Supplementation of Sulfur-Containing Amino Acids or Essential Amino Acids Does Not Reverse the Hepatic Lipid-Lowering Effect of a Protein-Rich Insect Meal in Obese Zucker Rats

 , ,
, ,
Abstract
1. Introduction
2. Materials and Methods
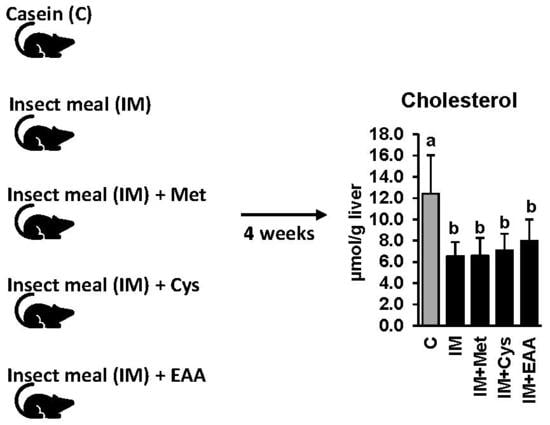
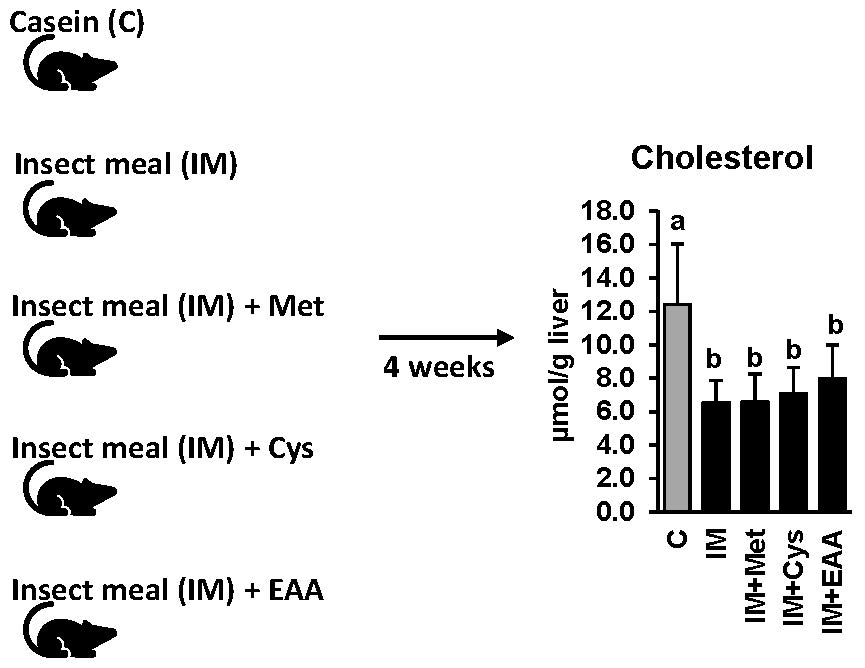
2.1. Animals and Diets
2.2. Analysis of Feed Composition
2.3. Determination of Hepatic Lipid Concentrations
2.4. RNA Extraction and qPCR Analysis
2.5. Determination of Hepatic Enzyme Activities
2.6. Determination of Hepatic Concentrations of Methionine and Its Metabolites
2.7. Determination of Plasma Concentrations of Amino Acids
2.8. Statistical Analysis
3. Results and Discussion
3.1. Characterization of the Experimental Diets
3.2. The Effect of IM, without and with the Adjustment of Dietary Amino Acids, on the Growth Performance of the Rats
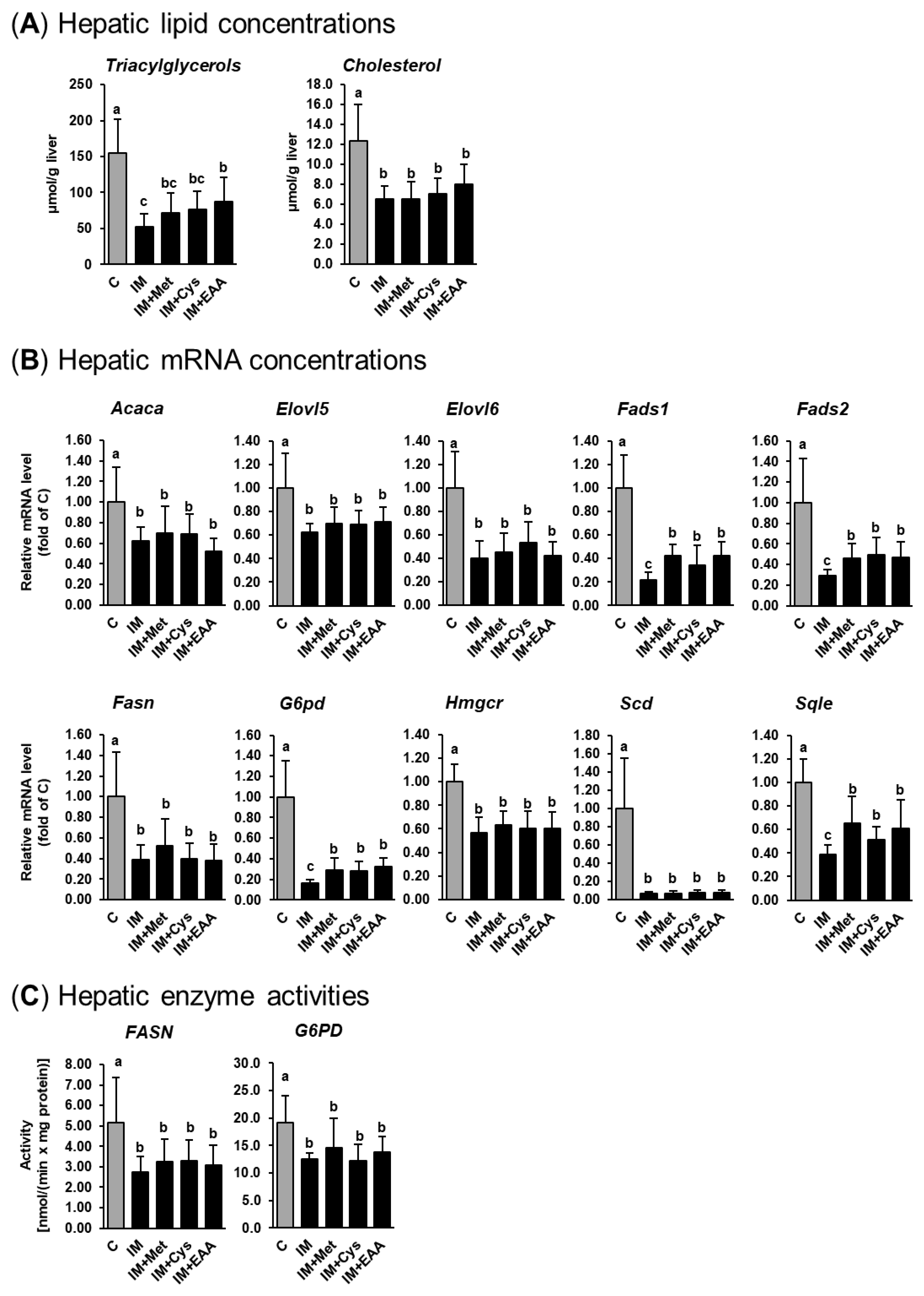
3.3. The Effect of IM, without and with Adjustment of Dietary Amino Acids, on Hepatic Lipid Concentrations and the Expression and Activities of Lipogenic and Cholesterogenic Enzymes
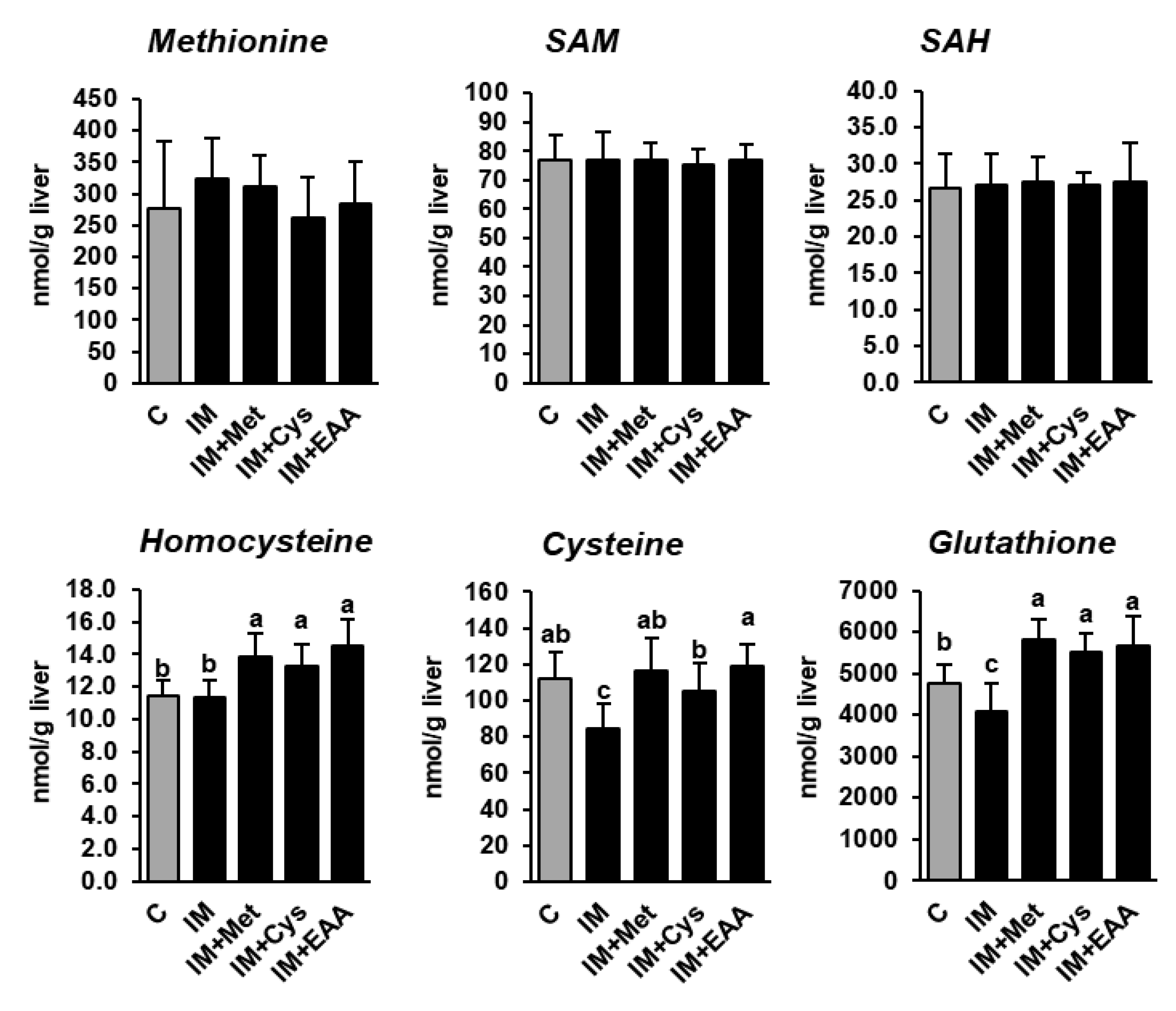
3.4. The Effect of IM, without and with the Adjustment of Dietary Amino Acids, on Hepatic Methionine Metabolism
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- United Nations, Department of Economic and Social Affairs, Population Division. World Population Prospects: The 2017 Revision, Key Findings and Advance Tables. Available online: https://esa.un.org/unpd/wpp/Publications/Files/WPP2017_KeyFindings.pdf (accessed on 10 December 2019).
- Wu, G.; Fanzo, J.; Miller, D.D.; Pingali, P.; Post, M.; Steiner, J.L.; Thalacker-Mercer, A.E. Production and supply of high-quality food protein for human consumption: Sustainability, challenges, and innovations. Ann. N. Y. Acad. Sci. 2014, 1321, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Van Huis, A.; Oonincx, D.G. The environmental sustainability of insects as food and feed. A review. Agron. Sustain. Dev. 2017, 37, 43. [Google Scholar] [CrossRef]
- Oonincx, D.G.; de Boer, I.J. Environmental impact of the production of mealworms as a protein source for humans—A life cycle assessment. PLoS ONE 2012, 7, e51145. [Google Scholar] [CrossRef]
- Jin, X.H.; Heo, P.S.; Hong, J.S.; Kim, N.J.; Kim, Y.Y. Supplementation of Dried Mealworm (Tenebrio molitor larva) on Growth Performance, Nutrient Digestibility and Blood Profiles in Weaning Pigs. Asian-Australas. J. Anim. Sci. 2016, 29, 979–986. [Google Scholar] [CrossRef]
- Biasato, I.; Renna, M.; Gai, F.; Dabbou, S.; Meneguz, M.; Perona, G.; Martinez, S.; Lajusticia, A.C.B.; Bergagna, S.; Sardi, L.; et al. Partially defatted black soldier fly larva meal inclusion in piglet diets: Effects on the growth performance, nutrient digestibility, blood profile, gut morphology and histological features. J. Anim. Sci. Biotechnol. 2019, 10, 12. [Google Scholar] [CrossRef]
- Meyer, S.; Gessner, D.K.; Braune, M.S.; Friedhoff, T.; Most, E.; Höring, M.; Liebisch, G.; Zorn, H.; Eder, K.; Ringseis, R. Comprehensive evaluation of the metabolic effects of insect meal from Tenebrio molitor L. in growing pigs by transcriptomics, metabolomics and lipidomics. J. Anim. Sci. Biotechnol. 2020, in press. [Google Scholar] [CrossRef] [PubMed]
- Ascencio, C.; Torres, N.; Isoard-Acosta, F.; Gómez-Pérez, F.J.; Hernández-Pando, R.; Tovar, A.R. Soy protein affects serum insulin and hepatic SREBP-1 mRNA and reduces fatty liver in rats. J. Nutr. 2004, 134, 522–529. [Google Scholar] [CrossRef]
- Shukla, A.; Brandsch, C.; Bettzieche, A.; Hirche, F.; Stangl, G.I.; Eder, K. Isoflavone-poor soy protein alters the lipid metabolism of rats by SREBP-mediated down-regulation of hepatic genes. J. Nutr. Biochem. 2007, 18, 313–321. [Google Scholar] [CrossRef]
- Bettzieche, A.; Brandsch, C.; Weisse, K.; Hirche, F.; Eder, K.; Stangl, G.I. Lupin protein influences the expression of hepatic genes involved in fatty acid synthesis and triacylglycerol hydrolysis of adult rats. Br. J. Nutr. 2008, 99, 952–962. [Google Scholar] [CrossRef]
- Spielmann, J.; Stangl, G.I.; Eder, K. Dietary pea protein stimulates bile acid excretion and lowers hepatic cholesterol concentration in rats. J. Anim. Physiol. Anim. Nutr. (Berl.) 2008, 92, 683–693. [Google Scholar] [CrossRef]
- Manzoni, C.; Duranti, M.; Eberini, I.; Scharnag, H.; März, W.; Castiglioni, S.; Lovati, M.R. Subcellular localization of soybean 7S globulin in HepG2 cells and LDL receptor up-regulation by its alpha’ constituent subunit. J. Nutr. 2003, 133, 2149–2155. [Google Scholar] [CrossRef] [PubMed]
- Onomi, S.; Okazaki, Y.; Katayama, T. Effect of dietary level of phytic acid on hepatic and serum lipid status in rats fed a high-sucrose diet. Biosci. Biotechnol. Biochem. 2004, 68, 1379–1381. [Google Scholar] [CrossRef] [PubMed]
- Gessner, D.K.; Schwarz, A.; Meyer, S.; Wen, G.; Most, E.; Zorn, H.; Ringseis, R.; Eder, K. Insect Meal as Alternative Protein Source Exerts Pronounced Lipid-Lowering Effects in Hyperlipidemic Obese Zucker Rats. J. Nutr. 2019, 149, 566–577. [Google Scholar] [CrossRef] [PubMed]
- Meyer, S.; Gessner, D.K.; Wen, G.; Most, E.; Liebisch, G.; Zorn, H.; Ringseis, R.; Eder, K. The Antisteatotic and Hypolipidemic Effect of Insect Meal in Obese Zucker Rats is Accompanied by Profound Changes in Hepatic Phospholipid and 1-Carbon Metabolism. Mol. Nutr. Food Res. 2019, 63, e1801305. [Google Scholar] [CrossRef]
- Orentreich, N.; Matias, J.R.; DeFelice, A.; Zimmerman, J.A. Low methionine ingestion by rats extends life span. J. Nutr. 1993, 123, 269–274. [Google Scholar]
- Richie, J.P., Jr.; Leutzinger, Y.; Parthasarathy, S.; Malloy, V.; Orentreich, N.; Zimmerman, J.A. Methionine restriction increases blood glutathione and longevity in F344 rats. FASEB J. 1994, 8, 1302–1307. [Google Scholar] [CrossRef]
- Perrone, C.E.; Mattocks, D.A.; Jarvis-Morar, M.; Plummer, J.D.; Orentreich, N. Methionine restriction effects on mitochondrial biogenesis and aerobic capacity in white adipose tissue, liver, and skeletal muscle of F344 rats. Metabolism 2010, 59, 1000–1011. [Google Scholar] [CrossRef]
- Hirche, F.; Schröder, A.; Knoth, B.; Stangl, G.I.; Eder, K. Effect of dietary methionine on plasma and liver cholesterol concentrations in rats and expression of hepatic genes involved in cholesterol metabolism. Br. J. Nutr. 2006, 95, 879–888. [Google Scholar] [CrossRef]
- van der Veen, J.N.; Lingrell, S.; Vance, D.E. The membrane lipid phosphatidylcholine is an unexpected source of triacylglycerol in the liver. J. Biol. Chem. 2012, 287, 23418–23426. [Google Scholar] [CrossRef]
- Martínez-Uña, M.; Varela-Rey, M.; Cano, A.; Fernández-Ares, L.; Beraza, N.; Aurrekoetxea, I.; Martínez-Arranz, I.; García-Rodríguez, J.L.; Buqué, X.; Mestre, D.; et al. Excess S-adenosylmethionine reroutes phosphatidylethanolamine towards phosphatidylcholine and triglyceride synthesis. Hepatology 2013, 58, 1296–1305. [Google Scholar] [CrossRef]
- Werstuck, G.H.; Lentz, S.R.; Dayal, S.; Hossain, G.S.; Sood, S.K.; Shi, Y.Y.; Zhou, J.; Maeda, N.; Krisans, S.K.; Malinow, M.R.; et al. Homocysteine-induced endoplasmic reticulum stress causes dysregulation of the cholesterol and triglyceride biosynthetic pathways. J. Clin. Investig. 2001, 107, 1263–1273. [Google Scholar] [CrossRef]
- Ghosh, S.; Forney, L.A.; Wanders, D.; Stone, K.P.; Gettys, T.W. An integrative analysis of tissue-specific transcriptomic and metabolomic responses to short-term dietary methionine restriction in mice. PLoS ONE 2017, 12, e0177513. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.; Valdivia-Garcia, M.; Refsum, H.; Smith, A.D.; Mattocks, D.A.L.; Perrone, C.E. Sulfur amino acids in methionine-restricted rats: Hyperhomocysteinemia. Nutrition. Nutrition 2010, 26, 1201–1204. [Google Scholar] [CrossRef] [PubMed]
- Elshorbagy, A.; Valdivia-Garcia, M.; Mattocks, D.A.L.; Plummer, J.D.; Smith, A.D.; Drevon, C.A.; Refsum, H.; Perrone, C.E. Cysteine supplementation reverses methionine restriction effects on rat adiposity: Significance of stearoyl-coenzyme A desaturase. J. Lipid Res. 2011, 52, 104–112. [Google Scholar] [CrossRef] [PubMed]
- VDLUFA (Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten). Die chemische Untersuchung von Futtermitteln. VDLUFA-Methodenbuch. Band III, Ergänzungslieferungen von 1983, 1988, 1992, 1997, 2004, 2006, 2007; VDLUFA-Verlag: Darmstadt, Germany, 2007. [Google Scholar]
- Commission Regulation (EC). No 152/2009 of 27 January 2009 laying down the methods of sampling and analysis for the official control of feed. Off. J. Eur. Union L 2009, 54, 1–130. [Google Scholar]
- Vandesompele, J.; De Preter, K.; Pattyn, F.; Poppe, B.; Van Roy, N.; De Paepe, A.; Speleman, F. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, RESEARCH0034. [Google Scholar] [CrossRef] [PubMed]
- Nepokroeff, C.M.; Lakshmanan, M.R.; Porter, J.W. Fatty-acid synthase from rat liver. Methods Enzymol. 1975, 39, 37–44. [Google Scholar]
- Burren, K.A.; Mills, K.; Copp, A.J.; Greene, N.D. Quantitative analysis of s-adenosylmethionine and s-adenosylhomocysteine in neurulation-stage mouse embryos by liquid chromatography tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2006, 844, 112–118. [Google Scholar] [CrossRef]
- Dewolfe, M.S.; Baskurt, S.; Cochrane, W.A. Automatic amino acid analysis of blood serum and plasma. Clin. Biochem. 1967, 1, 75–81. [Google Scholar] [CrossRef]
- Su, J.; Ma, C.; Liu, C.; Gao, C.; Nie, R.; Wang, H. Hypolipidemic Activity of Peony Seed Oil Rich in α-Linolenic, is Mediated Through Inhibition of Lipogenesis and Upregulation of Fatty Acid β-Oxidation. J. Food Sci. 2016, 81, H1001–H1009. [Google Scholar] [CrossRef]
- National Research Council. Nutrient Requirements of Laboratory Animals, 4th ed.; National Academy Press: Washington, DC, USA, 1995. [Google Scholar]
- Gades, M.D.; Stern, J.S. Chitosan supplementation and fecal fat excretion in men. Obes. Res. 2003, 11, 683–688. [Google Scholar] [CrossRef] [PubMed]
- Walsh, A.M.; Sweeney, T.; Bahar, B.; O’Doherty, J.V. Multi-functional roles of chitosan as a potential protective agent against obesity. PLoS ONE 2013, 8, e53828. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, J.; Li, L.; Xia, W. Dietary chitosan improves hypercholesterolemia in rats fed high-fat diets. Nutr. Res. 2008, 28, 383–390. [Google Scholar] [CrossRef] [PubMed]
- Delbecque, J.P.; Prost, M.; Maume, B.F.; Delachambre, J.; Lafont, R.; Mauchamp, B. Dosage de la β-ecdysone par fragmentographie de masse au cours du dernier stade larvaire de Tenebrio molitor L (Insecte, Coléoptère). C. R. Acad. Sci. Paris 1975, 281, 309–312. [Google Scholar]
- Kizelsztein, P.; Govorko, D.; Komarnytsky, S.; Evans, A.; Wang, Z.; Cefalu, W.T.; Raskin, I. 20-Hydroxyecdysone decreases weight and hyperglycemia in a diet-induced obesity mice model. Am. J. Physiol. Endocrinol. Metab. 2009, 296, E433–E439. [Google Scholar] [CrossRef] [PubMed]
- Foucault, A.S.; Even, P.; Lafont, R.; Dioh, W.; Veillet, S.; Tomé, D.; Huneau, J.F.; Hermier, D.; Quignard-Boulangé, A. Quinoa extract enriched in 20-hydroxyecdysone affects energy homeostasis and intestinal fat absorption in mice fed a high-fat diet. Physiol. Behav. 2014, 128, 226–231. [Google Scholar] [CrossRef]
- Parr, M.K.; Zhao, P.; Haupt, O.; Ngueu, S.T.; Hengevoss, J.; Fritzemeier, K.H.; Piechotta, M.; Schlörer, N.; Muhn, P.; Zheng, W.Y.; et al. Estrogen receptor beta is involved in skeletal muscle hypertrophy induced by the phytoecdysteroid ecdysterone. Mol. Nutr. Food Res. 2014, 58, 1861–1872. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| C | IM | IM+Met | IM+Cys | IM+EAA | |
|---|---|---|---|---|---|
| Components (g/kg) | |||||
| Corn starch | 530 | 530 | 530 | 530 | 530 |
| Casein | 200 | - | - | - | - |
| Insect meal | - | 262 | 262 | 262 | 262 |
| Sucrose | 100 | 100 | 100 | 100 | 100 |
| Soybean oil | 50 | - | - | - | - |
| Safflower oil | - | 19.2 | 19.2 | 19.2 | 19.2 |
| Linseed oil | - | 3.8 | 3.8 | 3.8 | 3.8 |
| Cellulose | 56.5 | 21.9 | 21.9 | 20.9 | 20.8 |
| Mineral mix 1 | 35 | 35 | 35 | 35 | 35 |
| Vitamin mix 2 | 10 | 10 | 10 | 10 | 10 |
| TiO2 | 5 | 5 | 5 | 5 | 5 |
| L-Cysteine (76.9%) | 1.2 | - | - | 3.3 | - |
| DL-Methionine (99%) | - | 0.9 | 3.1 | 0.9 | 3.1 |
| L-Threonine (98.5%) | 0.2 | - | - | - | - |
| L-Valine (96.5%) | 0.6 | - | - | - | - |
| L-Leucine (99%) | - | - | - | - | 3 |
| L-Isoleucine (99%) | - | - | - | - | 0.5 |
| L-Phenylalanine (98.5%) | - | - | - | - | 2.1 |
| L-Histidine (99%) | - | - | - | - | 0.4 |
| L-Lysine (78%) | - | - | - | - | 4.9 |
| L-Arginine (98%) | 3.5 | - | - | - | - |
| L-Tryptophan (98%) | - | - | - | - | 0.1 |
| L-Glutamic acid (98%) | 8.0 | 12.2 | 9.9 | 9.9 | - |
| Analyzed crude nutrient and energy content | |||||
| Dry matter (% FM) | 87.7 | 89.2 | 88.9 | 88.6 | 88.5 |
| Crude protein (% DM) | 23.1 | 23.0 | 22.8 | 22.9 | 23.1 |
| Crude fat (% DM) | 5.3 | 5.5 | 5.5 | 5.5 | 5.4 |
| Crude ash (% DM) | 3.5 | 4.4 | 4.3 | 4.3 | 4.3 |
| Crude fiber (% DM) | 4.6 | 4.4 | 4.5 | 4.4 | 4.5 |
| Gross energy (MJ/kg DM) | 19.5 | 19.3 | 19.3 | 19.3 | 19.4 |
| Concentration | |
|---|---|
| Crude nutrients (% DM) | |
| Crude protein | 77.1 |
| Crude fat | 10.4 |
| Crude fiber | 10.6 |
| Crude ash | 4.4 |
| Chitin | 13.1 |
| Gross energy, MJ/kg DM | 24.1 |
| Amino acids (g/kg FM) | |
| Alanine | 59.5 |
| Arginine | 36.5 |
| Aspartic acid1 | 61.4 |
| Cysteine | 5.7 |
| Glutamic acid2 | 84.5 |
| Glycine | 37.3 |
| Histidine | 18.3 |
| Isoleucine | 33.5 |
| Leucine | 53.2 |
| Lysine | 39.6 |
| Methionine | 8.8 |
| Phenylalanine | 26.9 |
| Proline | 42.3 |
| Serine | 30.0 |
| Threonine | 29.7 |
| Tryptophan | 8.3 |
| Tyrosine | 54.1 |
| Valine | 46.9 |
| Fatty acids3 (g/100 g total fatty acids) | |
| 12:0 | 0.3 |
| 14:0 | 2.4 |
| 16:0 | 17.6 |
| 16:1 n-9 | 0.6 |
| 18:0 | 4.5 |
| 18:1 n-9 | 36.1 |
| 18:2 n-6 | 36.4 |
| 18:3 n-3 | 1.2 |
| 20:0 | 0.1 |
| C | IM | IM+Met | IM+Cys | IM+EAA | |
|---|---|---|---|---|---|
| Amino acids (g/kg FM) | |||||
| Alanine | 5.4 | 12.4 | 12.2 | 12.6 | 12.2 |
| Arginine | 9.9 | 9.3 | 9.1 | 9.5 | 9.3 |
| Aspartic acid1 | 10.8 | 13.0 | 12.5 | 13.4 | 13.1 |
| Cysteine | 1.5 | 1.4 | 1.5 | 3.6 | 1.5 |
| Glutamic acid2 | 46.7 | 31.8 | 29.8 | 30.1 | 20.6 |
| Glycine | 3.3 | 9.2 | 9.1 | 9.4 | 9.0 |
| Histidine | 5.1 | 4.6 | 4.5 | 4.7 | 4.8 |
| Isoleucine | 9.7 | 8.4 | 8.7 | 8.4 | 8.5 |
| Leucine | 16.7 | 13.7 | 13.2 | 13.9 | 15.7 |
| Lysine | 14.4 | 9.6 | 9.3 | 9.7 | 12.8 |
| Methionine | 5.6 | 3.2 | 5.3 | 3.2 | 5.1 |
| Phenylalanine | 9.1 | 6.8 | 6.4 | 6.9 | 8.4 |
| Proline | 20.6 | 11.1 | 10.5 | 10.9 | 10.5 |
| Serine | 9.5 | 7.3 | 7.0 | 7.5 | 7.1 |
| Threonine | 8.0 | 7.2 | 7.2 | 7.5 | 7.3 |
| Tryptophan | 2.3 | 2.1 | 2.1 | 2.0 | 2.1 |
| Tyrosine | 10.1 | 12.7 | 13.2 | 13.4 | 12.6 |
| Valine | 12.5 | 11.4 | 12.4 | 11.5 | 10.9 |
| Fatty acids3 (g/100 g total fatty acids) | |||||
| 12:0 | 0.1 | 0.2 | 0.2 | 0.2 | 0.2 |
| 14:0 | 0.1 | 1.3 | 1.3 | 1.3 | 1.3 |
| 16:0 | 11.2 | 12.9 | 12.9 | 12.8 | 12.6 |
| 16:1 n-9 | 0.1 | 0.3 | 0.3 | 0.3 | 0.2 |
| 18:0 | 5.3 | 4.1 | 4.2 | 4.2 | 4.1 |
| 18:1 n-9 | 26.9 | 26.1 | 26.1 | 26.1 | 25.5 |
| 18:2 n-6 | 49.9 | 50.0 | 49.8 | 49.9 | 50.8 |
| 18:3 n-3 | 4.7 | 3.8 | 3.7 | 3.8 | 4.0 |
| 20:0 | 0.4 | 0.3 | 0.3 | 0.3 | 0.3 |
| C | IM | IM + Met | IM + Cys | IM + EAA | p-Value | |
|---|---|---|---|---|---|---|
| Initial BW, g | 337 ± 46 | 338 ± 44 | 338 ± 46 | 337 ± 41 | 337 ± 43 | 1.000 |
| Final BW, g | 491 ± 45 | 488 ± 35 | 484 ± 56 | 495 ± 36 | 480 ± 40 | 0.945 |
| BW gain, g/d | 5.53 ± 0.94 | 5.35 ± 0.73 | 5.24 ± 0.98 | 5.63 ± 0.79 | 5.11 ± 0.85 | 0.671 |
| Feed intake, g/d | 30.3 ± 2.0 | 31.2 ± 1.6 | 30.7 ± 2.6 | 33.0 ± 1.9 | 31.7 ± 2.6 | 0.356 |
| Feed:gain ratio, g/g | 5.61 ± 0.87 | 5.95 ± 0.94 | 6.03 ± 1.07 | 5.96 ± 0.76 | 6.34 ± 1.02 | 0.552 |
| Liver weight, g/100 g BW | 5.78 ± 0.70 | 5.66 ± 0.49 | 5.72 ± 0.71 | 6.01 ± 0.50 | 6.21 ± 0.75 | 0.301 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Meyer, S.; Schäfer, L.; Röhrig, J.; Maheshwari, G.; Most, E.; Zorn, H.; Ringseis, R.; Eder, K.; Gessner, D.K. Supplementation of Sulfur-Containing Amino Acids or Essential Amino Acids Does Not Reverse the Hepatic Lipid-Lowering Effect of a Protein-Rich Insect Meal in Obese Zucker Rats. Nutrients 2020, 12, 987. https://doi.org/10.3390/nu12040987
Meyer S, Schäfer L, Röhrig J, Maheshwari G, Most E, Zorn H, Ringseis R, Eder K, Gessner DK. Supplementation of Sulfur-Containing Amino Acids or Essential Amino Acids Does Not Reverse the Hepatic Lipid-Lowering Effect of a Protein-Rich Insect Meal in Obese Zucker Rats. Nutrients. 2020; 12(4):987. https://doi.org/10.3390/nu12040987
Chicago/Turabian StyleMeyer, Sandra, Lea Schäfer, Julia Röhrig, Garima Maheshwari, Erika Most, Holger Zorn, Robert Ringseis, Klaus Eder, and Denise K. Gessner. 2020. "Supplementation of Sulfur-Containing Amino Acids or Essential Amino Acids Does Not Reverse the Hepatic Lipid-Lowering Effect of a Protein-Rich Insect Meal in Obese Zucker Rats" Nutrients 12, no. 4: 987. https://doi.org/10.3390/nu12040987
APA StyleMeyer, S., Schäfer, L., Röhrig, J., Maheshwari, G., Most, E., Zorn, H., Ringseis, R., Eder, K., & Gessner, D. K. (2020). Supplementation of Sulfur-Containing Amino Acids or Essential Amino Acids Does Not Reverse the Hepatic Lipid-Lowering Effect of a Protein-Rich Insect Meal in Obese Zucker Rats. Nutrients, 12(4), 987. https://doi.org/10.3390/nu12040987