H.; Mannaa, F.; Abdel-Wahhab, M.A. Red ginseng extract protects against aflatoxin B1 and fumonisins-induced hepatic pre-cancerous lesions in rats. Food Chem. Toxicol. 2010, 48, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Park, S.E.; Park, C.; Kim, S.H.; Hossain, M.A.; Kim, M.Y.; Chung, H.Y.; Son, W.S.; Kim, G.-Y.; Choi, Y.H.; Kim, N.D. Korean red ginseng extract induces apoptosis and decreases telomerase activity in human leukemia cells. J. Ethnopharmacol. 2009, 121, 304–312. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Hahm, D.H.; Yang, D.C.; Kim, J.H.; Lee, H.J.; Shim, I. Effect of crude saponin of Korean red ginseng on high-fat diet-induced obesity in the rat. J. Pharmacol. Sci. 2005, 97, 124–131. [Google Scholar] [CrossRef]
- Shin, S.S.; Yoon, M. Korean red ginseng (Panax ginseng) inhibits obesity and improves lipid metabolism in high fat diet-fed castrated mice. J. Ethnopharmacol. 2018, 210, 80–87. [Google Scholar] [CrossRef]
- Kim, J.H.; Kang, S.A.; Han, S.-M.; Shim, I. Comparison of the antiobesity effects of the protopanaxadiol- and protopanaxatriol-type saponins of red ginseng. Phytother. Res. 2009, 23, 78–85. [Google Scholar] [CrossRef]
- Hwang, H.; Kim, J.; Park, J.; Yun, H.; Cheon, W.-K.; Kim, B.; Lee, C.-H.; Suh, H.; Lim, K. Red Ginseng Treatment for Two Weeks Promotes Fat Metabolism during Exercise in Mice. Nutrients 2014, 6, 1874–1885. [Google Scholar] [CrossRef]
- Jung, S.; Lee, M.-S.; Shin, Y.; Kim, C.-T.; Kim, I.-H.; Kim, Y.S.; Kim, Y. Anti-obesity and anti-inflammatory effects of high hydrostatic pressure extracts of ginseng in high-fat diet induced obese rats. J. Funct. Foods 2014, 10, 169–177. [Google Scholar] [CrossRef]
- Karu, N.; Reifen, R.; Kerem, Z. Weight Gain Reduction in Mice FedPanaxginsengSaponin, a Pancreatic Lipase Inhibitor. J. Agric. Food Chem. 2007, 55, 2824–2828. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.J.; Lee, Y.-H.; Park, S.K.; Kang, E.S.; Kim, H.-J.; Lee, Y.C.; Choi, C.S.; Park, S.E.; Ahn, C.W.; Cha, B.-S.; et al. Korean red ginseng (Panax ginseng) improves insulin sensitivity and attenuates the development of diabetes in Otsuka Long-Evans Tokushima fatty rats. Metabolism 2009, 58, 1170–1177. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Lee, H.J.; Lee, Y.-H.; Lee, B.-W.; Cha, B.-S.; Kang, E.S.; Ahn, C.W.; Park, J.S.; Kim, H.J.; Lee, E.Y.; et al. Korean Red Ginseng (Panax ginseng) Improves Insulin Sensitivity in High Fat Fed Sprague-Dawley Rats. Phytother. Res. 2011, 26, 142–147. [Google Scholar] [CrossRef]
- Oh, J.S.; Lee, S.R.; Hwang, K.T.; Ji, G.E. The Anti-Obesity Effects of the Dietary Combination of Fermented Red Ginseng with Levan in High Fat Diet Mouse Model. Phytother. Res. 2013, 28, 617–622. [Google Scholar] [CrossRef]
- Erickson, J.C.; Hollopeter, G.; Palmiter, R.D. Attenuation of the Obesity Syndrome of ob/ob Mice by the Loss of Neuropeptide Y. Science 1996, 274, 1704–1707. [Google Scholar] [CrossRef]
- Park, S.-J.; Lee, D.; Kim, D.; Lee, M.; In, G.; Han, S.-T.; Kim, S.W.; Lee, M.-H.; Kim, O.-K.; Lee, J. The non-saponin fraction of Korean Red Ginseng (KGC05P0) decreases glucose uptake and transport in vitro and modulates glucose production via down-regulation of the PI3K/AKT pathway in vivo. J. Ginseng Res. 2020, 44, 362–372. [Google Scholar] [CrossRef]
- Xiao, H.; Tan, C.; Yang, G.; Dou, D. The effect of red ginseng and ginseng leaves on the substance and energy metabolism in hypothyroidism rats. J. Ginseng Res. 2017, 41, 556–565. [Google Scholar] [CrossRef]
- Cho, H.-M.; Kang, Y.-H.; Yoo, H.; Yoon, S.-Y.; Kang, S.-W.; Chang, E.-J.; Song, Y. Panax red ginseng extract regulates energy expenditures by modulating PKA dependent lipid mobilization in adipose tissue. Biochem. Biophys. Res. Commun. 2014, 447, 644–648. [Google Scholar] [CrossRef]
- Bang, H.; Kwak, J.H.; Ahn, H.Y.; Shin, D.Y.; Lee, J.H. Korean Red Ginseng Improves Glucose Control in Subjects with Impaired Fasting Glucose, Impaired Glucose Tolerance, or Newly Diagnosed Type 2 Diabetes Mellitus. J. Med. Food 2014, 17, 128–134. [Google Scholar] [CrossRef]
- Na Hong, B.; Ji, M.G.; Kang, T.H. The Efficacy of Red Ginseng in Type 1 and Type 2 Diabetes in Animals. Evidence-Based Complement. Altern. Med. 2013, 2013, 1–7. [Google Scholar] [CrossRef]
- Mu, Q.; Fang, X.; Li, X.; Zhao, D.; Mo, F.; Jiang, G.; Yu, N.; Zhang, Y.; Guo, Y.; Fu, M.; et al. Ginsenoside Rb1 promotes browning through regulation of PPARγ in 3T3-L1 adipocytes. Biochem. Biophys. Res. Commun. 2015, 466, 530–535. [Google Scholar] [CrossRef] [PubMed]
- Li, J.-B.; Zhang, R.; Han, X.; Piao, C.-L. Ginsenoside Rg1 inhibits dietary-induced obesity and improves obesity-related glucose metabolic disorders. Braz. J. Med Boil. Res. 2018, 51, 51. [Google Scholar] [CrossRef] [PubMed]
- Karunasagara, S.; Hong, G.-L.; Park, S.-R.; Lee, N.-H.; Jung, D.-Y.; Kim, T.W.; Jung, J.-Y. Korean red ginseng attenuates hyperglycemia-induced renal inflammation and fibrosis via accelerated autophagy and protects against diabetic kidney disease. J. Ethnopharmacol. 2020, 254, 112693. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.; Kim, S.; Han, S.; Suh, H. PND90’The impact of ministry of food and drug safety (mfds) safety notice on the use of metoclopramide in korea: A population-based study with an interrupted time series analysis. Value Health 2018, 21, S344. [Google Scholar] [CrossRef]
- Coles, L.; Litt, J.; Hatta, H.; Bonen, A. Exercise rapidly increases expression of the monocarboxylate transporters MCT1 and MCT4 in rat muscle. J. Physiol. 2004, 561, 253–261. [Google Scholar] [CrossRef]
- Sebastián, D.; Guitart, M.; Martinez, C.G.; Mauvezin, C.; Orellana-Gavaldà, J.M.; Serra, D.; Gómez-Foix, A.M.; Hegardt, F.G.; Asins, G. Novel role of FATP1 in mitochondrial fatty acid oxidation in skeletal muscle cells. J. Lipid Res. 2009, 50, 1789–1799. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession Number | Sequences | Annealing Temperature (°C) | Product Length (bp) |
|---|---|---|---|---|
| GAPDH | BC145810.1 | F-5′-aactttggcattgtg gaagg–3′ R-5′-acacattgggggtaggaaca–3′ | 52 | 215 |
| GLUT4 | AB008453.1 | F-5′-ttctggctctcacagtactc–3′ R–5′-cattgatgcctgagagctgt–3′ | 52 | 300 |
| MCT1 | NM_009196.4 | F-5′-gctggaggtcctatcagcag–3′ R-5′-agttgaaagcaagcccaaga–3′ | 52 | 172 |
| MCT4 | NM_001038654 | F-5′-acggctggtttcataacagg-3′ R-5′-ccaatggcactggagaactt-3′ | 52 | 233 |
| FAT/CD 36 | NM_007643.4 | F-5′-ggccaagctattgcgacat-3′ R-5′-cagatccgaacacagcgtaga-3′ | 53 | 129 |
| CPT1b | NM_009948.2 | F-5′-atcatgtatcgccgcaaact-3′ R-5′-ccatctggtaggagcacatgg-3′ | 53 | 85 |
| CON | RG | |
|---|---|---|
| Initial body weight (g) | 35.79 ± 0.31 | 35.87 ± 0.56 |
| Final body weight (g) | 36.10 ± 0.38 | 36.80 ± 0.58 |
| Body weight gain (g) | 0.31 ± 0.29 | 0.93 ± 0.30 |
| Total amount of food intake (g/2 weeks) | 96.23 ± 2.67 | 85.95 ± 0.96 ** |
| Total amount of food intake (kcal/2 weeks) | 308.92 ± 8.96 | 275.91 ± 3.23 ** |
| Daily food intake (g) | 6.01 ± 0.16 | 5.37 ± 0.06 ** |
| Daily food intake (kcal) | 19.31 ± 0.56 | 17.24 ± 0.20 ** |
| Epididymal (g) | 0.71 ± 0.05 | 0.86 ± 0.09 |
| Perirenal (g) | 0.32 ± 0.03 | 0.38 ± 0.05 |
| Mesenteric (g) | 0.68 ± 0.02 | 0.66 ± 0.03 |
| Total adipose tissue (g) | 1.70 ± 0.01 | 1.90 ± 0.03 |
| CON | RG | |
|---|---|---|
| RMR (kcal/day) | 8.42 ± 0.57 | 7.77 ± 0.55 |
| Active period (kcal/12 h) | 4.26 ± 0.24 | 4.00 ± 0.36 |
| Inactive period (kcal/12 h) | 4.52 ± 0.25 | 4.44 ± 0.37 |
| CON | RG | |
|---|---|---|
| Glucose (mg/dL) | 159.61 ± 11.96 | 164.10 ± 10.39 |
| Insulin (ng/mg) | 0.91 ± 0.09 | 1.03 ± 0.16 |
| FFA (mEq/L) | 0.46 ± 0.06 | 0.38 ± 0.03 |
| CON | RG | |
|---|---|---|
| Liver | 343.1 ± 12.2 | 330.2 ± 20.1 |
| Gastrocnemius white muscle | 29.7 ± 2.3 | 31.4 ± 2.2 |
| Gastrocnemius red muscle | 17.8 ± 2.3 | 16.7 ± 0.5 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hwang, H.; Kim, J.; Lim, K. The Effect of a 2-Week Red Ginseng Supplementation on Food Efficiency and Energy Metabolism in Mice. Nutrients 2020, 12, 1726. https://doi.org/10.3390/nu12061726
Hwang H, Kim J, Lim K. The Effect of a 2-Week Red Ginseng Supplementation on Food Efficiency and Energy Metabolism in Mice. Nutrients. 2020; 12(6):1726. https://doi.org/10.3390/nu12061726
Chicago/Turabian StyleHwang, Hyejung, Jisu Kim, and Kiwon Lim. 2020. "The Effect of a 2-Week Red Ginseng Supplementation on Food Efficiency and Energy Metabolism in Mice" Nutrients 12, no. 6: 1726. https://doi.org/10.3390/nu12061726
APA StyleHwang, H., Kim, J., & Lim, K. (2020). The Effect of a 2-Week Red Ginseng Supplementation on Food Efficiency and Energy Metabolism in Mice. Nutrients, 12(6), 1726. https://doi.org/10.3390/nu12061726