Changes in Salivary Proteome in Response to Bread Odour

 ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Material and Methods
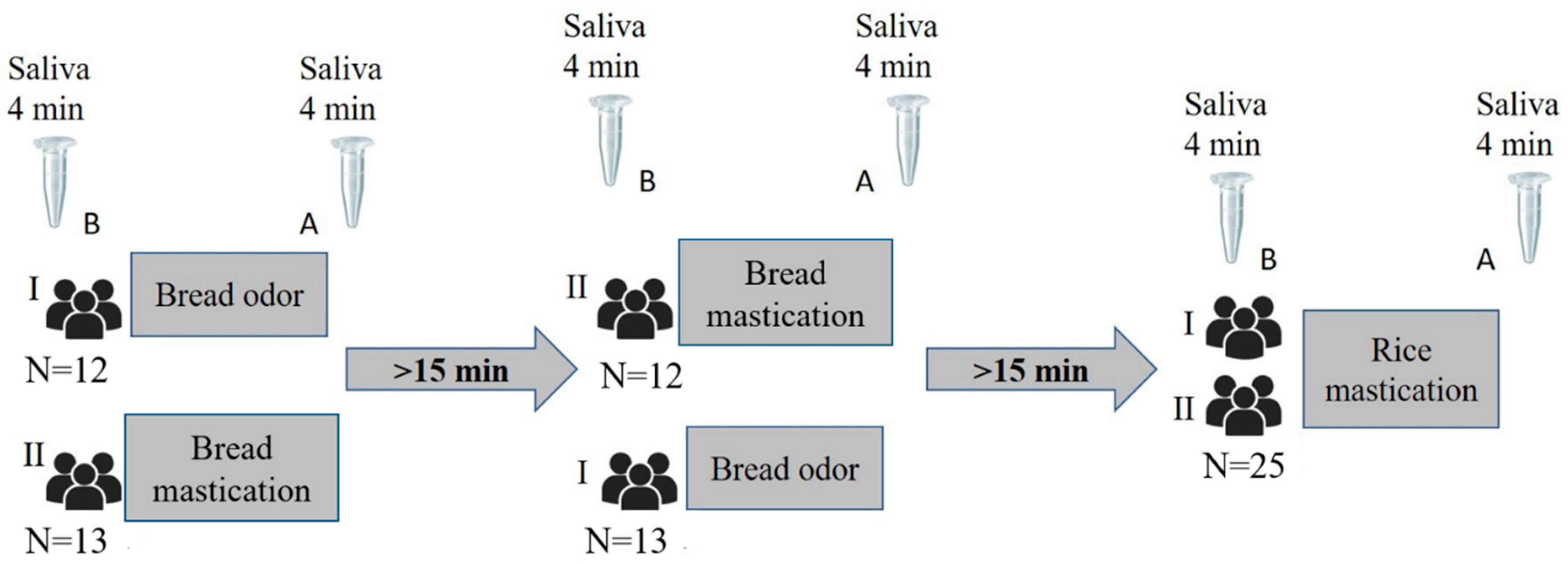
2.1. Participants
2.2. Stimulation and Saliva Collection
2.3. Salivary Flow Rate and Total Protein Quantification
2.4. Salivary Amylase Enzymatic Activity
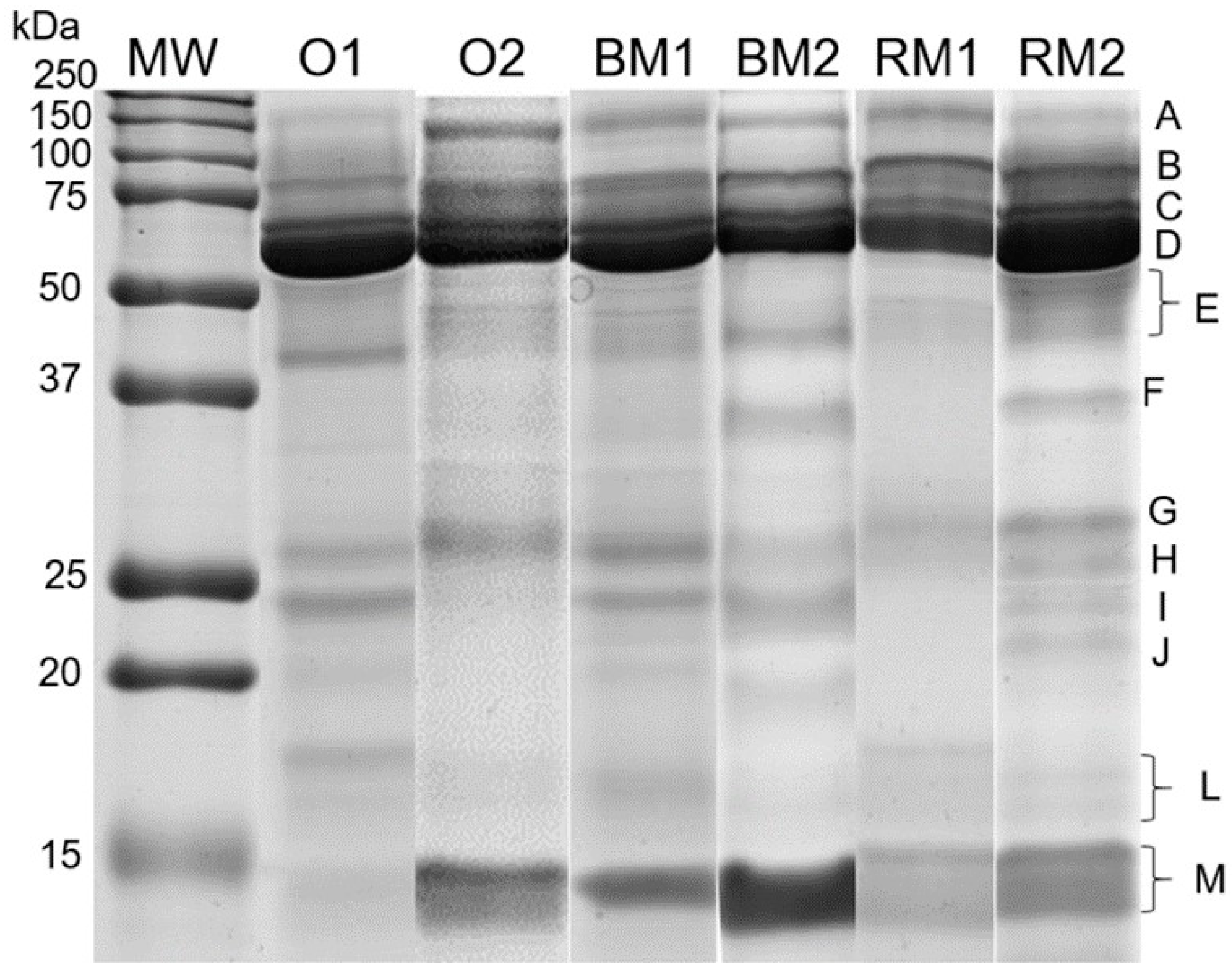
2.5. Sodium Dodecyl Sulphate-Polyacrylamide Gel Electrophoresis (SDS-PAGE) Salivary Protein Separation and Protein Band Identification
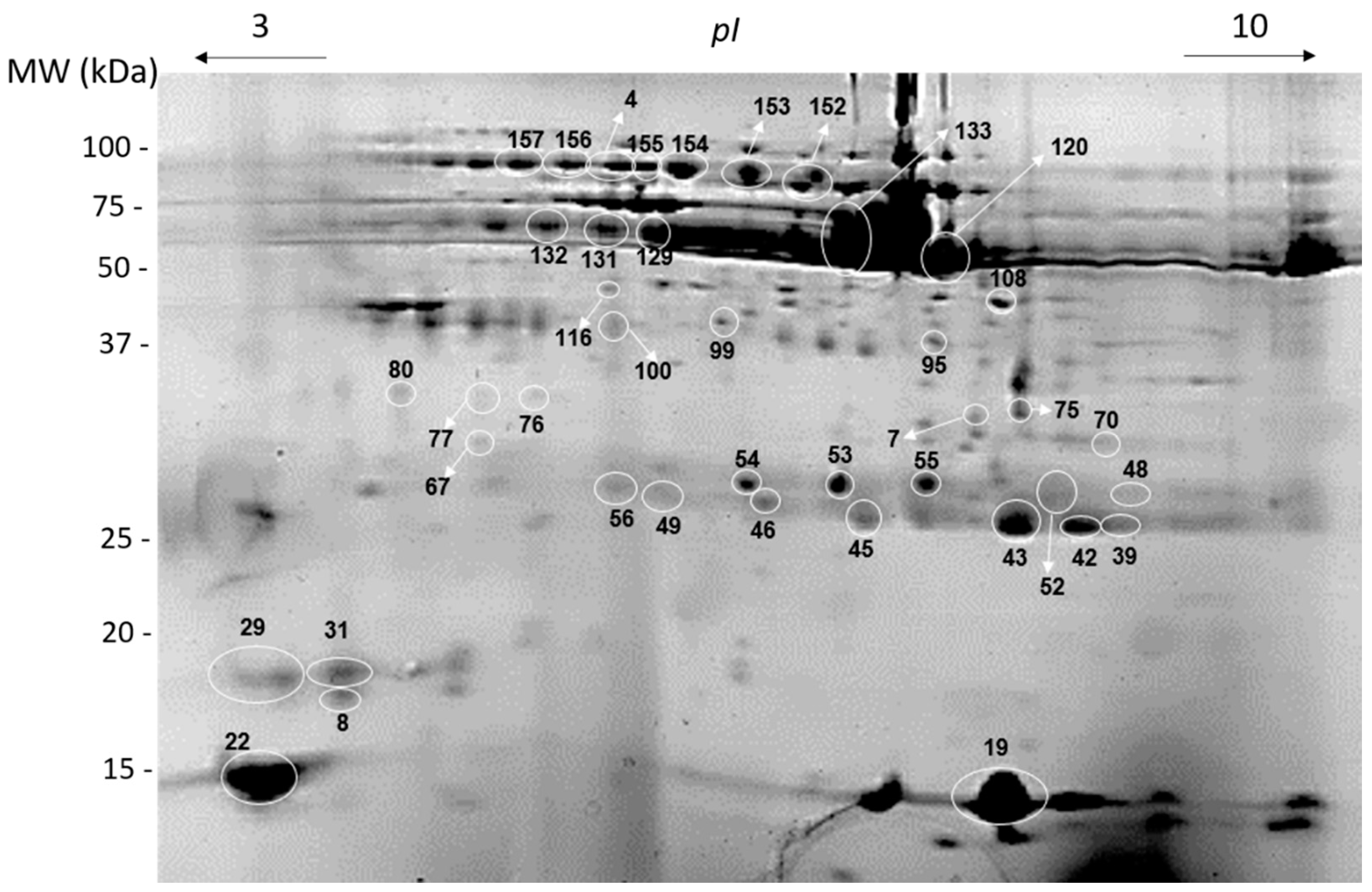
2.6. Two-Dimensional Electrophoresis
2.7. Statistical Analysis
3. Results
3.1. Salivary Flow Rate, Total Protein Concentration and Amylase Enzymatic Activity
3.2. Salivary SDS-PAGE Protein Profile
3.3. Salivary Two-Dimensional Electrophoretic (2-DE) Profile
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Van Dongen, M.V.; Van Den Berg, M.C.; Vink, N.; Kok, F.J.; De Graaf, C. Taste-Nutrient relationships in commonly consumed foods. Br. J. Nutr. 2012, 108, 140–147. [Google Scholar] [CrossRef] [PubMed]
- Sclafani, A. Learned controls of ingestive behaviour. Appetite 1997, 29, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Yeomans, M.R. Olfactory influences on appetite and satiety in humans. Physiol. Behav. 2006, 89, 10–14. [Google Scholar] [CrossRef] [PubMed]
- Ferriday, D.; Brunstrom, J.M. I just can’t help myself: Effects of food-cue exposure in overweight and lean individuals. Int. J. Obes. 2011, 35, 142–149. [Google Scholar] [CrossRef] [PubMed]
- Ramaekers, M.G.; Luning, P.A.; Lakemond, C.M.M.; Van Boekel, M.A.J.S.; Gort, G.; Boesveldt, S. Food preference and appetite after switching between sweet and savoury odours in women. PLoS ONE 2016, 11, e0146652. [Google Scholar] [CrossRef]
- Ramaekers, M.G.; Boesveldt, S.; Lakemond, C.M.M.; Van Boekel, M.A.J.S.; Luning, P.A. Odors: Appetizing or satiating development of appetite during odor exposure over time. Int. J. Obes. 2014, 38, 650–656. [Google Scholar] [CrossRef]
- Ramaekers, M.G.; Boesveldt, S.; Gort, G.; Lakemond, C.M.M.; van Boekel, M.A.J.S.; Luning, P.A. Sensory-Specific appetite is affected by actively smelled food odors and remains stable over time in normal-weight women. J. Nutr. 2014, 144, 1314–1319. [Google Scholar] [CrossRef]
- Smeets, P.A.M.; Erkner, A.; De Graaf, C. Cephalic phase responses and appetite. Nutr. Rev. 2010, 68, 643–655. [Google Scholar] [CrossRef]
- Proserpio, C.; de Graaf, C.; Laureati, M.; Pagliarini, E.; Boesveldt, S. Impact of ambient odors on food intake, saliva production and appetite ratings. Physiol. Behav. 2017, 174, 35–41. [Google Scholar] [CrossRef]
- Morquecho-Campos, P.; Bikker, F.J.; Nazmi, K.; de Graaf, K.; Laine, M.L.; Boesveldt, S. Impact of food odors signaling specific taste qualities and macronutrient content on saliva secretion and composition. Appetite 2019, 143, 104399. [Google Scholar] [CrossRef] [PubMed]
- Soares, S.; Mateus, N.; de Freitas, V. Interaction of different classes of salivary proteins with food tannins. Food Res. Int. 2012, 49, 807–813. [Google Scholar] [CrossRef]
- Soares, S.; García-Estévez, I.; Ferrer-Galego, R.; Brás, N.F.; Brandão, E.; Silva, M.; Teixeira, N.; Fonseca, F.; Sousa, S.F.; Ferreira-da-Silva, F.; et al. Study of human salivary proline-rich proteins interaction with food tannins. Food Chem. 2018, 243, 175–185. [Google Scholar] [CrossRef] [PubMed]
- Lamy, E.; Mowe, M.; Pinheiro, C.; Rodrigues, L.; Lopes, O.; Capela e Silva, F. Oral food perception and polyphenol-rich foods acceptance—The importance of knowing individuals saliva characteristics for promoting consumption. J. Int. Soc. Antioxid. 2016, 3. [Google Scholar] [CrossRef]
- Martin, L.E.; Kay, K.E.; Torregrossa, A.-M. Bitter-Induced salivary proteins increase detection threshold of quinine, but not sucrose. Chem. Senses 2019, 44, 379–388. [Google Scholar] [CrossRef]
- Rodrigues, L.; da Costa, G.; Cordeiro, C.; Pinheiro, C.C.; Amado, F.; Lamy, E. Relationship between saliva protein composition and 6-n-Propylthiouracil bitter taste responsiveness in young adults. J. Sens. Stud. 2017, 32, e12275. [Google Scholar] [CrossRef]
- Cabras, T.; Melis, M.; Castagnola, M.; Padiglia, A.; Tepper, B.J.; Messana, I.; Tomassini Barbarossa, I. Responsiveness to 6-n-propylthiouracil (PROP) is associated with salivary levels of two specific basic proline-rich proteins in humans. PLoS ONE 2012, 7, e30962. [Google Scholar] [CrossRef]
- Dsamou, M.; Palicki, O.; Septier, C.; Chabanet, C.; Lucchi, G.; Ducoroy, P.; Chagnon, M.-C.; Morzel, M. Salivary protein profiles and sensitivity to the bitter taste of caffeine. Chem. Senses 2012, 37, 87–95. [Google Scholar] [CrossRef]
- Rodrigues, L.; Costa, G.; Cordeiro, C.; Pinheiro, C.; Amado, F.; Lamy, E. Salivary proteome and glucose levels are related with sweet taste sensitivity in young adults. Food Nutr. Res. 2017, 61, 1389208. [Google Scholar] [CrossRef]
- Lamy, E.; Simões, C.; Rodrigues, L.; Costa, A.R.; Vitorino, R.; Amado, F.; Antunes, C.; do Carmo, I. Changes in the salivary protein profile of morbidly obese women either previously subjected to bariatric surgery or not. J. Physiol. Biochem. 2015, 71, 691–702. [Google Scholar] [CrossRef]
- Jessie, K.; Pang, W.W.; Rahim, Z.H.A.; Hashim, O.H. Proteomic analysis of whole human saliva detects enhanced expression of interleukin-1 receptor antagonist, thioredoxin and lipocalin-1 in cigarette smokers compared to non-smokers. Int. J. Mol. Sci. 2010, 11, 4488–4505. [Google Scholar] [CrossRef]
- Rodrigues, L.; Espanca, R.; Costa, A.R.; Antunes, C.M.; Pomar, C.; Capela-Silva, F.; Pinheiro, C.C.; Domingues, P.; Amado, F.; Lamy, E. Comparison of salivary proteome of children with different sensitivities for bitter and sweet tastes: Association with body mass index. Int. J. Obes. 2019, 43, 701–712. [Google Scholar] [CrossRef] [PubMed]
- Chan, H.H.; Rahim, Z.H.A.; Jessie, K.; Hashim, O.H.; Taiyeb-Ali, T.B. Salivary proteins associated with periodontitis in patients with type 2 diabetes mellitus. Int. J. Mol. Sci. 2012, 13, 4642–4654. [Google Scholar] [CrossRef] [PubMed]
- Feldman, M.; Richardson, C.T. Role of thought, sight, smell, and taste of food in the cephalic phase of gastric acid secretion in humans. Gastroenterology 1986, 90, 428–433. [Google Scholar] [CrossRef]
- Al-Manei, K.; Almotairy, N.; Bostanci, N.; Kumar, A.; Grigoriadis, A. Effect of chewing on the expression of salivary protein composition: A systematic review. Proteom. Clin. Appl. 2019, e1900039. [Google Scholar] [CrossRef]
- Sánchez, G.A.; Miozza, V.; Delgado, A.; Busch, L. Determination of salivary levels of mucin and amylase in chronic periodontitis patients. J. Periodontal Res. 2011, 46, 221–227. [Google Scholar] [CrossRef]
- Neyraud, E.; Bult, J.H.F.; Dransfield, E. Continuous analysis of parotid saliva during resting and short-duration simulated chewing. Arch. Oral Biol. 2009, 54, 449–456. [Google Scholar] [CrossRef]
- Joubert, M.; Septier, C.; Brignot, H.; Salles, C.; Panouillé, M.; Feron, G.; Tournier, C. Chewing bread: Impact on alpha-amylase secretion and oral digestion. Food Funct. 2017, 8. [Google Scholar] [CrossRef]
- Proctor, G.B.; Carpenter, G.H. Chewing stimulates secretion of human salivary secretory immunoglobulin A. J. Dent. Res. 2001, 80, 909–913. [Google Scholar] [CrossRef]
- Mandel, A.L.; Peyrot des Gachons, C.; Plank, K.L.; Alarcon, S.; Breslin, P.A.S. Individual differences in AMY1 gene copy number, salivary α-amylase levels, and the perception of oral starch. PLoS ONE 2010, 5, e13352. [Google Scholar] [CrossRef]
- Santos, J.L.; Saus, E.; Smalley, S.V.; Cataldo, L.R.; Alberti, G.; Parada, J.; Gratacòs, M.; Estivill, X. Copy number polymorphism of the salivary amylase gene: Implications in human nutrition research. J. Nutrigenet. Nutrigenom. 2012, 5, 117–131. [Google Scholar] [CrossRef]
- Mackie, D.A.; Pangborn, R.M. Mastication and its influence on human salivary flow and alpha-amylase secretion. Physiol. Behav. 1990, 47, 593–595. [Google Scholar] [CrossRef]
- Marquezin, M.C.S.; Pedroni-Pereira, A.; Araujo, D.S.; Rosar, J.V.; Barbosa, T.S.; Castelo, P.M. Descriptive analysis of the masticatory and salivary functions and gustatory sensitivity in healthy children. Acta Odontol. Scand. 2016, 74, 443–448. [Google Scholar] [CrossRef] [PubMed]
- Kazemi, H.H.; Sefidgar, S.A.; Mortazavi-Amiri, H.; Bijani, A. The effects of dominant chewing side on amylase enzyme activities in the parotid gland. J. Babol Univ. Med. Sci. 2015, 17, 52–58. [Google Scholar]
- Jensen, J.L.; Brodin, P.; Berg, T.; Aars, H. Parotid secretion of fluid, amylase and kallikrein during reflex stimulation under normal conditions and after acute administration of autonomic blocking agents in man. Acta Physiol. Scand. 1991, 143, 321–329. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.; Linden, R. An olfactory-submandibular salivary reflex in humans. Exp. Physiol. 1992, 77, 221–224. [Google Scholar] [CrossRef] [PubMed]
- Lee, V.; Linden, R. An olfactory-parotid salivary reflex in humans? Exp. Physiol. 1991, 76, 347–355. [Google Scholar] [CrossRef] [PubMed]
- Ilangakoon, Y.; Carpenter, G.H. Is the mouthwatering sensation a true salivary reflex? J. Texture Stud. 2011, 42, 212–216. [Google Scholar] [CrossRef]
- Walz, A.; Stühler, K.; Wattenberg, A.; Hawranke, E.; Meyer, H.E.; Schmalz, G.; Blüggel, M.; Ruhl, S. Proteome analysis of glandular parotid and submandibular-sublingual saliva in comparison to whole human saliva by two-dimensional gel electrophoresis. Proteomics 2006, 6, 1631–1639. [Google Scholar] [CrossRef]
- Jasim, H.; Olausson, P.; Hedenberg-Magnusson, B.; Ernberg, M.; Ghafouri, B. The proteomic profile of whole and glandular saliva in healthy pain-free subjects. Sci. Rep. 2016, 6, 39073. [Google Scholar] [CrossRef] [PubMed]
- Crystal, S.R.; Teff, K.L. Tasting fat: Cephalic phase hormonal responses and food intake in restrained and unrestrained eaters. Physiol. Behav. 2006, 89, 213–220. [Google Scholar] [CrossRef]
- Carpenter, G.H. The secretion, components, and properties of saliva. Annu. Rev. Food Sci. Technol. 2013, 4, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Brandtzaeg, P. Secretory immunity with special reference to the oral cavity. J. Oral Microbiol. 2013, 5. [Google Scholar] [CrossRef] [PubMed]
- Schiffman, S.S. Effect of taste and smell on secretion rate of salivary iga in elderly and young persons. J. Nutr. Health Aging 1999, 3, 158–164. [Google Scholar] [PubMed]
- Allgrove, J.E.; Oliveira, M.; Gleeson, M. Stimulating whole saliva affects the response of antimicrobial proteins to exercise. Scand. J. Med. Sci. Sports 2014, 24, 649–655. [Google Scholar] [CrossRef][Green Version]
- Brandtzaeg, P. The secretory immune system of lactating human mammary glands compared with other exocrine exocrine organs. Ann. N. Y. Acad. Sci. 1983, 409, 353–382. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Salivary Parameter | Bread Odour | Bread Chewing | Rice Chewing | |||
|---|---|---|---|---|---|---|
| Before | After | Before | After | Before | After | |
| Secretion rate (mL/min) (N = 25) | 0.55 ± 0.33 a | 0.61 ± 0.37 b | 0.52 ± 0.27 a | 0.74 ± 0.40 b | 0.63 ± 0.31 a | 0.82 ± 0.44 b |
| Protein concentration (µg/mL) (N = 25) | 657.2 ± 412.9 a | 459.7 ± 201.5 b | 612.1 ± 307.0 a | 621.4 ± 240.4 a | 498.9 ± 225.8 a | 469.7 ± 236.1 a |
| α-amylase (U/L) (N = 24) | 126.6 ± 94.0 | 121.3 ± 97.8 | 135.1 ± 99.7 | 161.9 ± 87.1 | 133.9 ± 102.8 | 121.6 ± 106.0 |
| Protein Band | Protein | Assession Number (Uniprot) | MW (kDa) (Est/Theor.) # | Mascot ID Score | Sequence Coverage | Bread Odour | Bread Mastication | Rice Mastication | Interaction Period * Treatment p-Value | |||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Before | After | Before | After | Before | After | |||||||
| A | Ig polymeric receptor + Lactotransferrin | P01833 | 125.0/84.4 | 107 | 21 | 5.32 ± 1.23 a | 6.04 ± 1.63 b | 6.02 ± 1.09 a | 4.76 ± 1.39 b | 5.55 ± 1.45 a | 4.39 ± 1.29 b | <0.001 * |
| P02788 | 125.0/80.0 | 89 | 20 | |||||||||
| B | Serum albumin | P02768 | 71.0/71.3 | 109 | 19 | 8.86 ± 2.26 | 8.75 ± 2.48 | 8.91 ± 2.24 a | 7.78 ± 2.33 b | 8.12 ± 2.75 a | 7.00 ± 2.12 b | 0.457 |
| C | α-Amylase 1 | P04745 | 66.0/58.4 | 154 | 43 | 9.57 ± 2.75 | 9.81 ± 3.33 | 10.61 ± 3,69 | 8.85 ± 3,76 | 9.83 ± 3.23 | 8.68 ± 2.36 | 0.287 |
| D | α-Amylase 1 | P04745 | 60.0/58.4 | 100 | 21 | 14.90 ± 4.95 | 13.92 ± 4.69 | 14.53 ± 5.75 | 15.65 ± 5.61 | 15.65 ± 5.61 | 13.99 ± 4.91 | 0.913 |
| E | Zinc-α2-glycoprotein + Carbonic anhydrase VI | P25311 | 41.0/34.5 | 71 | 30 | 9.99 ± 2.20 | 9.66 ± 1.93 | 9.68 ± 2.11 a | 8.48 ± 1.93 b | 9.79 ± 2.13 a | 8.49 ± 2.05 b | 0.308 |
| P23280 | 41.0/35.5 | 150 | 39 | |||||||||
| H | Immunoglobulin kappa constant + Zymogen granule protein 16 homolog B | P01834 | 28.0/11.9 | 76 | 50 | 8.36 ± 2.15 a | 8.83 ± 2.25 a | 8.48 ± 2.55 a | 6.71 ± 2.05 b | 8.12 ± 2.56 a | 7.13 ± 2.57 a | 0.021 * |
| Q96DA0 | 28.0/22.7 | 74 | 37 | |||||||||
| I | Immunoglobulin kappa constant | P01834 | 23.5/11.9 | 74 | 50 | 7.41 ± 2.07 | 7.16 ± 2.20 | 7.19 ± 2.33 | 7.38 ± 1.95 | 7.56 ± 1.47 | 7.93 ± 1.30 | 0.616 |
| L | Prolactin-inducible protein | P12273 | 16.5/16.8 | 131 | 60 | 5.85 ± 1.68 | 6.32 ± 2.01 | 6.18 ± 1.47 | 6.06 ± 1.60 | 6.01 ± 1.85 | 6.41 ± 2.18 | 0.785 |
| M | Cystatin-SN | P01037 | 14.0/16.6 | 110 | 54 | 15.80 ± 4.55 a | 15.93 ± 4.14 a | 15.58 ± 3.87 a | 17.53 ± 4.20 b | 15.59 ± 3.12 a | 18.16 ± 5.41 b | 0.097 |
| Cystatin-S | P01036 | 14.0/16.5 | 109 | 58 | ||||||||
| Spot | Bread Odour | Bread Mastication | Rice Mastication | 1 Interaction Period * Treatment p-Value | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Before | After | p | Ratio (A/B) 1 | Before | After | p | Ratio (A/B) 1 | Before | After | p | Ratio (A/B) 1 | ||
| 4 | 0.404 ± 0.171 | 0.503 ± 0.177 | 0.382 | 1.243 | 0.508 ± 0.164 | 0.377 ± 0.986 | 0.02 * | 0.743 | 0.562 ± 0.136 | 0.357 ± 0.221 | 0.039 * | 0.635 | |
| 7 | 0.063 ± 0.054 | 0.071 ± 0.053 | 0.768 | 1.127 | 0.117 ± 0.057 | 0.093 ± 0.038 | 0.482 | 0.794 | 0.092 ± 0.050 | 0.159 ± 0.056 | 0.049 * | 1.735 | |
| 8 | 0.525 ± 0.349 | 0.551 ± 0.267 | 1.000 | 1.050 | 0.501 ± 0.152 | 0.680 ± 0.318 | 0.128 | 1.356 | 0.383 ± 0.171 | 0.633 ± 0.299 | 0.009 * | 1.652 | |
| 19 | 2.692 ± 1.319 | 4.763 ± 2.249 | 0.009 * | 1.769 | 2.326 ± 0.855 | 4.568 ± 2.357 | 0.03 * | 1.964 | 3.130 ± 1.417 | 3.721 ± 1.161 | 0.306 | 1.189 | |
| 22 | 2.862 ± 1.637 | 5.456 ± 3.245 | 0.009 * | 1.906 | 3.116 ± 1.681 | 5.319 ± 2.186 | 0.002 * | 1.707 | 3.476 ± 1.573 | 3.968 ± 1.764 | 0.493 | 7.066 | |
| 29 | 0.384 ± 0.128 | 0.372 ± 0.087 | 0.808 | 0.968 | 0.410 ± 0.063 | 0.721 ± 0.133 | 0.004 * | 1.757 | 0.549 ± 0.184 | 0.596 ± 0.233 | 0.634 | 1.087 | 0.03 * |
| 31 | 0.476 ± 0.202 | 0.549 ± 0.290 | 0.518 | 1.154 | 0.655 ± 0.205 | 0.706 ± 0.220 | 0.675 | 1.078 | 0.452 ± 0.180 | 0.653 ± 0.172 | 0.046 * | 1.443 | |
| 39 | 0.078 ± 0.048 | 0.772 ± 0.329 | 0.028 * | 11.501 | 0.648 ± 0.163 | 0.309 ± 0.130 | 0.018 * | 0.478 | 0.511 ± 0.343 | 0.364 ± 0.186 | 0.176 | 0.713 | <0.001 * |
| 42 | 0.101 ± 0.079 | 1.131 ± 0.900 | 0.022 * | 11.188 | 0.970 ± 0.190 | 0.594 ± 0.194 | 0.003 * | 0.613 | 0.817 ± 0.295 | 0.498 ± 0.104 | 0.054 | 0.610 | 0.002 * |
| 43 | 0.641 ± 0.334 | 1.143 ± 0.491 | 0.117 | 1.785 | 1.330 ± 0.332 | 0.839 ± 0.282 | 0.003 * | 0.631 | 1.200 ± 0.405 | 0.638 ± 0.233 | 0.013 * | 0.531 | 0.004 * |
| 45 | 0.561 ± 0.261 | 0.681 ± 0.338 | 0.444 | 1.214 | 0.783 ± 0.191 | 0.378 ± 0.163 | 0.0005 * | 0.482 | 0.544 ± 0.150 | 0.506 ± 0.252 | 0.723 | 0.930 | 0.006 * |
| 46 | 0.482 ± 0.311 | 0.338 ± 0.115 | 0.176 | 0.702 | 0.412 ± 0.220 | 0.225 ± 0.133 | 0.128 | 0.546 | 0.282 ± 0.066 | 0.125 ± 0.052 | 0.0005 * | 0.443 | |
| 48 | 0.127 ± 0.082 | 0.245 ± 0.186 | 0.269 | 1.937 | 0.504 ± 0.171 | 0.253 ± 0.155 | 0.039 * | 0.502 | 0.394 ± 0.218 | 0.119 ± 0.128 | 0.015* | 0.301 | 0.001 * |
| 49 | 0.317 ± 0.242 | 0.229 ± 0.177 | 0.542 | 0.723 | 0.494 ± 0.244 | 0.195 ± 0.209 | 0.009 * | 0.394 | 0.188 ± 0.131 | 0.186 ± 0.146 | 0.981 | 0.990 | |
| 52 | 0.131 ± 0.110 | 0.411 ± 0.314 | 0.029 * | 3.136 | 0.308 ± 0.147 | 0.274 ± 0.113 | 0.720 | 0.887 | 0.385 ± 0.287 | 0.098 ± 0.067 | 0.024 * | 0.254 | 0.001 * |
| 53 | 0.340 ± 0.189 | 0.656 ± 0.299 | 0.034 * | 1.928 | 0.683 ± 0.237 | 0.431 ± 0.165 | 0.012 * | 0.631 | 0.354 ± 0.090 | 0.400 ± 0.168 | 0.310 | 1.129 | |
| 54 | 0.513 ± 0.198 | 0.363 ± 0.240 | 0.045 * | 0.708 | 0.348 ± 0.096 | 0.265 ± 0.071 | 0.119 | 0.762 | 0.297 ± 0.074 | 0.152 ± 0.060 | 0.016 * | 0.512 | |
| 55 | 0.339 ± 0.119 | 0.548 ± 0.126 | 0.02 * | 1.600 | 0.640 ± 0.124 | 0.424 ± 0.109 | 0.001* | 0.663 | 0.436 ± 0.101 | 0.223 ± 0.144 | 0.002 * | 0.512 | 0.006 * |
| 56 | 0.546 ± 0.227 | 0.157 ± 0.151 | 0.011* | 0.287 | 0.328 ± 0.161 | 0.168 ± 0.110 | 0.070 | 0.512 | 0.154 ± 0.183 | 0.074 ± 0.096 | 0.412 | 0.480 | |
| 67 | 0.083 ± 0.052 | 0.202 ± 0.208 | 0.179 | 2.434 | 0.100 ± 0.094 | 0.209 ± 0.093 | 0.019 * | 2.102 | 0.169 ± 0.132 | 0.122 ± 0.096 | 0.238 | 0.723 | |
| 70 | 0.106 ± 0.078 | 0.220 ± 0.123 | 0.064 | 2.084 | 0.216 ± 0.169 | 0.170 ± 0.085 | 0.612 | 0.787 | 0.283 ± 0.215 | 0.122 ± 0.117 | 0.077 | 0.431 | 0.011 * |
| 75 | 0.069 ± 0.055 | 0.142 ± 0.059 | 0.028 * | 2.009 | 0.206 ± 0.132 | 0.204 ± 0.089 | 0.866 | 0.994 | 0.142 ± 0.043 | 0.077 ± 0.067 | 0.018 * | 0.544 | 0.012 * |
| 76 | 0.175 ± 0.062 | 0.113 ± 0.138 | 0.237 | 0.644 | 0.113 ± 0.036 | 0.080 ± 0.056 | 0.128 | 0.709 | 0.094 ± 0.035 | 0.058 ± 0.042 | 0.013 * | 0.613 | |
| 77 | 0.091 ± 0.051 | 0.397 ± 0.312 | 0.075 | 4.050 | 0.131 ± 0.086 | 0.284 ± 0.085 | 0.018 * | 2.175 | 0.129 ± 0.155 | 0.140 ± 0.147 | 0.753 | 1.195 | |
| 80 | 0.056 ± 0.055 | 0.404 ± 0.367 | 0.018 * | 7.178 | 0.168 ± 0.123 | 0.419 ± 0.152 | 0.018 * | 2.495 | 0.345 ± 0.213 | 0.268 ± 0.136 | 0.398 | 0.777 | 0.015 * |
| 95 | 0.490 ± 0.262 | 0.089 ± 0.037 | 0.028 * | 0.162 | 0.086 ± 0.104 | 0.221 ± 0.074 | 0.028 * | 2.579 | 0.236 ± 0.160 | 0.083 ± 0.059 | 0.068 | 0.352 | 0.019 * |
| 99 | 0.335 ± 0.243 | 0.065 ± 0.016 | 0.032 * | 0.194 | 0.094 ± 0.060 | 0.156 ± 0.057 | 0.398 | 1.600 | 0.091 ± 0.020 | 0.114 ± 0.056 | 0.735 | 1.512 | 0.012 * |
| 100 | 0.169 ± 0.160 | 0.140 ± 0.109 | 0.608 | 1.235 | 0.177 ± 0.084 | 0.257 ± 0.082 | 0.130 | 1.452 | 0.245 ± 0.083 | 0.143 ± 0.094 | 0.011 * | 0.582 | 0.003 * |
| 108 | 0.682 ± 0.293 | 0.166 ± 0.147 | 0.008 * | 0.243 | 0.107 ± 0.073 | 0.209 ± 0.084 | 0.013 * | 1.953 | 0.091 ± 0.043 | 0.098 ± 0.064 | 0.735 | 1.077 | 0.045 * |
| 116 | 0.155 ± 0.082 | 0.043 ± 0.030 | 0.037* | 0.277 | 0.028 ± 0.009 | 0.062 ± 0.017 | 0.004 * | 2.198 | 0.044 ± 0.028 | 0.074 ± 0.040 | 0.114 | 1.684 | 0.019 * |
| 120 | 0.096 ± 0.060 | 2.308 ± 1.602 | 0.019 * | 24.042 | 1.723 ± 0.945 | 2.462 ± 0.811 | 0.018* | 1.428 | 2.625 ± 1.676 | 1.793 ± 0.859 | 0.261 | 0.6830 | |
| 129 | 0.737 ± 0.707 | 1.732 ± 1.217 | 0.091 | 2.348 | 1.837 ± 0.373 | 0.979 ± 0.390 | 0.028 * | 0.533 | 0.698 ± 0.297 | 0.622 ± 0.209 | 0.866 | 0.892 | 0.005 * |
| 131 | 0.072 ± 0.058 | 0.871 ± 0.474 | 0.01 * | 8.736 | 1.335 ± 0.680 | 0.893 ± 0.235 | 0.083 | 0.669 | 1.207 ± 0.400 | 0.688 ± 0.322 | 0.017 * | 0.570 | 0.002 * |
| 132 | 1.143 ± 0.547 | 1.141 ± 0.676 | 0.994 | 0.998 | 1.392 ± 0.499 | 0.692 ± 0.172 | 0.004 * | 0.497 | 1.292 ± 0.879 | 0.888 ± 0.464 | 0.174 | 0.918 | |
| 133 | 0.820 ± 0.210 | 3.580 ± 0.977 | 0.028 * | 4.123 | 4.767 ± 1.911 | 0.959 ± 0.460 | 0.043 * | 1.510 | 3.819 ± 0.601 | 2.930 ± 1.162 | 0.063 | 0.794 | 0.01 * |
| 152 | 0.043 ± 0.112 | 0.503 ± 0.244 | 0.003 * | 11.698 | 0.779 ± 0.264 | 0.439 ± 0.169 | 0.024 * | 0.564 | 0.510 ± 0.197 | 0.296 ± 0.099 | 0.032 * | 0.580 | 0.002 * |
| 153 | 0.433 ± 0.288 | 0.654 ± 0.256 | 0.264 | 1.980 | 0.713 ± 0.189 | 0.478 ± 0.175 | 0.012 * | 0.671 | 0.623 ± 0.200 | 0.404 ± 0.236 | 0.062 | 0.649 | 0.011 * |
| 154 | 0.417 ± 0.041 | 0.589 ± 0.240 | 0.612 | 1.307 | 0.924 ± 0.492 | 0.484 ± 0.115 | 0.028 * | 0.524 | 0.693 ± 0.297 | 0.399 ± 0.229 | 0.018 * | 0.576 | |
| 155 | 0.450 ± 0.115 | 0.400 ± 0.297 | 0.678 | 0.889 | 0.690 ± 0.322 | 0.446 ± 0.876 | 0.047 * | 0.646 | 0.665 ± 0.158 | 0.430 ± 0.243 | 0.027 * | 0.647 | 0.005 * |
| 156 | 0.436 ± 0.132 | 0.411 ± 0.257 | 0.810 | 0.944 | 0.491 ± 0.251 | 0.320 ± 0.047 | 0.096 | 0.651 | 0.475 ± 0.140 | 0.304 ± 0.146 | 0.046 * | 0.641 | 0.023 * |
| 157 | 0.301 ± 0.093 | 0.345 ± 0.185 | 0.548 | 1.146 | 0.421 ± 0.196 | 0.266 ± 0.050 | 0.058 | 0.632 | 0.386 ± 0.119 | 0.273 ± 0.110 | 0.047 * | 0.709 | 0.047 * |
| Spot | Protein Identification | Apparent Isoelectric Point (pI) | Apparent Molecular Mass (kDa) | Ref (Identifying the Proteins by Mass Spectrometry) |
|---|---|---|---|---|
| 4 | Ig polymeric receptor | 5.7 | 100 | [20,21,22] |
| 7 | n.i. | 6.8 | 32 | |
| 8 | Prolactin inducible protein | 4.5 | 18 | [21] |
| 19 | Cystatin SA | 7.1 | 15 | [22] |
| 22 | Cystatin S | 4.0 | 15 | [21] |
| 29 | Prolactin inducible protein | 4.0 | 19 | [20,22] |
| 31 | 4.7 | 19 | [20,21,22] | |
| 39 | Ig Kappa chain reaction | 8.3 | 27 | [20,21,22] |
| 42 | 7.7 | 27 | ||
| 43 | 7.1 | 28 | ||
| 45 | 7.0 | 28 | ||
| 46 | 6.8 | 29 | ||
| 48 | 8.3 | 29 | ||
| 49 | 5.8 | 29 | ||
| 52 | 7.8 | 29 | ||
| 53 | 6.1 | 30 | ||
| 54 | 5.9 | 30 | ||
| 55 | 6.9 | 29 | ||
| 56 | n.i. | 5.7 | 29 | |
| 67 | n.i. | 5.2 | 31 | |
| 70 | n.i. | 8.3 | 31 | |
| 75 | n.i. | 7.3 | 33 | |
| 76 | SPLUNC | 5.4 | 33 | [20,22] |
| 77 | 5.3 | 33 | ||
| 80 | n.i. | 5.0 | 34 | |
| 95 | carbonic anhydrase VI | 6.9 | 42 | [22] |
| 99 | 5.9 | 45 | ||
| 100 | n.i. | 5.7 | 44 | |
| 108 | n.i. | 7.3 | 47 | |
| 116 | n.i. | 5.7 | 49 | |
| 120 | α-amylase | 7.0 | 60 | [19,21] |
| 129 | Ig alpha-1 chain C region | 5.8 | 66 | [15] |
| 131 | 5.7 | 66 | ||
| 132 | 5.6 | 68 | ||
| 133 | α-amylase | 6.3 | 68 | [19,21] |
| 152 | Ig polymeric receptor | 6.3 | 89 | [20,21,22] |
| 153 | 6.2 | 93 | ||
| 154 | 5.9 | 96 | ||
| 155 | 5.8 | 100 | ||
| 156 | 5.7 | 100 | ||
| 157 | 5.6 | 100 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carreira, L.; Midori Castelo, P.; Simões, C.; Capela e Silva, F.; Viegas, C.; Lamy, E. Changes in Salivary Proteome in Response to Bread Odour. Nutrients 2020, 12, 1002. https://doi.org/10.3390/nu12041002
Carreira L, Midori Castelo P, Simões C, Capela e Silva F, Viegas C, Lamy E. Changes in Salivary Proteome in Response to Bread Odour. Nutrients. 2020; 12(4):1002. https://doi.org/10.3390/nu12041002
Chicago/Turabian StyleCarreira, Laura, Paula Midori Castelo, Carla Simões, Fernando Capela e Silva, Cláudia Viegas, and Elsa Lamy. 2020. "Changes in Salivary Proteome in Response to Bread Odour" Nutrients 12, no. 4: 1002. https://doi.org/10.3390/nu12041002
APA StyleCarreira, L., Midori Castelo, P., Simões, C., Capela e Silva, F., Viegas, C., & Lamy, E. (2020). Changes in Salivary Proteome in Response to Bread Odour. Nutrients, 12(4), 1002. https://doi.org/10.3390/nu12041002