Inhibition of Neuromuscular Contractions of Human and Rat Colon by Bergamot Essential Oil and Linalool: Evidence to Support a Therapeutic Action

 ,
,
Abstract
1. Introduction
2. Methods
2.1. Human Colon
2.2. Rat Colon
2.3. Tissue Bath Technique
2.4. Materials
2.5. Data Analysis
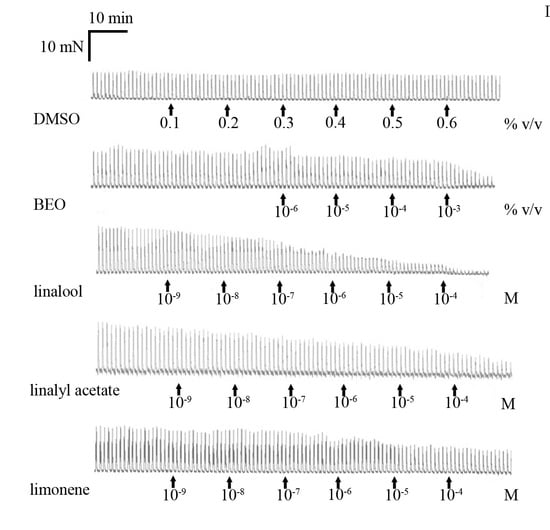
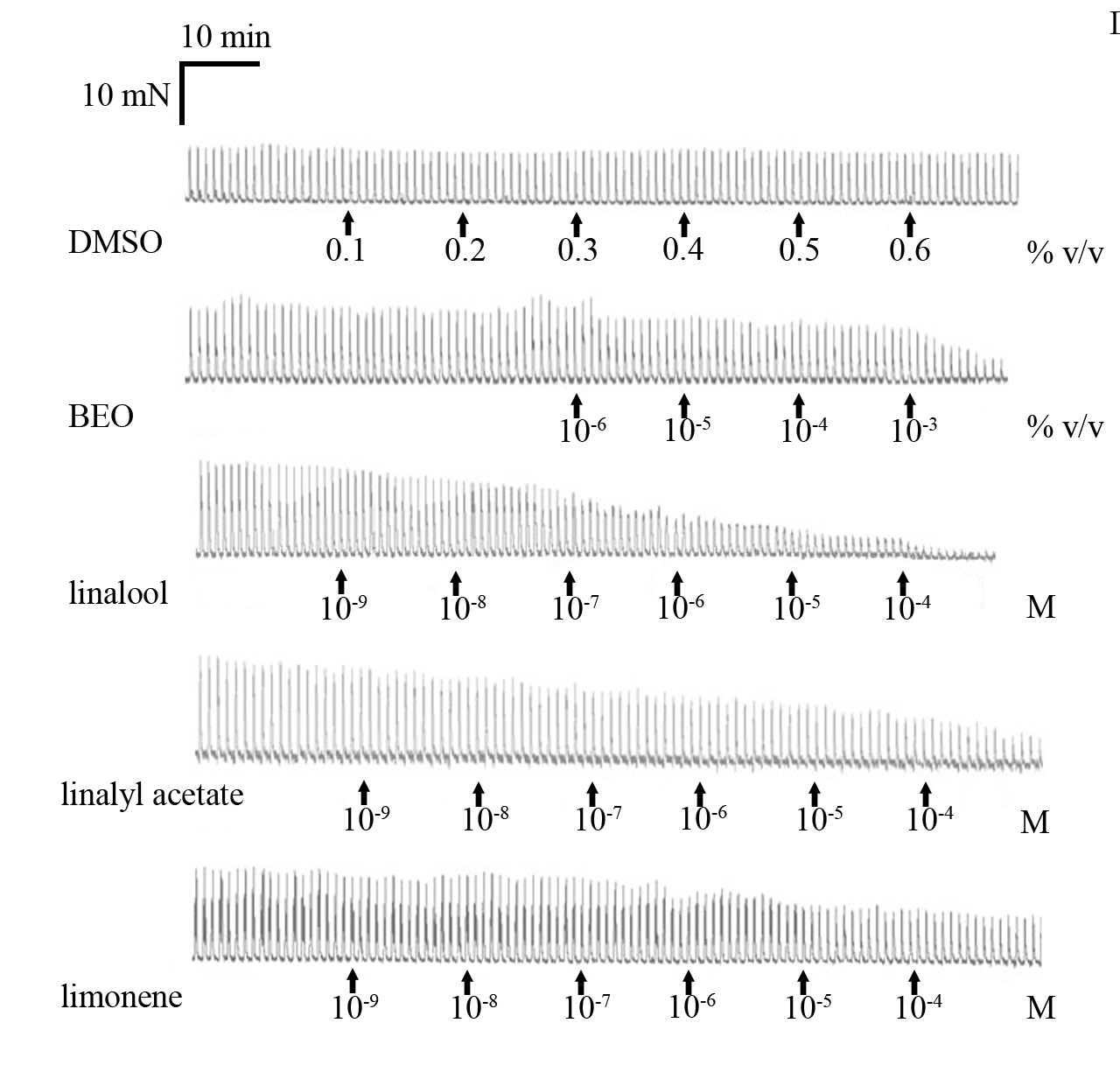
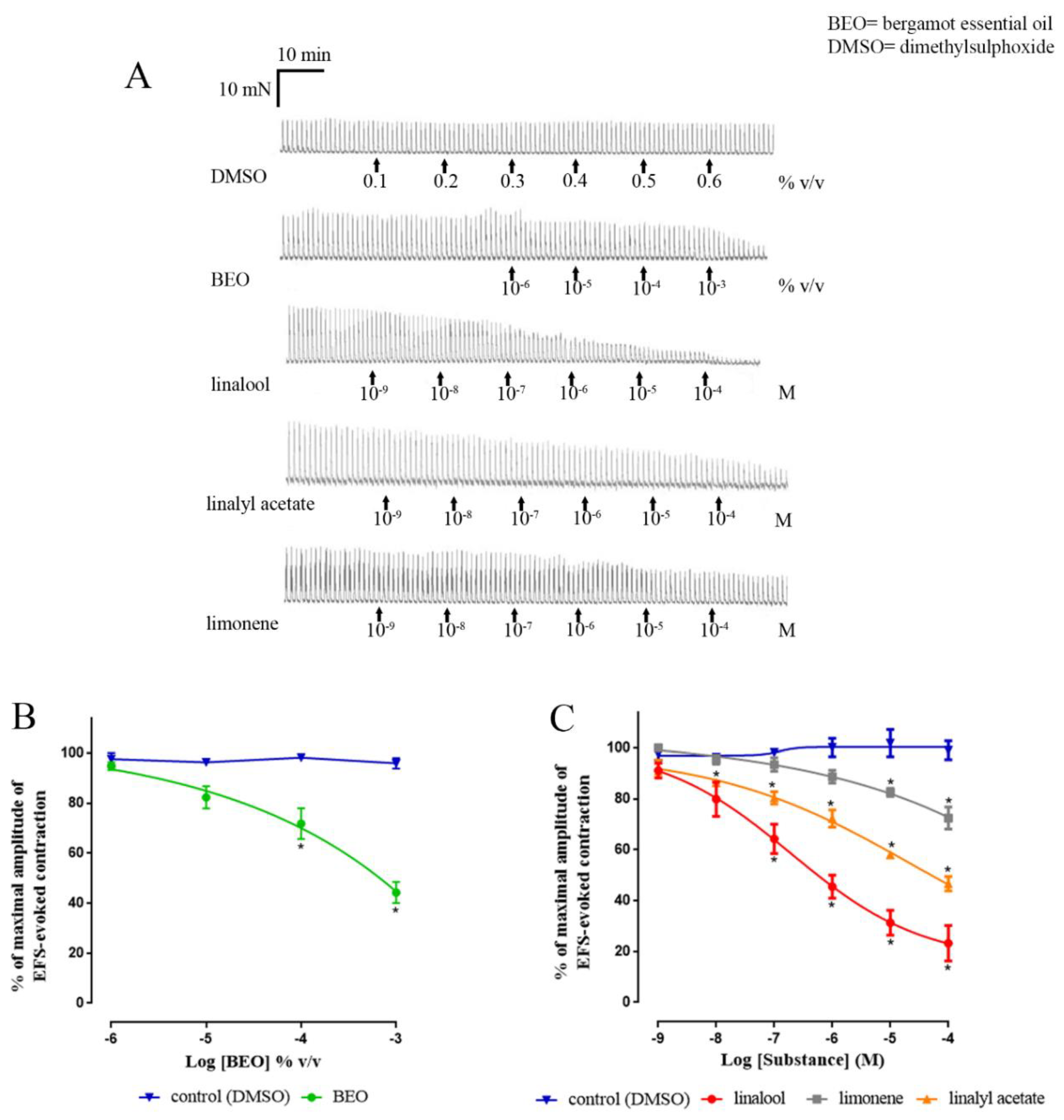
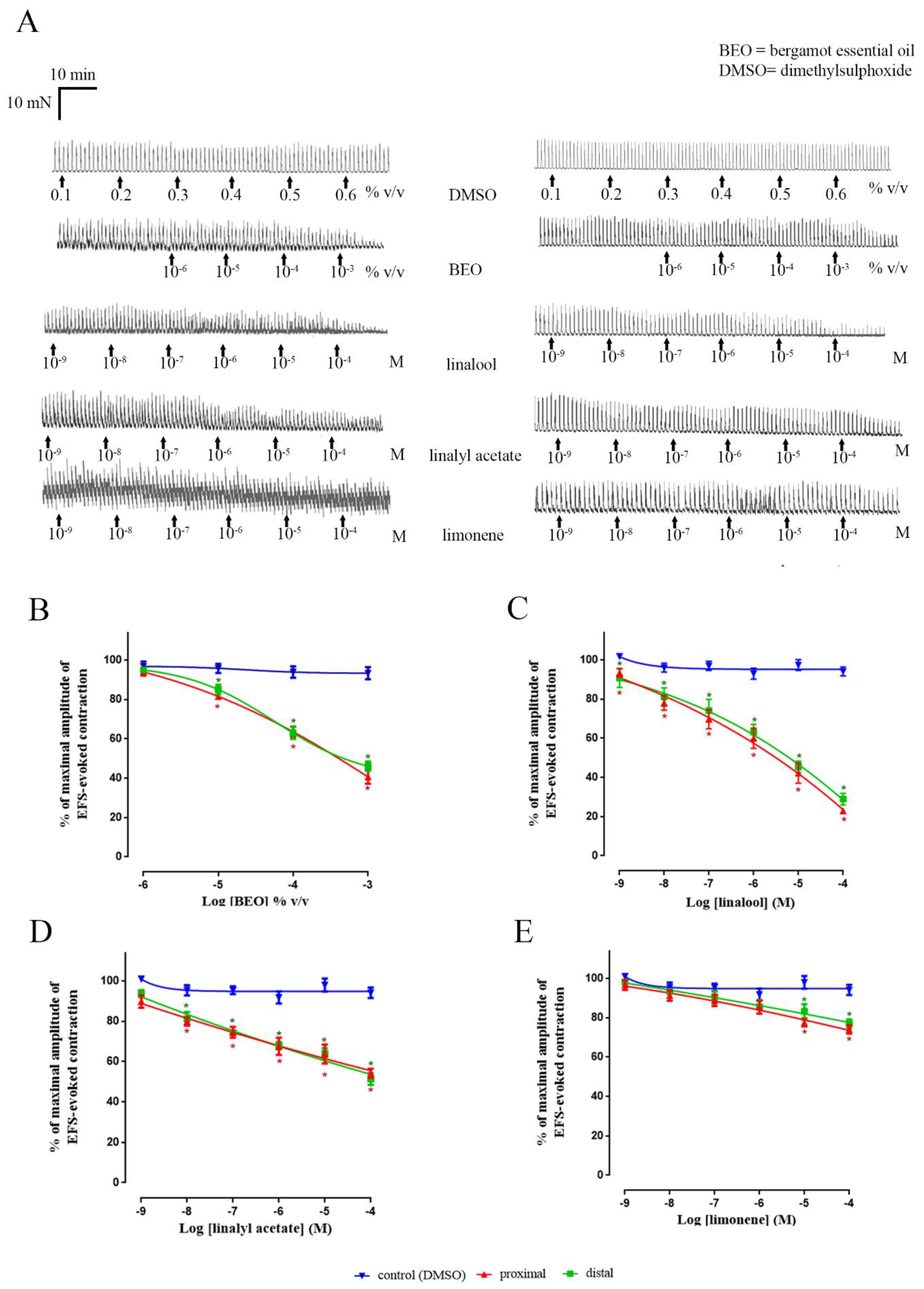
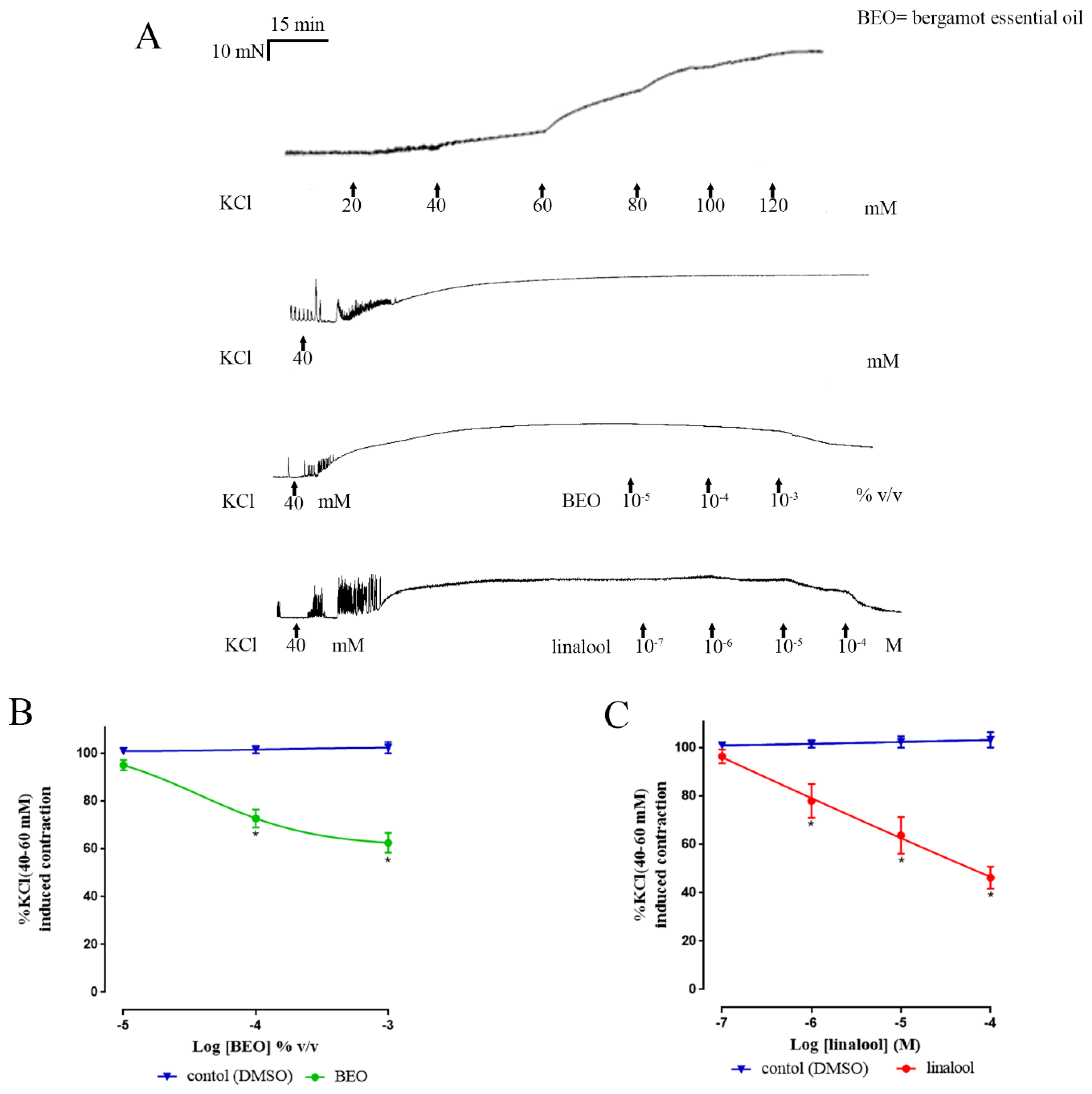
3. Results
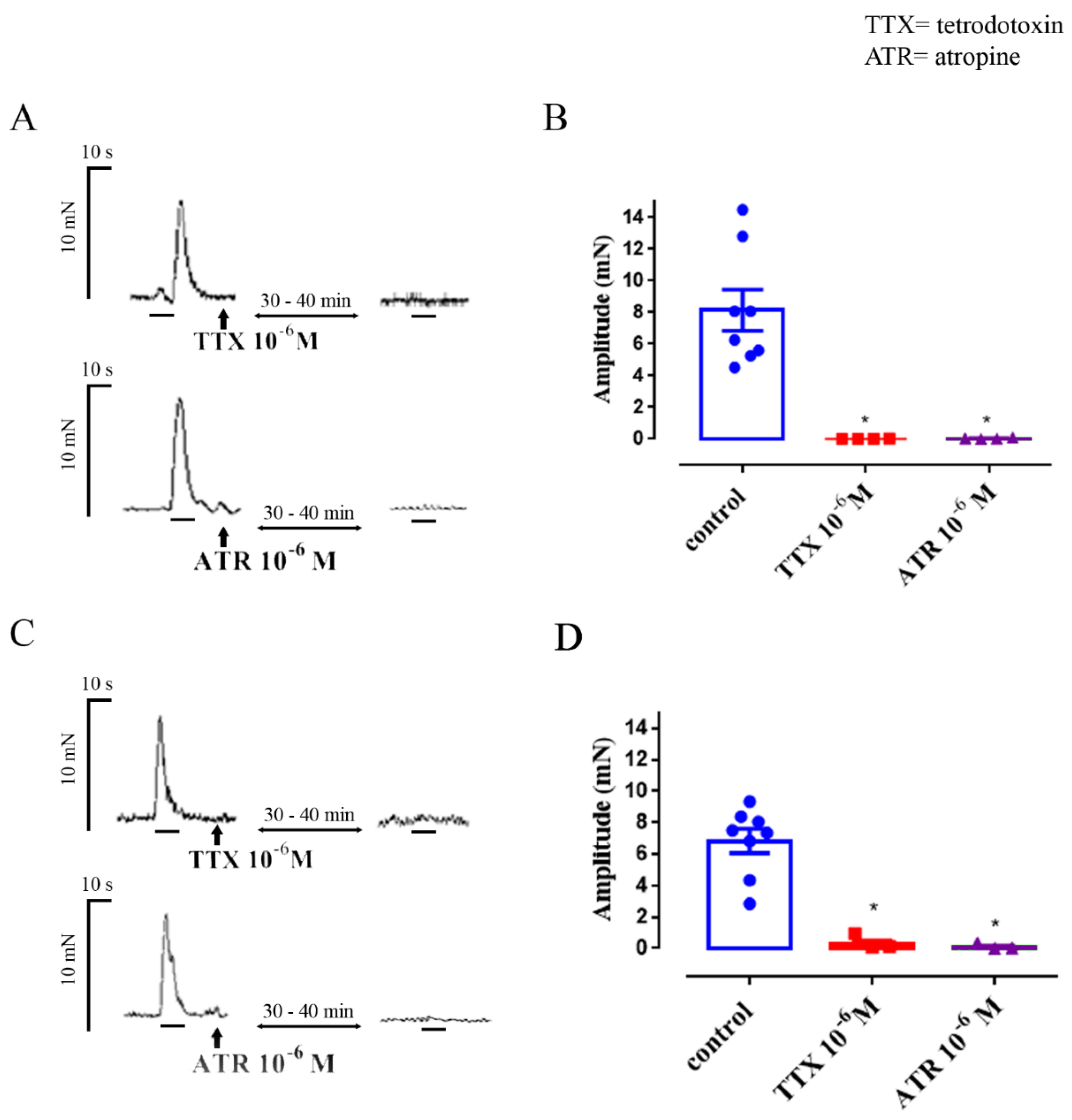
3.1. Neuronally-Mediated Contractions
3.2. Muscle Contractions Evoked by KCl
4. Discussion
5. Conclusions
Ackonwledgement
Author Contributions
Funding
Conflicts of Interest
References
- Calapai, G.; Delbò, M. Assessment Report on Citrus Bergamia Risso et Poiteau, Aetheroleum; EMA/HMPC/56155/2011; Committee on Herbal Medicine Products (HMPC), European Medicine Agency: London, UK, 2012. [Google Scholar]
- Costa, R.; Dugo, P.; Navarra, M.; Raymo, V.; Dugo, G.; Mondelloa, L. Study on the chemical composition variability of some processed bergamot (Citrus bergamia) essential oils. Flavour Fragr. J. 2009, 25, 4–12. [Google Scholar] [CrossRef]
- Donato, P.; Bonaccorsi, I.; Russo, M.; Dugo, P. Determination of new bioflavonoids in bergamot (Citrus bergamia) peel oil by liquid chromatography coupled to tandem ion-trap–time-of-flight mass spectrometry. Flavour Fragr. J. 2014, 29, 131–136. [Google Scholar] [CrossRef]
- Melliou, E.; Michaelakis, A.; Koliopoulos, G.; Skaltsounis, A.L.; Magiatis, P. High quality bergamot oil from Greece: Chemical analysis using chiral gas chromatography and larvicidal activity against the West Nile virus vector. Molecules 2009, 14, 839–849. [Google Scholar] [PubMed]
- Laird, K.; Armitage, D.; Phillips, C. Reduction of surface contamination and biofilms of Enterococcus sp. and Staphylococcus aureus using a citrus-based vapour. J. Hosp. Infect. 2012, 80, 61–66. [Google Scholar] [CrossRef] [PubMed]
- Sanguinetti, M.; Posteraro, B.; Romano, L.; Battaglia, F.; Lopizzo, T.; de Caroli, E.; Fadda, G. In vitro activity of Citrus bergamia (bergamot) oil against clinical isolates of dermatophytes. J. Antimicrob. Chemother. 2007, 59, 305–308. [Google Scholar] [CrossRef] [PubMed]
- Cosentino, M.; Luini, A.; Bombelli, R.; Corasaniti, M.T.; Bagetta, G.; Marino, F. The essential oil of bergamot stimulates reactive oxygen species production in human polymorphonuclear leukocytes. Phytother. Res. 2014, 28, 232–239. [Google Scholar] [CrossRef] [PubMed]
- Bagetta, G.; Morrone, L.A.; Rombolà, L.; Amantea, D.; Russo, R.; Berliocchi, L.; Sakurad, S.; Sakurada, T.; Rotiroti, D.; Corasaniti, M.T. Neuropharmacology of the essential oil of bergamot. Fitoterapia 2010, 81, 453–461. [Google Scholar] [CrossRef] [PubMed]
- Morrone, L.A.; Rombola, L.; Pelle, C.; Corasaniti, M.T.; Zappettini, S.; Paudice, P.; Bonanno, G.; Bagetta, G. The essential oil of bergamot enhances the levels of amino acid neurotransmitters in the hippocampus of rat: Implication of monoterpene hydrocarbons. Pharmacol. Res. 2007, 55, 255–262. [Google Scholar] [CrossRef]
- Amantea, D.; Fratto, V.; Maida, S.; Rotiroti, D.; Ragusa, S.; Nappi, G.; Bagetta, G.; Corasaniti, M.T. Prevention of glutamate accumulation and upregulation of phospho-akt may account for neuroprotection afforded by Bergamot Essential Oil against brain injury induced by focal cerebral ischemia in rat. Int. Rev. Neurobiol. 2009, 85, 389–405. [Google Scholar]
- Rombolà, L.; Tridico, L.; Scuteri, D.; Sakurada, T.; Sakurada, S.; Mizoguchi, H.; Avato, P.; Corasaniti, M.T.; Bagetta, G.; Morrone, L.A. Bergamot essential oil attenuates anxiety-like behaviour in rats. Molecules 2017, 22, 614. [Google Scholar] [CrossRef]
- Kang, P.; Suh, S.H.; Min, S.S.; Seol, G.H. The essential oil of Citrus bergamia Risso induces vasorelaxation of the mouse aorta by activating K+ channels and inhibiting Ca2+ influx. J. Pharm. Pharmacol. 2013, 65, 745–749. [Google Scholar] [CrossRef] [PubMed]
- Blanco, M.A.; Colareda, G.A.; van Baren, C.; Bandoni, A.L.; Ringuelet, J.; Consolini, A.E. Antispasmodic effects and composition of the essential oils from two South American chemotypes of Lippia alba. J. Ethnopharmacol. 2013, 149, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Sanger, G.J.; Broad, J.; Kung, V.; Knowles, C.H. Translational Neuropharmacology: The use of human isolated gastrointestinal tissues. Br. J. Pharmacol. 2013, 168, 28–43. [Google Scholar] [CrossRef] [PubMed]
- Straface, M.; Makwana, R.; Rombola, L.; Chinaleong, J.; Morrone, L.A.; Sanger, G.J. Bergamot essential oil and its constituent linalool, inhibit cholinergically-mediated contractions of human and rat isolated colon. Neurogastroenterol. Motil. 2019, 31 (Suppl. 4), 154. [Google Scholar]
- Northup, D.W.; Stickney, J.C.; Van Liere, E.J. Effect of carbon dioxide on intestinal motility. Am. J. Physiol. 1949, 158, 119–121. [Google Scholar] [CrossRef] [PubMed]
- Broad, J.; Mukherjee, S.; Samadi, M.; Martin, J.E.; Dukes, G.E.; Sanger, G.J. Regional- and agonist-dependent facilitation of human neurogastrointestinal functions by motilin receptor agonists. Br. J. Pharmacol. 2012, 167, 763–774. [Google Scholar] [CrossRef]
- Bassil, A.K.; Borman, R.A.; Jarvie, E.M.; McArthur-Wilson, R.J.; Thangiah, R.; Sung, E.Z.; Lee, K.; Sanger, G.J. Activation of prostaglandin EP receptors by lubiprostone in rat and human stomach and colon. Br. J. Pharmacol. 2008, 154, 126–135. [Google Scholar] [CrossRef]
- Karaki, H.; Ozaki, H.; Hori, M.; Mitsui-Saito, M.; Amano, K.; Harada, K.; Miyamoto, S.; Nakazawa, H.; Won, K.J.; Sato, K. Calcium movements, distribution, and functions in smooth muscle. Pharmacol. Rev. 1997, 49, 157–230. [Google Scholar]
- Rodriguez, R.; Ventura-Martinez, R.; Santiago-Mejia, J.; Avila-Costa, M.R.; Fortoul, T.I. Altered responsiveness of the guinea-pig isolated ileum to smooth muscle stimulants and to electrical stimulation after in situ ischemia. Br. J. Pharmacol. 2006, 147, 371–378. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential oils’ chemical characterization and investigation of some biological activities: A critical review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef]
- Lis-Balchin, M.; Hart, S. Studies on the mode of action of the essential oil of lavender (Lavandula angustifolia P. Miller). Phytother. Res. 1999, 13, 540–542. [Google Scholar] [CrossRef]
- De Cássia da Silveira e Sá, R.; Lima, T.C.; Da Nóbrega, F.R.; Medeiros de Brito, A.E.; De Sousa, D.P. Analgesic-like activity of essential oil constituents: An update. Int. J. Mol. Sci. 2017, 18, 2392. [Google Scholar] [CrossRef] [PubMed]
- Vatanparast, J.; Bazleh, S.; Janahmadi, M. The effects of linalool on the excitability of central neurons of snail Caucasotachea atrolabiata. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 192, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Narusuye, K.; Kawai, F.; Matsuzaki, K.; Miyachi, E. Linalool suppresses voltage-gated currents in sensory neurons and cerebellar Purkinje cells. J. Neural. Transm. 2005, 112, 193–203. [Google Scholar] [CrossRef] [PubMed]
- Okamura, H.; Fujitani, B.; Furukawa, K.; Une, T.; Nukuda, T.; Komiya, M.; Kurokawa, M. Pharmacologic properties of a novel Ca2+ entry blocker, AJ-2615, in vitro. J. Cardiovasc. Pharmacol. 1993, 22, 804–809. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, G.E.; Barbosa, R.; Thompson, A.J. Non-competitive inhibition of 5-HT3 receptors by citral, linalool, and eucalyptol revealed by nonlinear mixed-effects modelling. J. Pharmacol. Exp. Ther. 2016, 356, 549–562. [Google Scholar] [CrossRef]
- Behrendt, H.J.; Germann, T.; Gillen, C.; Hatt, H.; Jostock, R. Characterization of the mouse cold-menthol receptor TRPM8 and vanilloid receptor type-1 VR1 using a fluorometric imaging plate reader (FLIPR) assay. Br. J. Pharmacol. 2004, 141, 737–745. [Google Scholar] [CrossRef]
- Paschke, M.; Tkachenko, A.; Ackermann, K.; Hutzler, C.; Henkler, F.; Luch, A. Activation of the cold-receptor TRPM8 by low levels of menthol in tobacco products. Toxicol. Lett. 2017, 271, 50–57. [Google Scholar] [CrossRef]
- Fothergill, L.J.; Callaghan, B.; Rivera, L.R.; Lieu, T.; Poole, D.P.; Cho, H.J.; Bravo, D.M.; Furness, J.B. Effects of food components that activate TRPA1 receptors on mucosal ion transport in the mouse intestine. Nutrients 2016, 8, 623. [Google Scholar] [CrossRef]
- Annaházi, A.; Róka, R.; Rosztóczy, A.; Wittmann, T. Role of antispasmodics in the treatment of irritable bowel syndrome. World J. Gastroenterol. 2014, 20, 6031–6043. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Human | |||||||||
| Ascending | Descending | Ascending + Descending | |||||||
| Apparent pIC50 | Apparent Imax (%) | n | Apparent pIC50 | Apparent Imax (%) | n | Apparent pIC50 | Apparent Imax (%) | n | |
| Control (DMSO) | - | 4.6 ± 1.4 | 2 | - | 3.2 ± 3.2 | 2 | - | 3.9 ± 2.7 | 4 |
| BEO | 4.7 ± 0.3 | 59.1 ± 6.6 | 2 | 3.1 ± 0.6 | 53.1 ± 11.3 | 3 | 3.8 ± 0.3 | 55.8 ± 4.2 * | 5 |
| linalool | 6.3 ± 0.3 | 88.8 ± 5.4 | 2 | 6.8 ± 0.2 | 64.8 ± 3.2 | 2 | 6.7 ± 0.2 | 76.8 ± 6.9 * | 4 |
| linalyl acetate | 4.7 ± 0.6 | 58.4 ± 0.4 | 2 | 6.3 ± 0.3 | 48.4 ± 0.2 | 2 | 4.4 ± 0.4 | 53.3 ± 2.9 * | 4 |
| limonene | 5.2 ± 0.3 | 30.6 ± 5.3 | 2 | 5.9 | 21.4 | 1 | 5.5 ± 0.2 | 27.5 ± 4.3 * | 3 |
| Rat | |||||||||
| Proximal | Distal | Proximal + Distal | |||||||
| Apparent pIC50 | Apparent Imax (%) | n | Apparent pIC50 | Apparent Imax (%) | n | Apparent pIC50 | Apparent Imax (%) | n | |
| Control (DMSO) | - | 6.8 ± 4.7 | 3 | - | 4.9 ± 3.5 | 2 | - | 5.9 ± 2.7 | 5 (3 rats) |
| BEO | 4.2 ± 0.2 | 60.8 ± 3.49 * | 4 | 4.1 ± 0.1 | 53.7 ± 2.4 * | 4 | 4 ± 0.3 | 56.3 ± 2.2 * | 8 (4 rats) |
| linalool | 5.8 ± 0.2 | 77.8 ± 1.3 * | 4 | 5.8 ± 0.2 | 69.4 ± 3 * | 4 | 5.8 ± 0.1 | 75.3 ± 1.9 * | 8 (4 rats) |
| linalyl acetate | 6.8 ± 0.3 | 44.5 ± 2.2 * | 4 | 7 ± 0.3 | 49.7 ± 3 * | 4 | 7 ± 0.2 | 49.5 ± 1.7 * | 8 (4 rats) |
| limonene | 5.9 ± 0.3 | 25.2 ± 2.1 * | 4 | 6.4 ± 0.4 | 23.1 ± 2.2 * | 4 | 6.1 ± 0.3 | 24.7 ± 1.5 * | 8 (4 rats) |
| Human | |||||||||
| Ascending | Descending | Ascending + Descending | |||||||
| Apparent pIC50 | Apparent Imax (%) | n | Apparent pIC50 | Apparent Imax (%) | n | Apparent pIC50 | Apparent Imax (%) | n | |
| Control (40–60 mM KCl) | - | 6.41 ± 6.3 | 2 | - | 0.2 ± 0.1 | 2 | - | 3.2 ± 3.3 | 4 |
| BEO | 4.4 ± 0.8 | 38.8 ± 19.6 | 2 | 4.5 ± 0.2 | 36.1 ± 2.9 | 2 | 4.4 ± 0.3 | 37.5 ± 4.2 * | 4 |
| linalool | 5.3 ± 0.2 | 58.9 ± 4.3 | 2 | 5.9 ± 0.5 | 52.3 ± 4.2 | 2 | 5.6 ± 0.4 | 53.8 ± 4.6 * | 4 |
| Rat | |||||||||
| Proximal | Distal | Proximal + Distal | |||||||
| Apparent pIC50 | Apparent Imax (%) | n | Apparent pIC50 | Apparent Imax (%) | n | Apparent pIC50 | Apparent Imax (%) | n | |
| Control (40–60 mM KCl) | - | 1.4 ± 0.73 | 4 | - | 0.6 ± 5.6 | 4 | - | 0.9 ± 5.1 | 8 (4 rats) |
| BEO | 4.4 ± 0.4 | 32.8 ± 3.2 * | 4 | 3.7 ± 0.4 | 19.9 ± 5.5 * | 4 | 4.1 ± 0.5 | 26.3 ± 3.8 * | 8 (4 rats) |
| linalool | 5.6 ± 0.2 | 47.6 ± 2.3 *,† | 4 | 4.8 ± 0.2 | 21.7 ± 1 * | 4 | 5.4 ± 0.3 | 36.1 ± 4.8 * | 8 (4 rats) |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Straface, M.; Makwana, R.; Palmer, A.; Rombolà, L.; Aleong, J.C.; Morrone, L.A.; Sanger, G.J. Inhibition of Neuromuscular Contractions of Human and Rat Colon by Bergamot Essential Oil and Linalool: Evidence to Support a Therapeutic Action. Nutrients 2020, 12, 1381. https://doi.org/10.3390/nu12051381
Straface M, Makwana R, Palmer A, Rombolà L, Aleong JC, Morrone LA, Sanger GJ. Inhibition of Neuromuscular Contractions of Human and Rat Colon by Bergamot Essential Oil and Linalool: Evidence to Support a Therapeutic Action. Nutrients. 2020; 12(5):1381. https://doi.org/10.3390/nu12051381
Chicago/Turabian StyleStraface, Marilisa, Raj Makwana, Alexandra Palmer, Laura Rombolà, Joanne Chin Aleong, Luigi Antonio Morrone, and Gareth J. Sanger. 2020. "Inhibition of Neuromuscular Contractions of Human and Rat Colon by Bergamot Essential Oil and Linalool: Evidence to Support a Therapeutic Action" Nutrients 12, no. 5: 1381. https://doi.org/10.3390/nu12051381
APA StyleStraface, M., Makwana, R., Palmer, A., Rombolà, L., Aleong, J. C., Morrone, L. A., & Sanger, G. J. (2020). Inhibition of Neuromuscular Contractions of Human and Rat Colon by Bergamot Essential Oil and Linalool: Evidence to Support a Therapeutic Action. Nutrients, 12(5), 1381. https://doi.org/10.3390/nu12051381