MK-7 and Its Effects on Bone Quality and Strength

Abstract
1. Introduction
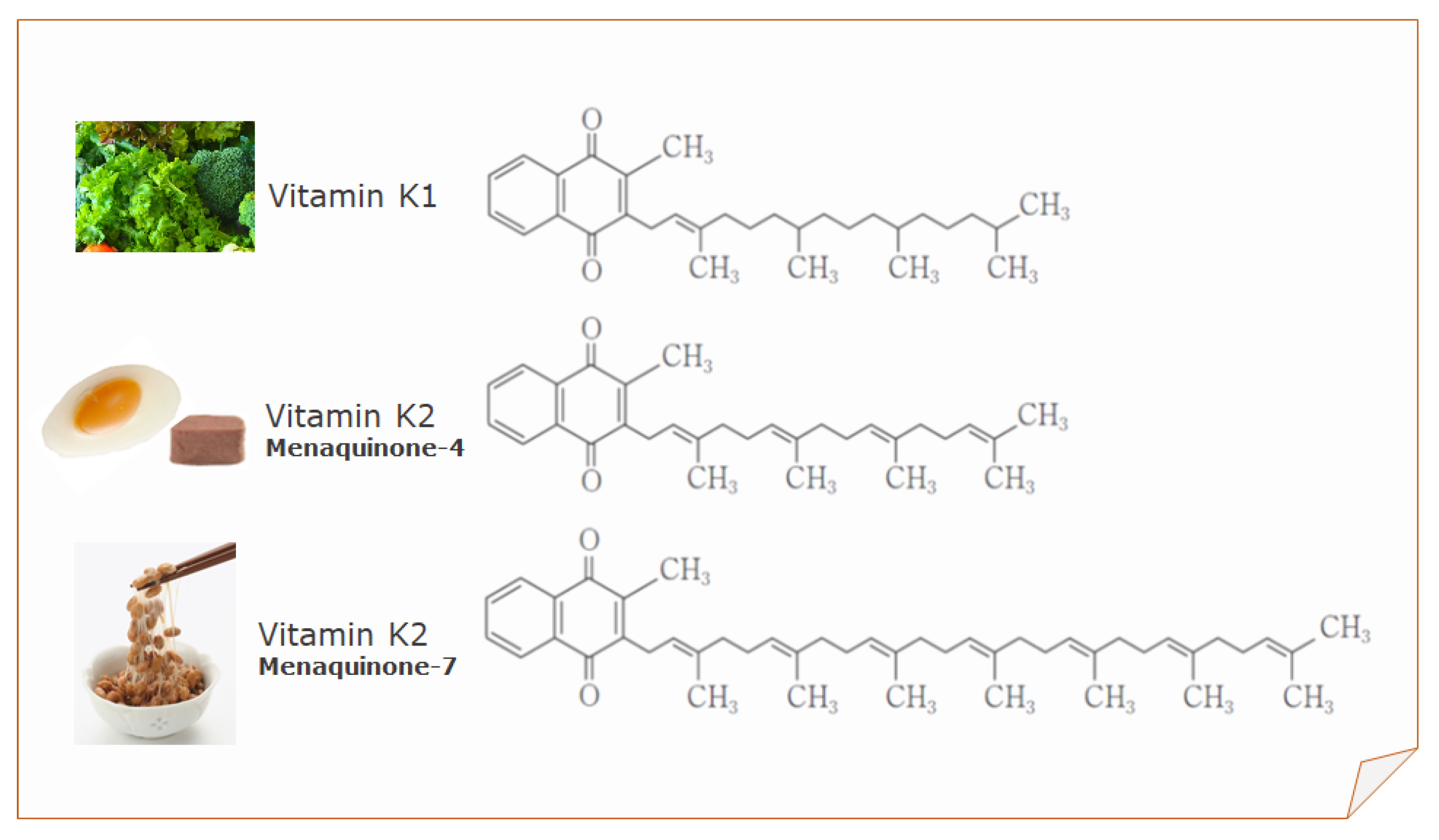
2. Sources of Vitamin K
3. Daily Requirement of Vitamin K
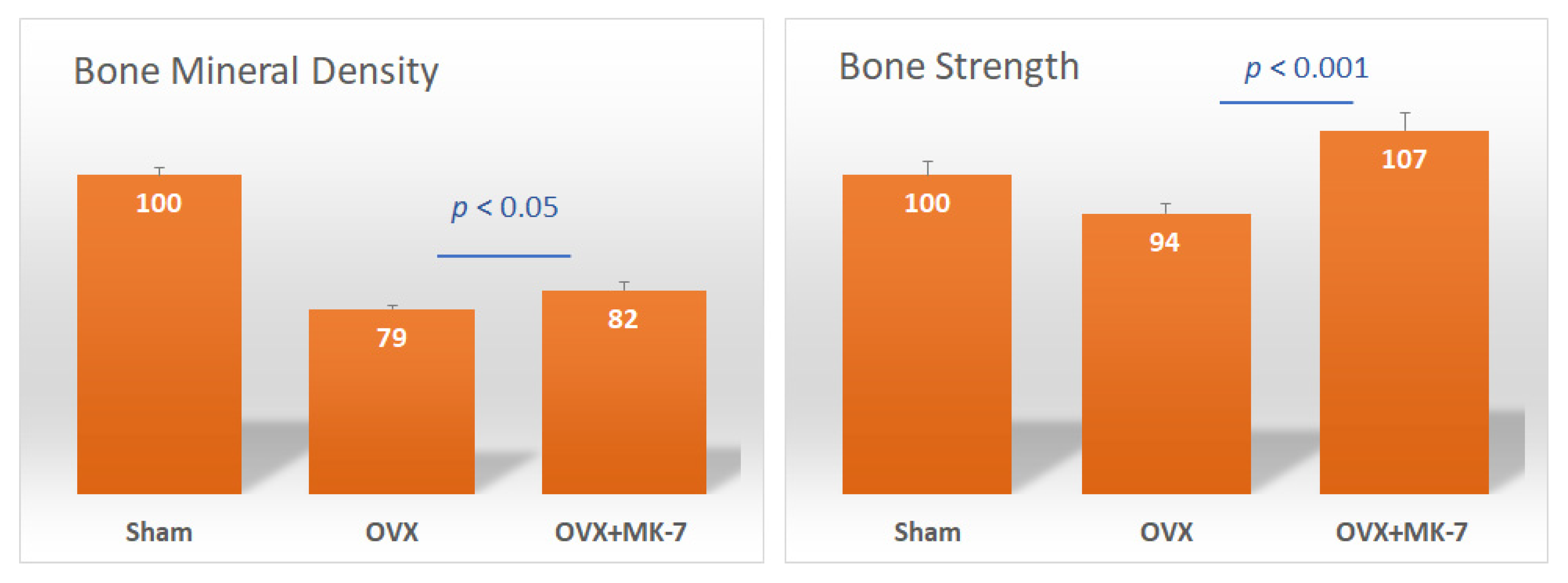
4. MK-7 and Bone Quality
5. Advantages of MK-7
6. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shearer, M.J. Vitamin K and vitamin K-dependent proteins. Br. J. Haematol. 1990, 75, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Furie, B.; Furie, B.C. The molecular basis of blood coagulation. Cell 1988, 53, 505–518. [Google Scholar] [CrossRef]
- Price, P.A. Role of vitamin-K-dependent proteins in bone metabolism. Annu. Rev. Nutr. 1988, 8, 565–583. [Google Scholar] [CrossRef] [PubMed]
- Wei, F.F.; Trenson, S.; Verhamme, P.; Vermeer, C.; Staessen, J.A. Vitamin K-dependent matrix Gla protein as multifaceted protector of vascular and tissue integrity. Hypertension 2019, 73, 1160–1169. [Google Scholar] [CrossRef]
- Cancela, M.L.; Conceico, N.; Laize, V. Gla-rich protein, a new player in tissue calcification? Adv. Nutr. 2012, 3, 174–181. [Google Scholar] [CrossRef]
- Coutu, D.L.; Wu, J.H.; Monette, A.; Rivard, G.E.; Blostein, M.D.; Galipeau, J. Periostin, a member of a novel family of vitamin K-dependent proteins, is expressed by mesenchymal stromal cells. J. Biol. Chem. 2008, 283, 17991–18001. [Google Scholar] [CrossRef]
- Ferland, G. Vitamin K and the nervous system: An overview of its actions. Adv. Nutr. 2012, 3, 204–212. [Google Scholar] [CrossRef]
- Laurance, S.; Lemarie, C.A.; Blostein, M.D. Growth arrest-specific gene 6 (gas6) and vascular hemostasis. Adv. Nutr. 2012, 3, 196–203. [Google Scholar] [CrossRef]
- Kulman, J.D.; Harris, J.E.; Xie, L.; Davie, E.W. Proline-rich Gla protein 2 is a cell-surface vitamin K-dependent protein that binds to the transcriptional coactivator Yes-associated protein. Proc. Natl. Acad. Sci. USA 2007, 104, 8767–8772. [Google Scholar] [CrossRef]
- Yazicioglu, M.N.; Monaldini, L.; Chu, K.; Khazi, F.R.; Murphy, S.L.; Huang, H.; Margaritis, P.; High, K.A. Cellular localization and characterization of cytosolic binding partners for Gla domain-containing proteins PRRG4 and PRRG2. J. Biol. Chem. 2013, 288, 25908–25914. [Google Scholar] [CrossRef]
- Rishavy, M.A.; Berkner, K.L. Vitamin K oxygenation, glutamate carboxylation, and processivity: Defining the three critical facets of catalysis by the vitamin K-dependent carboxylase. Adv. Nutr. 2012, 3, 135–148. [Google Scholar] [CrossRef] [PubMed]
- Willems, B.A.; Vermeer, C.; Reutelingsperger, C.P.; Schurgers, L.J. The realm of vitamin K dependent proteins: Shifting from coagulation toward calcification. Mol. Nutr. Food Res. 2014, 58, 1620–1635. [Google Scholar] [CrossRef] [PubMed]
- Halder, M.; Petsophonsakul, P.; Akbulut, A.C.; Pavlic, A.; Bohan, F.; Anderson, E.; Maresz, K.; Kramann, R.; Schurgers, L. Vitamin K: Double bonds beyond coagulation insights into differences between vitamin K1 and K2 in health and disease. Int. J. Mol. Sci. 2019, 20, 896. [Google Scholar] [CrossRef] [PubMed]
- Tabb, M.M.; Sun, A.; Zhou, C.; Grün, F.; Errandi, J.; Romero, K.; Pham, H.; Inoue, S.; Mallick, S.; Lin, M.; et al. Vitamin K2 regulation of bone homeostasis is mediated by the steroid and xenobiotic receptor SXR. J. Biol. Chem. 2003, 278, 43919–43927. [Google Scholar] [CrossRef] [PubMed]
- Azuma, K.; Ouchi, Y.; Inoue, S. Vitamin K: Novel molecular mechanisms of action and its roles in osteoporosis. Geriatr. Gerontol. Int. 2014, 14, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Booth, S.L.; Suttie, J.W. Dietary intake and adequacy of vitamin K. J. Nutr. 1998, 128, 785–788. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Okamoto, M. Production of hen’s eggs rich in vitamin K. Nutr. Res. 1997, 17, 1607–1615. [Google Scholar] [CrossRef]
- Terachi, T.; Inoue, Y.; Ashihara, N.; Kobayashi, M.; Ando, K.; Matsui, T. Plasma vitamin K concentration in horses supplemented with several vitamin K homologs. J. Anim. Sci. 2011, 89, 1056–1061. [Google Scholar] [CrossRef]
- Sakano, T.; Notsumoto, S.; Nagaoka, T.; Morimoto, A.; Fujimoto, K.; Masuda, S.; Suzuki, Y.; Hirauchi, K. Measurement of K vitamins in food by high performance liquid chromatography with fluorometric detection. Vitamins 1988, 62, 393–398. [Google Scholar]
- Schurgers, L.J.; Vermeer, C. Determination of phylloquinone and menaquinones in food. Effect of food matrix on circulating vitamin K concentrations. Haemostasis 2000, 30, 298–307. [Google Scholar] [CrossRef]
- Collins, M.D.; Jones, D. Distribution of isoprenoid quinone structural types in bacteria and their taxonomic implications. Microbiol. Rev. 1981, 45, 316–354. [Google Scholar] [CrossRef] [PubMed]
- Conly, J.M.; Stein, K. Quantitative and qualitative measurements of K vitamins in human intestinal contents. Am. J. Gastroenterol. 1992, 87, 311–316. [Google Scholar] [PubMed]
- Uchida, K.; Nomura, Y.; Takase, H.; Harauchi, T.; Yoshizaki, T.; Nakao, H. Effects of vitamin K-deficient diets and fasting on blood coagulation factors in conventional and germfree rats. Jpn. J. Pharmacol. 1986, 40, 115–122. [Google Scholar] [CrossRef]
- Will, B.H.; Suttie, J.W. Comparative metabolism of phylloquinone and menaquinone-9 in rat liver. J. Nutr. 1992, 122, 953–958. [Google Scholar] [CrossRef] [PubMed]
- Suttie, J.W.; Mummah-Schendel, L.L.; Shah, D.V.; Lyle, B.J.; Greger, J.L. Vitamin K deficiency from dietary vitamin K restriction in humans. Am. J. Clin. Nutr. 1988, 47, 475–480. [Google Scholar] [CrossRef] [PubMed]
- Ferland, G.; Sadowski, J.A.; O’Brien, M.E. Dietary induced subclinical vitamin K deficiency in normal human subjects. J. Clin. Investig. 1993, 91, 1761–1768. [Google Scholar] [CrossRef]
- Beulens, J.W.J.; Booth, S.L.; van den Heuvel, E.G.H.M.; Stoecklin, E.; Baka, A.; Vermeer, C. The role of menaquinones (vitamin K2) in human health. Br. J. Nutr. 2013, 110, 1357–1368. [Google Scholar] [CrossRef]
- Marles, R.J.; Roe, A.L.; Oketch-Rabah, H.A. US Pharmacopeial Convention safety evaluation of menaquinone-7, a form of vitamin K. Nutr. Rev. 2017, 75, 553–578. [Google Scholar] [CrossRef]
- National Research Council. Dietary Reference Intakes for Vitamin A, Vitamin K, Arsenic, Boron, Chromium, Copper, Iodine, Iron, Manganese, Molybdenum, Nickel, Silicon, Vanadium, and Zinc; National Academy Press: Washington, DC, USA, 2000; pp. 162–196. [Google Scholar] [CrossRef]
- World Health Organization and Food and Agriculture Organization of the United Nations. Vitamin K. In Vitamin and Mineral Requirements in Human Nutrition, 2nd ed.; World Health Organization: Geneva, Switzerland, 2004; pp. 108–129. [Google Scholar] [CrossRef]
- European Community. Commission Directive 2008/100/EC of 28 October 2008 Amending Council Directive 90/496/EEC on Nutrition Labelling for Food Stuffs as Regards Recommended Daily Allowances, Energy Conversion Factors and Definitions. Off. J. Eur. Union 2008, 285, 9. [Google Scholar]
- Tanaka, K.; Terao, J.; Shidoji, Y.; Tamai, H.; Imai, E.; Okano, T. Dietary reference intake for Japanese 2010: Fat-Soluble Vitamins. J. Nutr. Sci. Vitaminol. 2013, 59, S57–S66. [Google Scholar] [CrossRef]
- Cranenburg, E.C.; Schurgers, L.J.; Vermeer, C. Vitamin K: The coagulation vitamin that became omnipotent. Thromb. Haemost. 2007, 98, 120–125. [Google Scholar] [CrossRef] [PubMed]
- McCann, J.C.; Ames, B.N. Vitamin K, an example of triage theory: Is micronutrient inadequacy linked to diseases of aging? Am. J. Clin. Nutr. 2009, 90, 889–907. [Google Scholar] [CrossRef] [PubMed]
- Inaba, N.; Sato, T.; Yamashita, T. Low-dose daily intake of vitamin K2 (Menaquinone-7) improves osteocalcin γ-carboxylation: A double-blind, randomized controlled trial. J. Nutr. Sci. Vitaminol. 2015, 61, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Shearer, M.J.; Bach, A.; Kohlmeier, M. Chemistry, nutritional sources, tissue distribution and metabolism of vitamin K with special reference to bone health. J. Nutr. 1996, 126, 1181S–1186S. [Google Scholar] [CrossRef]
- Schurgers, L.J.; Vermeer, C. Differential lipoprotein transport pathways of K-vitamins in healthy subjects. Biochim. Biophys. Acta 2002, 1570, 27–32. [Google Scholar] [CrossRef]
- Theuwissen, E.; Cranenburg, E.C.; Knapen, M.H.; Magdeleyns, E.J.; Teunissen, K.J.; Schurgers, L.J.; Smit, E.; Vermeer, C. Low-dose menaquinone-7 supplementation improved extra-hepatic vitamin K status, but had no effect on thrombin generation in healthy subjects. Br. J. Nutr. 2012, 108, 1652–1657. [Google Scholar] [CrossRef]
- Knapen, M.H.; Drummen, N.E.; Smit, E.; Vermeer, C.; Theuwissen, E. Three-year low-dose menaquinone-7 supplementation helps decrease bone loss in healthy postmenopausal women. Osteoporos Int. 2013, 24, 2499–2507. [Google Scholar] [CrossRef]
- Knapen, M.H.J.; Braam, L.A.J.L.M.; Drummen, N.E.; Bekers, O.; Hoeks, A.P.G.; Vermeer, C. Menaquinone-7 supplementation improves arterial stiffness in healthy postmenopausal women. A double-blind randomised clinical trial. Thromb. Haemost. 2015, 113, 1135–1144. [Google Scholar] [CrossRef]
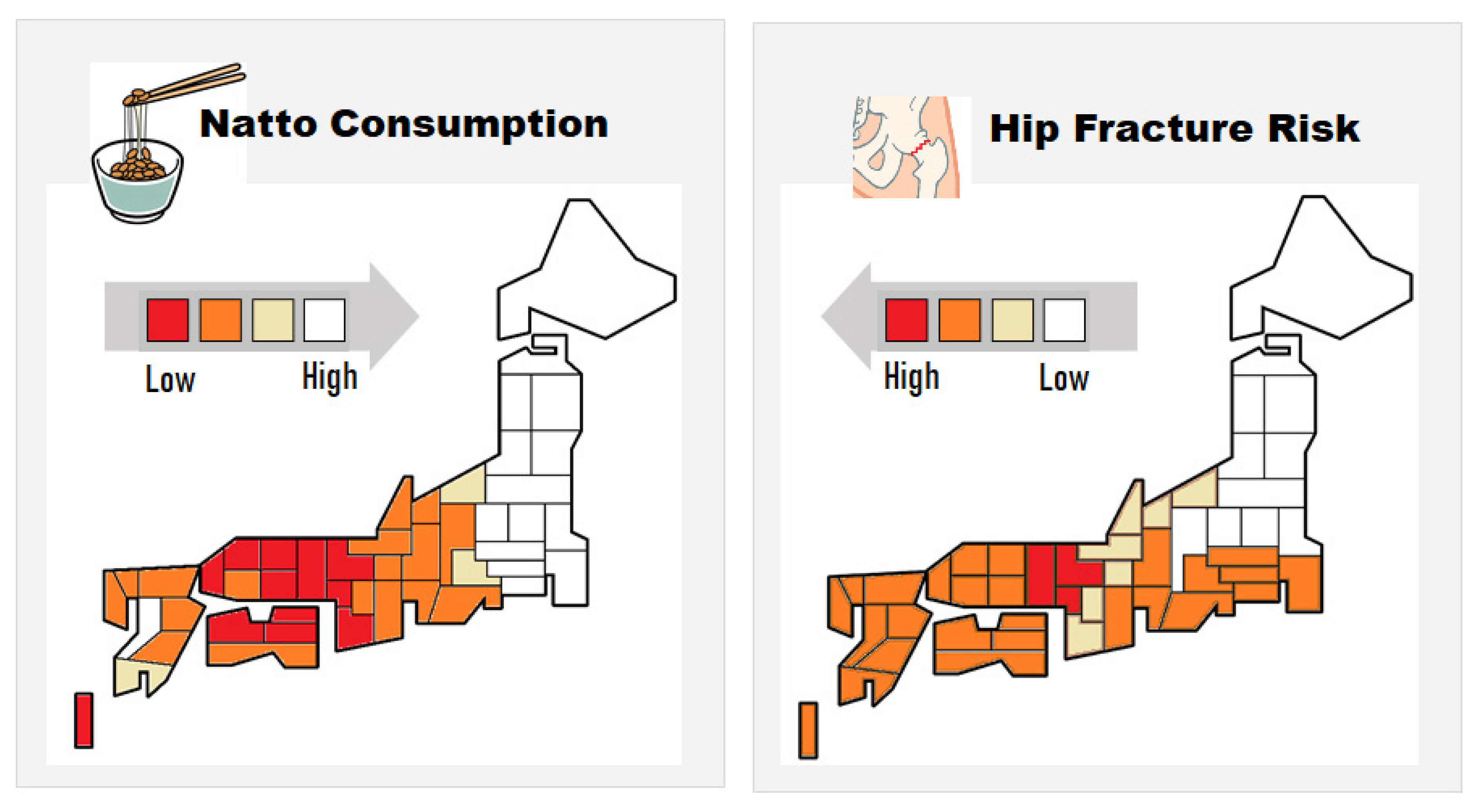
- Kaneki, M.; Hodges, S.J.; Hosoi, T.; Fujiwara, S.; Lyons, A.; Crean, S.J.; Ishida, N.; Nakagawa, M.; Takechi, M.; Sano, Y.; et al. Japanese fermented soybean food as the major determinant of the large geographic difference in circulating concentrations of vitamin K2: Possible implications for hip-fracture risk. Nutrition 2001, 17, 315–321. [Google Scholar] [CrossRef]
- Yaegashi, Y.; Onoda, T.; Tanno, K.; Kuribayashi, T.; Sakata, K.; Orimo, H. Association of hip fracture incidence and intake of calcium, magnesium, vitamin D., and vitamin K. Eur. J. Epidemiol. 2008, 23, 219–225. [Google Scholar] [CrossRef]
- Kojima, A.; Ikehara, S.; Kamiya, K.; Kajita, E.; Sato, Y.; Kouda, K.; Tamaki, J.; Kagamimori, S.; Iki, M. Natto intake is inversely associated with osteoporotic fracture risk in postmenopausal Japanese women. J. Nutr. 2019, 150, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.; Moreines, J.; Booth, S.L. Vitamin K supplementation does not prevent bone loss in ovariectomized Norway Rats. Nutr. Metab. 2012, 9, 12. [Google Scholar] [CrossRef] [PubMed]
- Murasawa, H.; Araki, S.; Mitsui, M.; Shirota, S.; Ikeda, R. Studies on high-vitamin K containing natto and its effect on osteoporosis. In Shokuhinsozai no Kinousei Souzou Seigyo Gijutsu, 1st ed.; The Japanese Research and Development Association for New Functional Foods, Koseisha Koseikaku: Tokyo, Japan, 1999; pp. 131–146. [Google Scholar]
- Knapen, M.H.; Schurgers, L.J.; Vermeer, C. Vitamin K2 supplementation improves hip bone geometry and bone strength indices in postmenopausal women. Osteoporosis Int. 2007, 18, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Ichikawa, T.; Horie-Inoue, K.; Ikeda, K.; Blumberg, B.; Inoue, S. Steroid and xenobiotic receptor SXR mediates vitamin K2-activated transcription of extracellular matrix-related genes and collagen accumulation in osteoblastic cells. J. Biol. Chem. 2006, 281, 16927–16934. [Google Scholar] [CrossRef]
- Sato, T. Vitamin K2 and Bone Quality. Vitam. Trace Elem. 2012, S6. [Google Scholar] [CrossRef]
- Maillard, C.; Berruyer, M.; Serre, C.M.; Dechavanne, M.; Delmas, P.D. Protein-S, a vitamin K-dependent protein, is a bone matrix component synthesized and secreted by osteoblasts. Endocrinology 1992, 130, 1599–1604. [Google Scholar] [CrossRef]
- Seibel, M.J.; Robins, S.P.; Bilezikian, J.P. Serum undercarboxylated osteocalcin and the risk of hip fracture. J. Clin. Endorcinol. Metab. 1997, 82, 717–718. [Google Scholar] [CrossRef]
- Vergnaud, P.; Garnero, P.; Meunier, P.J.; Bréart, G.; Kamihagi, K.; Delmas, P.D. Undercarboxylated osteocalcin measured with a specific immunoassay predicts hip fracture in elderly women: The EPIDOS study. J. Clin. Endocrinol. Metab. 1997, 82, 719–724. [Google Scholar] [CrossRef]
- Binkley, N.C.; Krueger, D.C.; Kawahara, T.N.; Engelke, J.A.; Chappell, R.J.; Suttie, J.W. A high phylloquinone intake is required to achieve maximal osteocalcin gamma-carboxylation. Am. J. Clin. Nutr. 2002, 76, 1055–1060. [Google Scholar] [CrossRef]
- Takeuchi, A.; Masuda, Y.; Kimura, M.; Marushima, R.; Matsuoka, R.; Hasegawa, M.; Takahara, M.; Onuki, M. Minimal effective dose of vitamin K2 (menaquinone-4) on serum osteocalcin concentration in Japanese subjects and safety evaluation of vitamin K2 supplemented in calcium tablet. J. Jpn. Soc. Clin. Nutr. 2005, 26, 254–260. [Google Scholar]
- Nakamura, E.; Aoki, M.; Watanabe, F.; Kamimura, A. Low-dose menaquinone-4 improves γ-carboxylation of osteocalcin in young males: A non-placebo-controlled dose–response study. Nutr. J. 2014, 13, 85. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Sato, T.; Schurgers, L.J.; Uenishi, K. Comparison of menaquinone-4 and menaquinone-7 bioavailability in Japanese. Nutr. J. 2012, 11, 93. [Google Scholar] [CrossRef]
- Brugè, F.; Bacchetti, T.; Principi, F.; Littarru, G.P.; Tiano, L. Olive oil supplemented with menaquinone-7 significantly affects osteocalcin carboxylation. Br. J. Nutr. 2011, 106, 1058–1062. [Google Scholar] [CrossRef] [PubMed]
- Schurgers, L.J.; Teunissen, K.J.; Hamulyak, K.; Knapen, M.H.; Vik, H.; Vermeer, C. Vitamin K-containing dietary supplements: Comparison of synthetic vitamin K1 and natto-derived menaquinone-7. Blood 2007, 109, 3279–3283. [Google Scholar] [CrossRef] [PubMed]
- Komai, M.; Shirakawa, H. Vitamin K metabolism. Menaquinone-4 (MK-4) formation from ingested VK analogues and its potent relation to bone function. Clin. Calcium. 2007, 17, 1663–1672. [Google Scholar] [PubMed]
- Nakagawa, K.; Hirota, Y.; Sawada, N.; Yuge, N.; Watanabe, M.; Uchino, Y.; Okuda, N.; Shimomura, Y.; Suhara, Y.; Okano, T. Identification of UBIAD1 as a novel human menaquinone-4 biosynthetic enzyme. Nature 2010, 468, 117–121. [Google Scholar] [CrossRef]
- Sato, T.; Kawahara, R.; Kamo, S.; Saito, S. Comparison of menaquinone-4 and menaquinone-7 in rats. Vitamins 2007, 81, 377–381. [Google Scholar]
- Yamaguchi, M.; Sugimoto, E.; Hachiya, S. Stimulatory effect of menaquinone-7 (vitamin K2) on osteoblastic bone formation in vitro. Mol. Cell. Biochem. 2001, 223, 131–137. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Ma, Z.J. Inhibitory effect of menaquinone-7 (vitamin K2) on osteoclast-like cell formation and osteoclastic bone resorption in rat bone tissues in vitro. Mol. Cell. Biochem. 2001, 228, 39–47. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Weitzmann, M.N. Vitamin K2 stimulates osteoblastogenesis and suppresses osteoclastogenesis by suppressing NF-κB activation. Int. J. Mol. Med. 2011, 27, 3–14. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | Function | Ref |
|---|---|---|
| Factors II (Prothrombin), VII, IX, X | Procoagulants | [1,2] |
| Proteins C, S, Z | Anticoagulants | [1,2] |
| Osteocalcin | Regulator of mineral deposition | [3] |
| Matrix γ-carboxyglutamic acid protein | Inhibition of ectopic calcification | [4] |
| γ-carboxyglutamic acid-rich protein | Inhibition of ectopic calcification, anti-inflammatory | [5] |
| Periostin | Inhibition of ectopic calcification, tissue regeneration | [6] |
| Growth arrest-specific protein 6 | Cell proliferation | [7,8] |
| Proline-rich γ-carboxy glutamyl proteins 1 and 2 | Not well-known | [9,10] |
| γ-glutamyl carboxylase | γ-glutamyl carboxylation of vitamin K-dependent proteins | [11] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sato, T.; Inaba, N.; Yamashita, T. MK-7 and Its Effects on Bone Quality and Strength. Nutrients 2020, 12, 965. https://doi.org/10.3390/nu12040965
Sato T, Inaba N, Yamashita T. MK-7 and Its Effects on Bone Quality and Strength. Nutrients. 2020; 12(4):965. https://doi.org/10.3390/nu12040965
Chicago/Turabian StyleSato, Toshiro, Naoko Inaba, and Takatoshi Yamashita. 2020. "MK-7 and Its Effects on Bone Quality and Strength" Nutrients 12, no. 4: 965. https://doi.org/10.3390/nu12040965
APA StyleSato, T., Inaba, N., & Yamashita, T. (2020). MK-7 and Its Effects on Bone Quality and Strength. Nutrients, 12(4), 965. https://doi.org/10.3390/nu12040965