Nutrition, Health, and Disease: Role of Selected Marine and Vegetal Nutraceuticals





Abstract
1. Introduction
2. Nutraceuticals, Food Supplements, and Functional Foods
3. The Ebiosea Program: Marine and Vegetal Nutraceutical Research
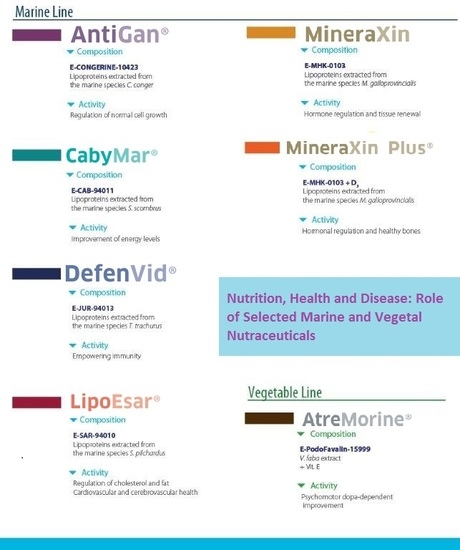
3.1. Marine Biotechnology-Related Products
3.1.1. LipoEsar (E-SAR-94010)
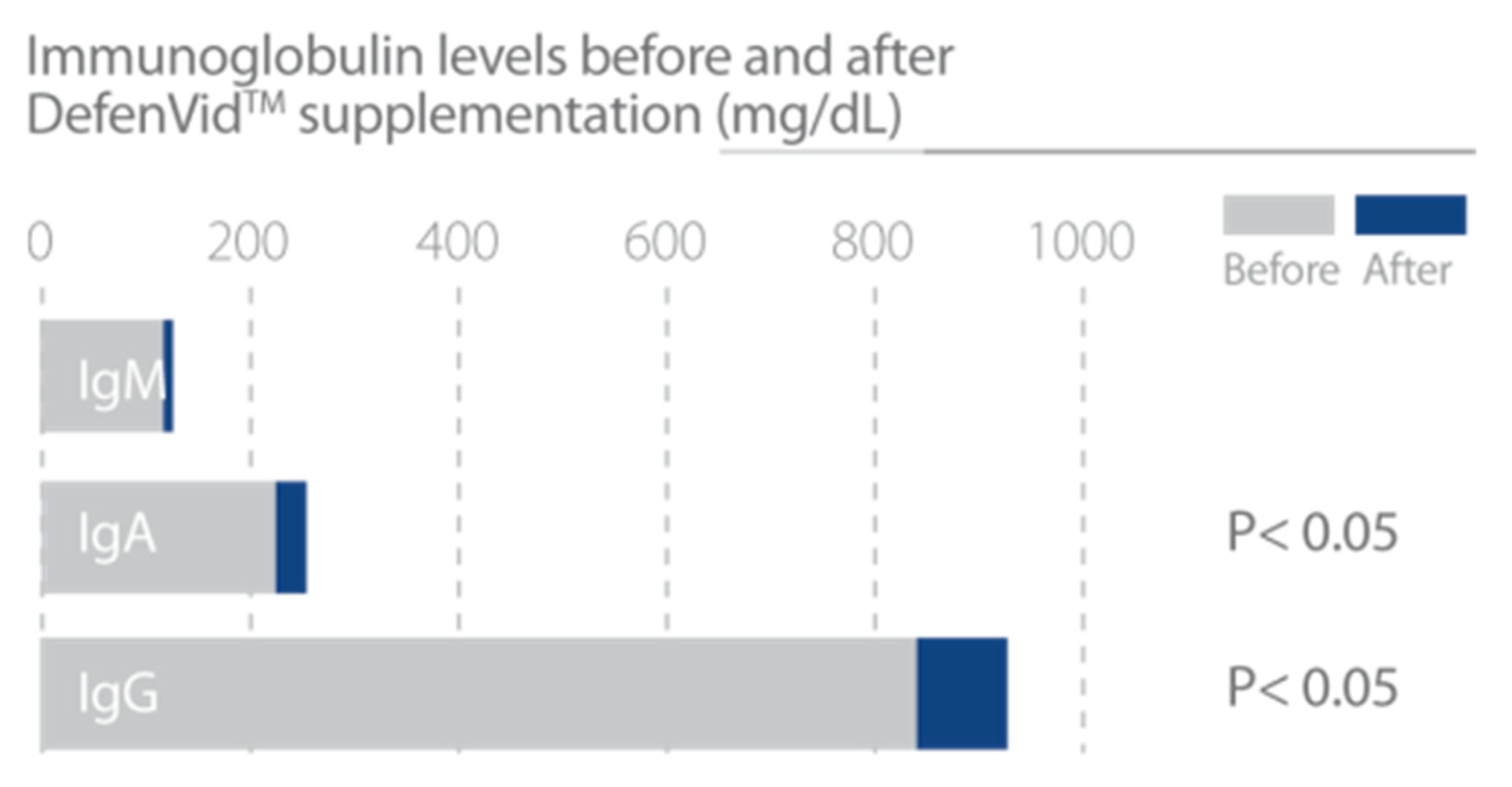
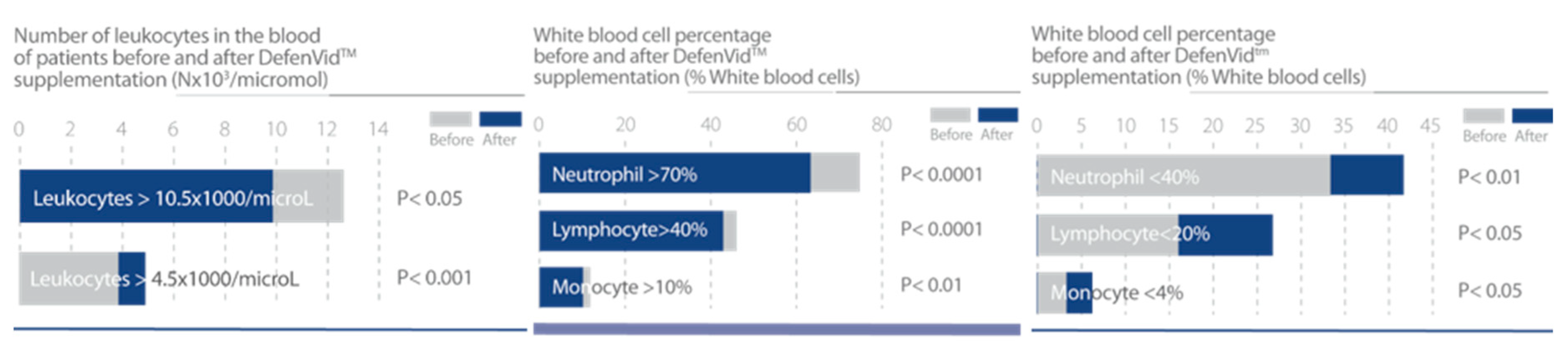
3.1.2. DefenVid (E-JUR-94013)
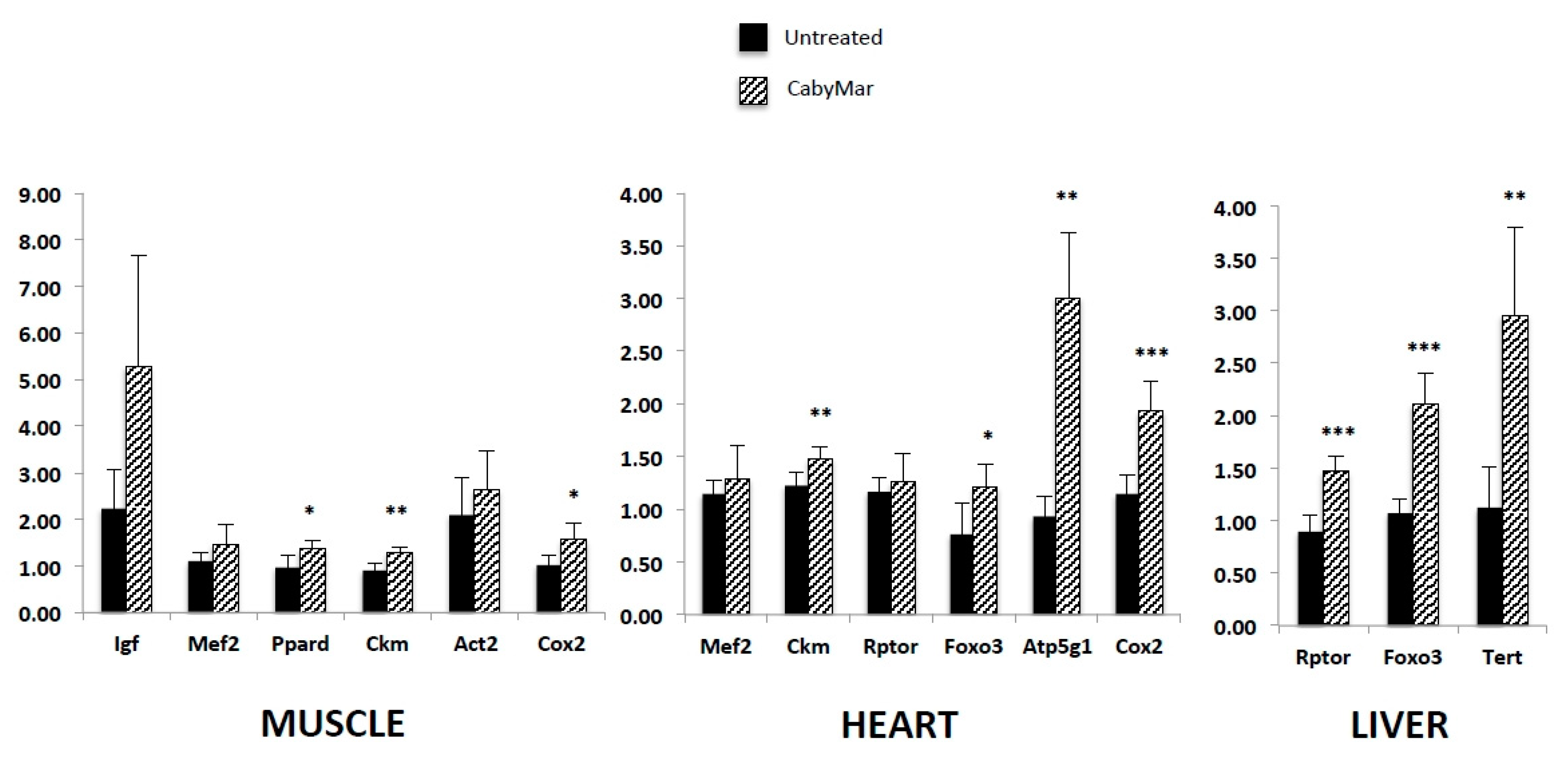
3.1.3. CabyMar (E-CAB-94011)
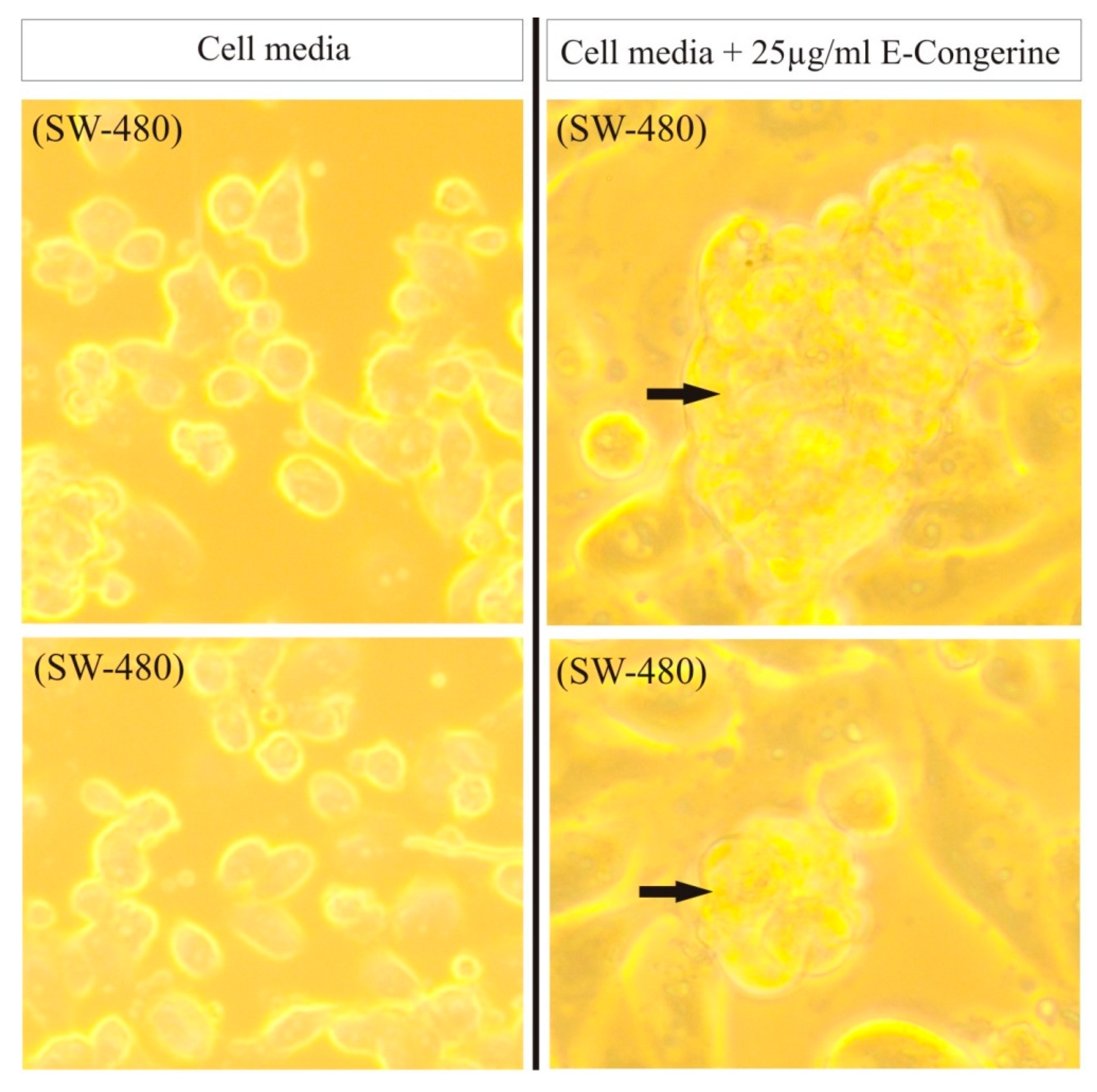
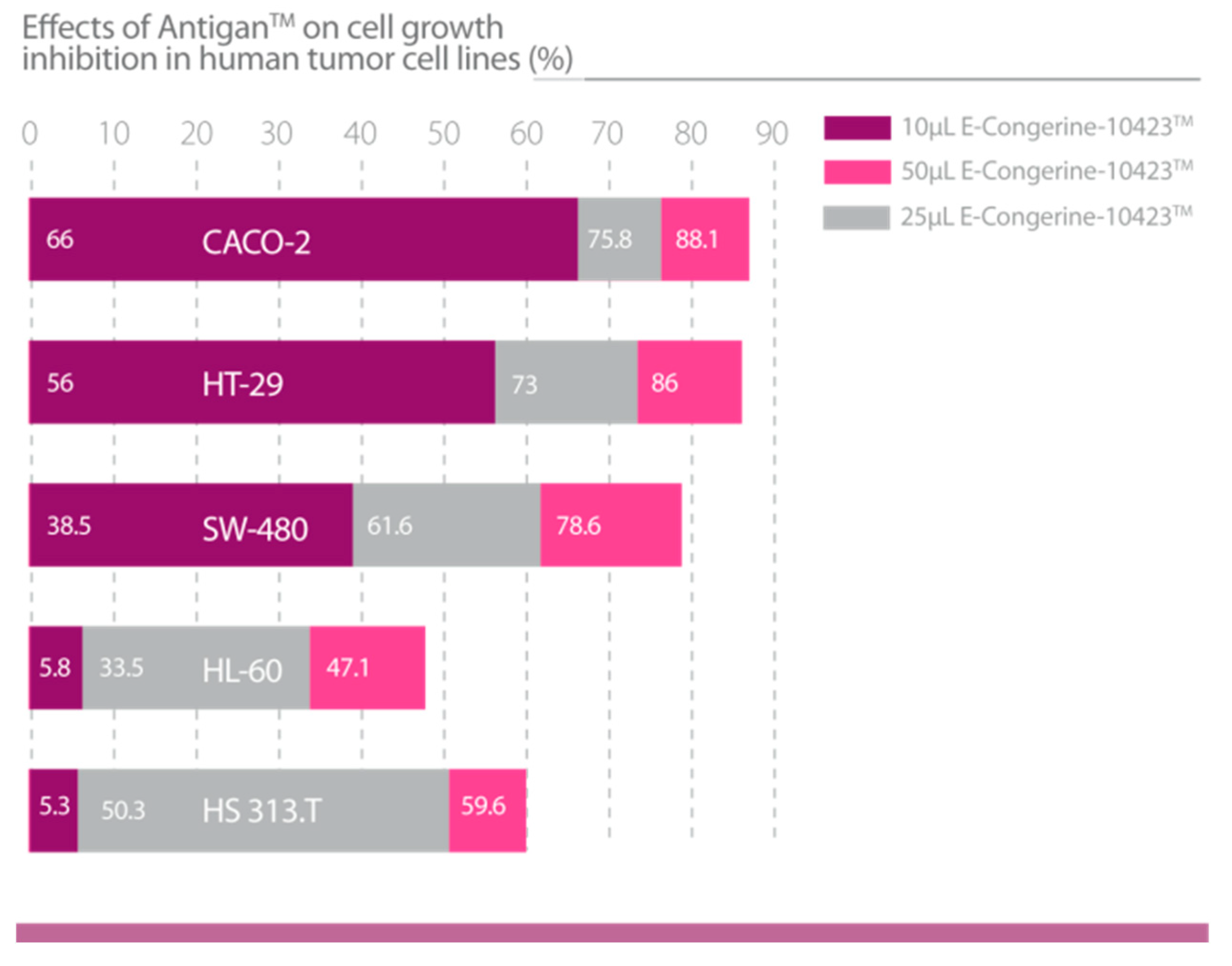
3.1.4. AntiGan (E-Congerine-10423)
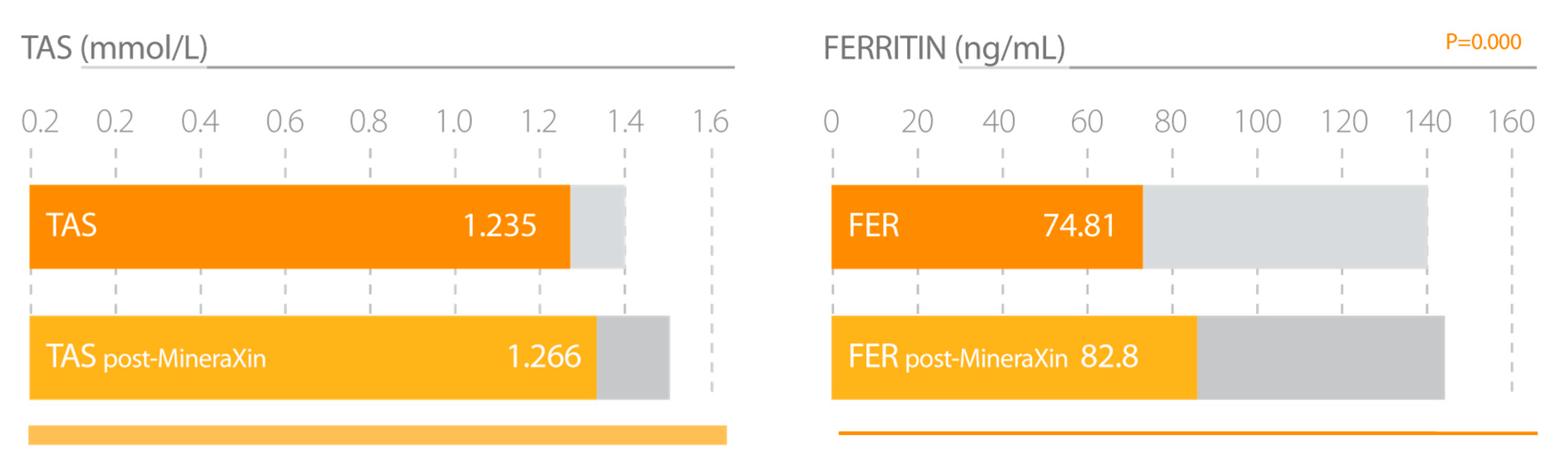
3.1.5. MineraXin (E-MHK-0103)
3.2. Plant Biotechnology-Related Products
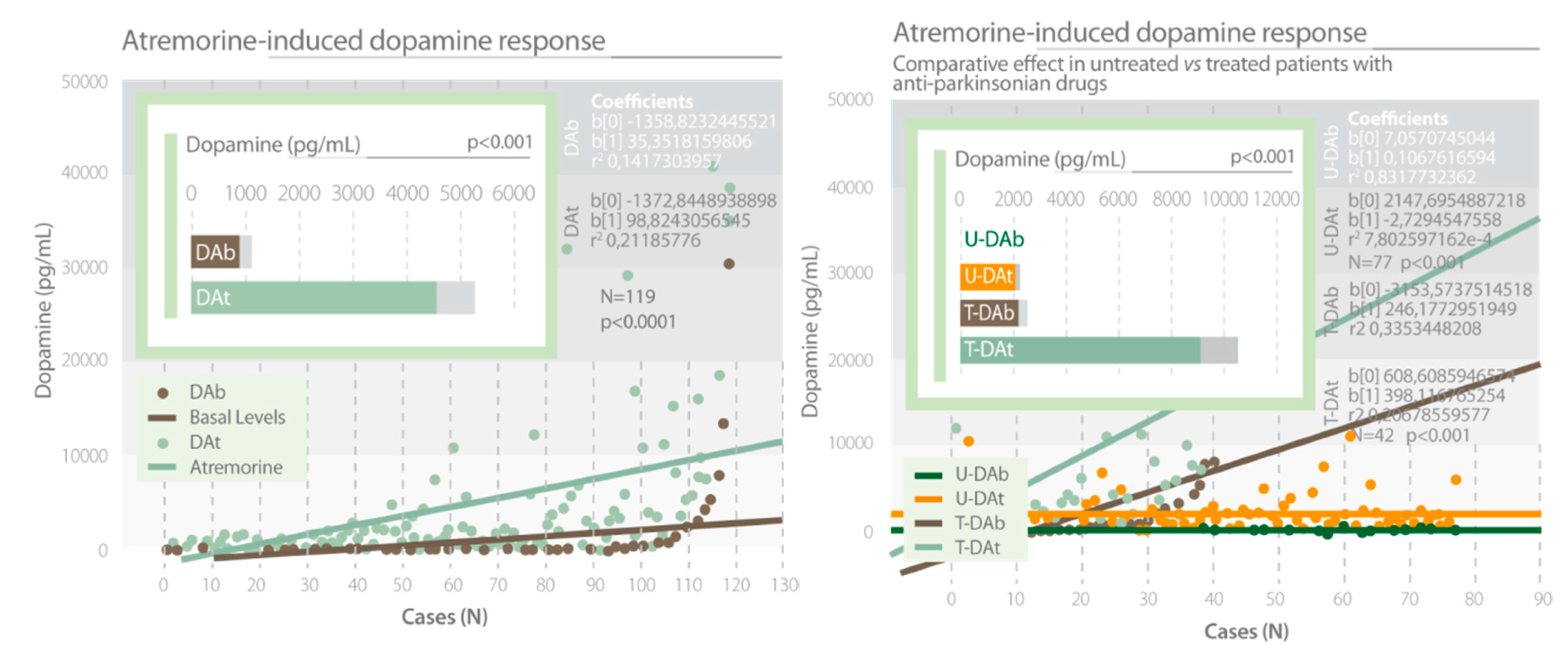
AtreMorine (E-PodoFavalin-15999)
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Callahan, D. The WHO definition of ‘health’. Stud. Hastings Cent. 1973, 1, 77–88. [Google Scholar] [CrossRef] [PubMed]
- WHO/FAO Expert Consultation. Diet, nutrition and the prevention of chronic diseases. World Health Organ. Tech. Rep. Ser. 2003, 916, 1–149. [Google Scholar]
- Ruel, G.; Shi, Z.; Zhen, S.; Zuo, H.; Kröger, E.; Sirois, C.; Lévesque, J.F.; Taylor, A.W. Association between nutrition and the evolution of multimorbidity: The importance of fruits and vegetables and whole grain products. Clin. Nutr. 2014, 33, 513–520. [Google Scholar] [CrossRef] [PubMed]
- Panico, S.; Mattiello, A.; Panico, C.; Chiodini, P. Mediterranean dietary pattern and chronic diseases. Cancer Treat. Res. 2014, 159, 69–81. [Google Scholar]
- Hesketh, J. Personalised nutrition: How far has nutrigenomics progressed? Eur. J. Clin. Nutr. 2013, 67, 430–435. [Google Scholar] [CrossRef]
- DeFelice, S.L. Nutrition Stymied: The Nutraceutical Solution. In Proceedings of the Plenary Lecture XXV National Congress of the Italian Chemical Society-SCI-The University of Calabria, Calabria, Italy, 9 September 2014; Available online: http://www.fimdefelice.org (accessed on 13 February 2019).
- H.R.3001–Nutraceutical Research and Education Act. Available online: https://www.congress.gov/bill/106th-congress/house-bill/3001/allactions? (accessed on 22 July 2019).
- Health Canada. Policy Paper—Nutraceuticals/Functional Foods and Health Claims on Foods. Available online: http://www.hc-sc.gc.ca/fn-an/label-etiquet/claims-reclam/nutra-funct_foods-nutrafonct_aliment-eng.php (accessed on 22 July 2019).
- Aronson, J.K. Defining ‘nutraceuticals’: Neither nutritious nor pharmaceutical. Br. J. Clin. Pharmacol. 2017, 83, 8–19. [Google Scholar] [CrossRef]
- Santini, A.; Tenore, G.C.; Novellino, E. Nutraceuticals: A paradigm of proactive medicine. Eur. J. Pharm. Sci. 2017, 96, 53–61. [Google Scholar] [CrossRef]
- Maneiro, E.; Lombardi, V.R.; Cacabelos, R. Rat cell cultures: Experimental models to study neurodegenerative disorders and new pharmacological compounds. Methods Find. Exp. Clin. Pharmacol. 1996, 18, 615–645. [Google Scholar]
- Lombardi, V.R.M.; Carrera, I.; Cacabelos, R. In vitro screening for cytotoxic activity of herbal extracts. Evid. Based Complement Altern. Med. 2017, 2017, 2675631. [Google Scholar] [CrossRef]
- Lombardi, V.R.M.; Cacabelos, R. E-SAR-94010: A marine fish extract obtained by advanced biotechnological methods. Drugs Future 1999, 24, 167–176. [Google Scholar] [CrossRef]
- Cacabelos, R. Parkinson’s disease: From pathogenesis to pharmacogenomics. Int. J. Mol. Sci. 2017, 18, 551. [Google Scholar] [CrossRef] [PubMed]
- Cacabelos, R.; Torrellas, C.; Carrera, I.; Cacabelos, P.; Corzo, L.; Fernández-Novoa, L.; Tellado, I.; Carril, J.C.; Aliev, G. Novel therapeutic strategies for dementia. CNS Neurol. Disord. Drug Targets 2016, 15, 141–241. [Google Scholar] [CrossRef] [PubMed]
- Cacabelos, R.; Lombardi, V.; Fernández-Novoa, L.; Carrera, I.; Cacabelos, P.; Corzo, L.; Carril, J.C.; Teijido, O. Basic and clinical studies with marine lipofishins and vegetal favalins in neurodegeneration and age-related disorders. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2018; Volume 59, pp. 195–225. [Google Scholar]
- Cacabelos, R. Application of Nutrigenomics to Alzheimer’s Disease. Agro Food Ind. hi-tech 2008, 19, 37. [Google Scholar]
- Lombardi, V.R.M.; Cagiao, A.; Fernández-Novoa, L.; Álvarez, X.A.; Corzo, M.D.; Zas, R.; Sampedro, C.; Cacabelos, R. Short-term food supplementation effects of a fish-derived extract on the immunological status of pregnant rats and their sucking pups. Nutr. Res. 2001, 21, 1425–1434. [Google Scholar] [CrossRef]
- Cacabelos, R.; Vallejo, A.I.; Lombardi, V.R.M.; Fernández-Novoa, L.; Pichel, V. E-SAR-94010®: A pleiotropic lipoprotein compound with powerful anti-atheromatous and lipid lowering effects. CNS Drug Rev. 2004, 10, 200–201. [Google Scholar] [CrossRef]
- Cacabelos, R. Pharmacogenomics, nutrigenomics and therapeutic optimization in Alzheimer’s disease. Aging Health 2005, 1, 303–348. [Google Scholar] [CrossRef]
- Cacabelos, R. Novel Biotechnological products from natural sources: Nutri/pharmacogenomic component. J. Nutr. Food Sci. 2016, 6, 6. [Google Scholar] [CrossRef]
- Cacabelos, R.; Carril, J.C.; Cacabelos, P.; Teijido, O.; Goldgaber, D. Pharmacogenomics of Alzheimer’s disease: Genetic determinants of phenotypic variation and therapeutic outcome. J. Gen. Med. Pharm. 2016, 1, 151–209. [Google Scholar]
- Lombardi, V.R.; Fernández-Novoa, L.; Etcheverría, I.; Seoane, S.; Cacabelos, R. Effects of fish-derived lipoprotein extracts on activation markers, Fas expression and apoptosis in peripheral blood lymphocytes. Int. Immunopharmacol. 2005, 5, 253–262. [Google Scholar] [CrossRef]
- Affane, F.; Louala, S.; El Imane Harrat, N.; Bensalah, F.; Chekkal, H.; Allaoui, A.; Lamri-Senhadji, M. Hypolipidemic, antioxidant and antiatherogenic property of sardine by-products proteins in high-fat diet induced obese rats. Life Sci. 2018, 199, 16–22. [Google Scholar] [CrossRef]
- Wang, D.D.; Hu, F.B. Dietary fat and risk of cardiovascular disease: Recent controversies and advances. Annu. Rev. Nutr. 2017, 37, 423–446. [Google Scholar] [CrossRef]
- Scaioli, E.; Liverani, E.; Belluzzi, A. The Imbalance between n-6/n-3 Polyunsaturated Fatty Acids and Inflammatory Bowel Disease: A Comprehensive Review and Future Therapeutic Perspectives. Int. J. Mol. Sci. 2017, 18, 2619. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.W.; Chien, Y.S.; Chen, Y.J.; Ajuwon, K.M.; Mersmann, H.M.; Ding, S.T. Role of n-3 polyunsaturated fatty acids in ameliorating the obesity-induced metabolic syndrome in animal models and humans. Int. J. Mol. Sci. 2016, 17, 1689. [Google Scholar] [CrossRef] [PubMed]
- Brown, A.L.; Zhu, X.; Rong, S.; Shewale, S.; Seo, J.; Boudyguina, E.; Gebre, A.K.; Alexander-Miller, M.A.; Parks, J.S. Omega-3 fatty acids ameliorate atherosclerosis by favorably altering monocyte subsets and limiting monocyte recruitment to aortic lesions. Arterioscler. Thromb. Vasc. Biol. 2012, 32, 2122–2130. [Google Scholar] [CrossRef] [PubMed]
- Jensen, I.J.; Mæhre, H.K. Preclinical and clinical studies on antioxidative, antihypertensive and cardioprotective effect of marine proteins and peptides—A review. Mar. Drugs 2016, 14, 211. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, V.R.; Fernández-Novoa, L.; Corzo, D.; Zas, R.; Cacabelos, R. Enhancement in immune function and growth using E-JUR-94013 supplementation. Methods Find. Exp. Clin. Pharmacol. 2002, 24, 573–578. [Google Scholar]
- Lombardi, V.R.M.; Corzo, L.; Carrera, I.; Cacabelos, R. The search for biomarine-derived compounds with immunomodulatory activity. J. Explor. Res. Pharmacol. 2018, 3, 30–41. [Google Scholar] [CrossRef]
- Cacabelos, R.; Carril, J.C.; Teijido, O. Pharmacogenomics and epigenomics of age-related neurodegenerative disorders: Strategies for drug development. In Antiaging Drugs: From Basic Research to Clinical Practice; Vaiserman, A.M., Ed.; Royal Society of Chemistry (RSC Drug Discovery Series): London, UK, 2017; Volume 57, pp. 75–141. [Google Scholar]
- Suzuki, D.; Furukawa, K.; Kimura, F.; Shimizu, H.; Yoshidome, H.; Ohtsuka, M.; Kato, A.; Yoshitomi, H.; Miyazaki, M. Effects of perioperative immunonutrition on cellmediated immunity, T helper type 1 (Th1)/Th2 differentiation, and Th17 response after pancreaticoduodenectomy. Surgery 2010, 148, 573–581. [Google Scholar] [CrossRef]
- Faber, J.; Berkhout, M.; Vos, A.P.; Sijben, J.W.; Calder, P.C.; Garssen, J.; van Helvoort, A. Supplementation with a fish oil-enriched, high-protein medical food leads to rapid incorporation of EPA into white blood cells and modulates immune responses within one week in healthy men and women. J. Nutr. 2011, 141, 964–970. [Google Scholar] [CrossRef]
- Totzeck, A.; Stettner, M.; Hagenacker, T. Early platelet and leukocyte decline in patients with neuroinflammatory disorders after intravenous immunoglobulins. Eur. J. Neurol. 2017, 24, 638–644. [Google Scholar] [CrossRef]
- Sassi, F.; Tamone, C.; D’Amelio, P. Vitamin D: Nutrient, Hormone, and Immunomodulator. Nutrients 2018, 10, 1656. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, V.R.; Fernández-Novoa, L.; Etcheverría, I.; Seoane, S.; Cacabelos, R. Studies on immunological, biochemical, hematological and growth regulation by Scomber scombrus fish protein extract supplementation in young pigs. Anim. Sci. J. 2005, 76, 159–170. [Google Scholar] [CrossRef]
- Rossignoli, C.P.; Dechandt, C.R.P.; Souza, A.O.; Sampaio, I.H.; Vicentini, T.M.; Teodoro, B.G.; Neto, M.P.C.; Ferrari, G.D.; Couto-Lima, C.A.; Alberici, L.C. Effects of intermittent dietary supplementation with conjugated linoleic acid and fish oil (EPA/DHA) on body metabolism and mitochondrial energetics in mice. J. Nutr. Biochem. 2018, 60, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Kawabata, F.; Mizushige, T.; Uozumi, K.; Hayamizu, K.; Han, L.; Tsuji, T.; Kishida, T. Fish protein intake induces fast-muscle hypertrophy and reduces liver lipids and serum glucose levels in rats. Biosci. Biotechnol. Biochem. 2015, 79, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Zhao, G.X.; Yang, X.R.; Wang, Y.M.; Zhao, Y.Q.; Chi, C.F.; Wang, B. Antioxidant Peptides from the Protein Hydrolysate of Spanish Mackerel (Scomberomorous niphonius) Muscle by in Vitro Gastrointestinal Digestion and Their In Vitro Activities. Mar. Drugs 2019, 17, 531. [Google Scholar] [CrossRef]
- Bradbury, K.E.; Tong, T.Y.N.; Key, T.J. Dietary Intake of High-Protein Foods and Other Major Foods in Meat-Eaters, Poultry-Eaters, Fish-Eaters, Vegetarians, and Vegans in UK Biobank. Nutrients 2017, 9, 1317. [Google Scholar] [CrossRef]
- Alexandrov, N.V.; Eelderink, C.; Singh-Povel, C.M.; Navis, G.J.; Bakker, S.J.L.; Corpeleijn, E. Dietary protein sources and muscle mass over the life course: The Lifelines Cohort Study. Nutrients 2018, 10, 1471. [Google Scholar] [CrossRef]
- Witard, O.C.; Combet, E.; Gray, S.R. Long-chain n-3 fatty acids as an essential link between musculoskeletal and cardio-metabolic health in older adults. Proc. Nutr. Soc. 2020, 79, 47–55. [Google Scholar] [CrossRef]
- Yubero-Serrano, E.M.; Delgado-Lista, J.; Tierney, A.C.; Perez-Martinez, P.; Garcia-Rios, A.; Alcala-Diaz, J.F.; Castaño, J.P.; Tinahones, F.J.; Drevon, C.A.; Defoort, C.; et al. Insulin resistance determines a differential response to changes in dietary fat modification on metabolic syndrome risk factors: The LIPGENE study. Am. J. Clin. Nutr. 2015, 102, 1509–1517. [Google Scholar] [CrossRef]
- Lombardi, V.R.M.; Carrera, I.; Cacabelos, R. In vitro and in vivo cytotoxic effect of AntiGan against tumor cells. Exp. Ther. Med. 2018, 15, 2547–2556. [Google Scholar] [CrossRef]
- Lombardi, V.R.M.; Carrera, I.; Corzo, L.; Cacabelos, R. Role of bioactive lipofishins in prevention of inflammation and colon cancer. Semin. Cancer Biol. 2019, 56, 175–184. [Google Scholar] [CrossRef] [PubMed]
- Lombardi, V.R.M.; Pereira, J.; Etcheverría, I.; Fernández-Novoa, L.; Seoane, S.; Cacabelos, R. Improvement of immune function by means of Conger conger extract in an in vivo rat model of cold stress. Food Agric. Immunol. 2006, 17, 115–127. [Google Scholar] [CrossRef]
- Lombardi, V.R.M.; Etcheverría, I.; Carrera, I.; Cacabelos, R.; Chacón, A. Prevention of chronic experimental colitis induced by dextran sulphate sodium (DSS) in mice treated with FR91. J. Biomed. Biotechnol. 2012, 2012, 826178. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kamiya, H.; Muramoto, K.; Goto, R. Purification and properties of agglutinins from conger eel, Conger Myriaster (Brevoort), skin mucus. Dev. Comp. Immunol. 1988, 12, 309–318. [Google Scholar] [CrossRef]
- Nakamura, O.; Matsuoka, H.; Ogawa, T.; Muramoto, K.; Kamiya, H.; Watanabe, T. Opsonic effect of congerin, a mucosal galectin of the Japanese conger, Conger myriaster (Brevoort). Fish. Shellfish Immunol. 2006, 20, 433–435. [Google Scholar] [CrossRef]
- Ogawa, T.; Watanabe, M.; Naganuma, T.; Muramoto, K. Diversified carbohydrate-binding lectins from marine resources. J. Amino Acids 2011, 2011, 838914. [Google Scholar] [CrossRef]
- Corzo, L.; Rodriguez, S.; Alejo, R.; Fernandez-Novoa, L.; Aliev, G.; Cacabelos, R. E-MHK-0103 (Mineraxin™): A novel nutraceutical with biological properties in menopausal conditions. Curr. Drug Metab. 2017, 18, 39–49. [Google Scholar] [CrossRef]
- Kariotoglou, D.M.; Mastronicolis, S.K. Phosphonolipids in the mussel Mytilus galloprovincialis. Z. Naturforsch. C 1998, 53, 888–896. [Google Scholar] [CrossRef]
- Burri, L.; Hoem, N.; Banni, S.; Berge, K. Marine omega-3 phospholipids: Metabolism and biological activities. Int. J. Mol. Sci. 2012, 13, 15401–15419. [Google Scholar] [CrossRef]
- Küllenberg, D.; Taylor, L.A.; Schneider, M.; Massing, U. Health effects of dietary phospholipids. Lipids Health Dis. 2012, 11, 3. [Google Scholar] [CrossRef]
- Grienke, U.; Silke, J.; Tasdemir, D. Bioactive compounds from marine mussels and their effects on human health. Food Chem. 2014, 142, 48–60. [Google Scholar] [CrossRef] [PubMed]
- Robledo, Y.; Madrid, J.F.; Leis, O.; Cajaraville, M.P. Analysis of the distribution of glycoconjugates in the digestive gland of the bivalve mollusc Mytilus galloprovincialis by conventional and lectin histochemistry. Cell Tissue Res. 1997, 288, 591–602. [Google Scholar] [CrossRef] [PubMed]
- Goggs, R.; Vaughan-Thomas, A.; Clegg, P.D.; Carter, S.D.; Innes, J.F.; Mobasheri, A.; Shakibaei, M.; Schwab, W.; Bondy, C.A. Nutraceutical therapies for degenerative joint diseases: A critical review. Crit. Rev. Food Sci. Nutr. 2005, 45, 145–164. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Chen, H.; Fan, F.; Shi, P.; Tu, M.; Cheng, S.; Wang, Z.; Du, M. Bone formation activity of an osteogenic dodecapeptide from blue mussels (Mytilus edulis). Food Funct. 2019, 10, 5616–5625. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Jia, Z.; Zhu, J.; Zou, Y.; Huang, G.; Hong, Y. Optimization of ultrasonic-assisted enzymatic extraction of polysaccharides from thick-shell mussel (Mytilus coruscus) and their antioxidant activities. Int. J. Biol. Macromol. 2019, 140, 1116–1125. [Google Scholar] [CrossRef] [PubMed]
- Romero, A.; Parada, E.; González-Lafuente, L.; Farré-Alins, V.; Ramos, E.; Cacabelos, R.; Egea, J. Neuroprotective effects of E-PodoFavalin-15999 (Atremorine®). CNS Neurosci. Ther. 2017, 23, 450–452. [Google Scholar] [CrossRef] [PubMed]
- Cacabelos, R.; Fernández-Novoa, L.; Alejo, L.; Corzo, L.; Rodríguez, S.; Alcaraz, M.; Nebril, L.; Cacabelos, P.; Fraile, C.; Carrera, I.; et al. E-PodoFavalin-15999 (Atremorine®)-induced neurotransmitter and hormonal response in Parkinson’s disease. J. Explor. Res. Pharmacol. 2016, 1, 1–12. [Google Scholar]
- Carrera, I.; Fernandez-Novoa, L.; Sampedro, C.; Cacabelos, R. Neuroprotective effect of Atremorine in an experimental model of Parkinson's disease. Curr. Pharm. Des. 2017, 23, 2673–2684. [Google Scholar] [CrossRef]
- Cacabelos, R.; Fernández-Novoa, L.; Alejo, R.; Corzo, L.; Alcaraz, M.; Nebril, L.; Cacabelos, P.; Fraile, C.; Carrera, I.; Carril, J.C. E-PodoFavalin-15999 (Atremorine®)-induced dopamine response in Parkinson’s disease: Pharmacogenetics-related effects. J. Genom. Med. Pharm. 2016, 1, 1–26. [Google Scholar]
- Cacabelos, R.; Carrera, I.; Alejo, R.; Fernández-Novoa, L.; Cacabelos, P.; Corzo, L.; Rodríguez, S.; Alcaraz, M.; Tellado, I.; Cacabelos, N.; et al. Pharmacogenetics of Atremorine-Induced Neuroprotection and Dopamine Response in Parkinson’s Disease. Planta Med. 2019, 85, 1351–1362. [Google Scholar]
- Kim, H.J.; Jeon, B.; Chung, S.J. Professional ethics in complementary and alternative medicines in management of Parkinson’s disease. J. Park. Dis. 2016, 6, 675–683. [Google Scholar] [CrossRef] [PubMed]
- Orlando, G.; Chiavaroli, A.; Leone, S.; Brunetti, L.; Politi, M.; Menghini, L.; Recinella, L.; Ferrante, C. Inhibitory Effects Induced by Vicia faba, Uncaria rhyncophylla, and Glycyrrhiza glabra Water Extracts on Oxidative Stress Biomarkers and Dopamine Turnover in HypoE22 Cells and Isolated Rat Striatum Challenged with 6-Hydroxydopamine. Antioxidants 2019, 8, 602. [Google Scholar] [CrossRef] [PubMed]
- Carange, J.; Longpré, F.; Daoust, B.; Martinoli, M.G. 24-Epibrassinolide, a Phytosterol from the Brassinosteroid Family, Protects Dopaminergic Cells against MPP-Induced Oxidative Stress and Apoptosis. J. Toxicol. 2011, 2011, 392859. [Google Scholar] [CrossRef] [PubMed]
- Hellenbrand, W.; Boeing, H.; Robra, B.P.; Seidler, A.; Vieregge, P.; Nischan, P.; Joerg, J.; Oertel, W.H.; Schneider, E.; Ulm, G. Diet and Parkinson’s disease. II: A possible role for the past intake of specific nutrients. Results from a self-administered food-frequency questionnaire in a case-control study. Neurology 1996, 47, 644–650. [Google Scholar] [CrossRef]
- Murakami, K.; Miyake, Y.; Sasaki, S.; Tanaka, K.; Fukushima, W.; Kiyohara, C.; Tsuboi, Y.; Yamada, T.; Oeda, T.; Miki, T.; et al. Disease Study Group. Dietary intake of folate, vitamin B6, vitamin B12 and riboflavin and risk of Parkinson's disease: A case-control study in Japan. Br. J. Nutr. 2010, 104, 757–764. [Google Scholar] [CrossRef]
- Rabin, M.L.; Stevens-Haas, C.; Havrilla, E.; Rosenstein, A.; Toffey, B.; Devi, T.; Earnhardt, M.C.; Kurlan, R. Complementary Therapies for Parkinson’s Disease: What’s Promoted, Rationale, Potential Risks and Benefits. Mov. Disord. Clin. Pract. 2015, 2, 205–212. [Google Scholar] [CrossRef]
- Kim, S.; Pallela, R. Medicinal Foods from marine animals: Current status and prospects. Adv. Food Nutr. Res. 2012, 65, 1–9. [Google Scholar]
- Calder, P.C. n-3 fatty acids and cardiovascular disease: Evidence explained and mechanisms explored. Clin. Sci. 2004, 107, 1–11. [Google Scholar] [CrossRef]
- Rudkowska, I.; Marcotte, B.; Pilon, G.; Lavigne, C.; Marette, A.; Vohl, M.C. Fish nutrients decrease expression levels of tumor necrosis factor-alpha in cultured human macrophages. Physiol. Genom. 2010, 40, 189–194. [Google Scholar] [CrossRef]
- Lund, E.K. Health benefits of seafood; Is it just the fatty acids? Food Chem. 2013, 140, 413–420. [Google Scholar] [CrossRef]
- Khalili Tilami, S.; Sampels, S. Nutritional value of fish: Lipids, proteins, vitamins, and minerals. Rev. Fish. Sci. Aquac. 2017, 26, 1–11. [Google Scholar] [CrossRef]
- Tacon, A.G.J.; Metian, M. Fish matters: Importance of aquatic foods in human nutrition and global food supply. Rev. Fish. Sci. 2013, 21, 22–38. [Google Scholar] [CrossRef]
- Health Organization & Food and Agriculture Organization of the United Nations. Report of the Joint FAO/WHO Expert Consultation on the Risks and Benefits of Fish Consumption, 25–29 January 2010, Rome, Italy; World Health Organization: Geneva, Switzerland, 2011. Available online: https://apps.who.int/iris/handle/10665/44666 (accessed on 17 October 2019).
- Wijesekara, I.; Kim, S.K. Angiotensin-i-converting enzyme (ACE) inhibitors from marine resources: Prospects in the pharmaceutical industry. Mar. Drugs 2010, 8, 1080–1093. [Google Scholar] [CrossRef] [PubMed]
- Wakimoto, T.; Kondo, H.; Nii, H.; Kimura, K.; Egami, Y.; Oka, Y.; Yoshida, M.; Kida, E.; Ye, Y.; Akahoshi, S.; et al. Furan fatty acid as an anti-inflammatory component from the green-lipped mussel Perna canaliculus. Proc. Natl. Acad. Sci. USA 2011, 108, 17533–17537. [Google Scholar] [CrossRef]
- Emelyanov, A.; Fedoseev, G.; Krasnoschekova, O.; Abulimity, A.; Trendeleva, T.; Barnes, P.J. Treatment of asthma with lipid extract of New Zealand green-lipped mussel: A randomised clinical trial. Eur. Respir. J. 2002, 20, 596–600. [Google Scholar] [CrossRef]
- Badiu, D.L.; Balu, A.M.; Barbes, L.; Luque, R.; Nita, R.; Radu, M.; Tanase, E.; Rosoiu, N. Physico-chemical characterisation of lipids from Mytilus galloprovincialis (L.) and Rapana venosa and their healing properties on skin burns. Lipids 2008, 43, 829–841. [Google Scholar] [CrossRef]
- Badiu, D.L.; Luque, R.; Dumitrescu, E.; Craciun, A.; Dinca, D. Amino acids from Mytilus galloprovincialis (L.) and Rapana venosa molluscs accelerate skin wounds healing via enhancement of dermal and epidermal neoformation. Protein J. 2010, 29, 81–92. [Google Scholar] [CrossRef]
- Rajapakse, N.; Jung, W.K.; Mendis, E.; Moon, S.H.; Kim, S.K. A novel anticoagulant purified from fish protein hydrolysate inhibits factor XIIa and platelet aggregation. Life Sci. 2005, 76, 2607–2619. [Google Scholar] [CrossRef]
- Norling, L.V.; Ly, L.; Dalli, J. Resolving inflammation by using nutrition therapy: Roles for specialized proresolving mediators. Curr. Opin. Clin. Nutr. Metab. Care 2017, 20, 145–152. [Google Scholar] [CrossRef]
- Harris, K.A.; Hill, A.M.; Kris-Etherton, P.M. Health benefits of marine derived omega-3 fatty acids. ACSMS Health Fit. J. 2010, 14, 22–28. [Google Scholar] [CrossRef]
- Sijtsma, L.; de Swaaf, M.E. Biotechnological production and applications of the omega-3 polyunsaturated fatty acid docosahexaenoic acid. Appl. Microbiol. Biotechnol. 2004, 64, 146–153. [Google Scholar] [CrossRef] [PubMed]
- Ramaa, C.S.; Shirode, A.R.; Mundada, A.S.; Kadam, V.J. Nutraceuticals-an emerging era in the treatment and prevention of cardiovascular diseases. Curr. Pharm. Biotechnol. 2006, 7, 15–23. [Google Scholar] [CrossRef] [PubMed]
- Mordor Intelligence. North America Nutraceutical Market—Growth, Trends and Forecasts (2018–2022). Available online: www.mordorintelligence.com (accessed on 25 July 2019).
- Europe Nutraceutical Market-Growth, Trends, and Forecast (2019–2024). Available online: www.researchandmarkets.com (accessed on 25 July 2019).
- Sapienza, C.; Issa, J.P. Diet, nutrition, and cancer epigenetics. Annu. Rev. Nutr. 2016, 36, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Mathers, J.C. Nutrigenomics in the modern era. Proc. Nutr. Soc. 2017, 76, 265–275. [Google Scholar] [CrossRef]
- Murgia, C.; Adamski, M.M. Translation of nutritional genomics into nutrition practice: The next step. Nutrients 2017, 9, 366. [Google Scholar] [CrossRef]
- Ramos-Lopez, O.; Milagro, F.I.; Allayee, H.; Chmurzynska, A.; Choi, M.S.; Curi, R.; De Caterina, R.; Ferguson, L.R.; Goni, L.; Kang, J.X.; et al. Guide for current nutrigenetic, nutrigenomic, and nutriepigenetic approaches for precision nutrition involving the prevention and management of chronic diseases associated with obesity. J. Nutr. Nutr. 2017, 10, 43–62. [Google Scholar] [CrossRef]
- Pérez-Jiménez, F.; Pascual, V.; Meco, J.F.; Pérez Martínez, P.; Delgado Lista, J.; Domenech, M.; Estruch, R.; León-Acuña, A.; López-Miranda, J.; Sánchez-Ramos, A.; et al. Document of recommendations of the SEA 2018. Lifestyle in cardiovascular prevention. Clin. Investig. Arterioscler. 2018, 30, 280–310. [Google Scholar]
- Subbiah, M.T. Nutrigenetics and nutraceuticals: The next wave riding on personalized medicine. Transl. Res. 2007, 149, 55–61. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| MARINE LINE | VEGETABLE LINE | |||||
|---|---|---|---|---|---|---|
| NUTRACEUTICALS | LipoEsar (E-SAR-94010) | DefenVid (E-JUR-94013) | CabyMar (E-CAB-94011) | AntiGan (E-Congerine-10423) | MineraXin (E-MHK-0103) | Atremorine (E-PodoFavalin-15999) |
| Origin | Sardina pilchardus (Dorsal muscle) | Trachurus trachurus (Dorsal muscle) | Scomber scombrus (Dorsal muscle) | Conger conger (Skin and muscular structures) | Mytillus galloprovincialis | Vicia faba L. (Structural components) |
| Bioactive properties | Atheroprotector Immune-enhancing properties | Immunorregulator Anti-inflammatory | Bioenergizing Antioxidant Anti-inflammatory | Apoptotic Carcinogenic | Antiosteoporotic Antioxidant Hormonal regulator | Pro-dopaminergic Antioxidant Neuroprotector |
| Nutritional composition | Protein 65% Unsaturated FA (% of total) 11% EPA, DHA, Oleic Saturated FA (% of total) 9% Vitamins C, D, B complex, inositol Minerals Potassium Phosphorous Calcium Sodium Magnesium Zinc Iron Manganese Copper | Protein 70% Unsaturated FA (% of total) 12% EPA, DHA, alpha-Linolenic, Oleic Saturated FA (% of total) 7% Vitamins A, C, D, B complex, inositol Minerals Calcium Potassium Phosphorus Magnesium Iron Iodine Zinc Copper | Protein 65% Unsaturated FA (% of total) 18% Monounsaturated FA (% of fat) LCMUFA 58% cis-gondoic acid cis-cetoleic acid Polyunsaturated FA (% of fat) 21% EPA, DHA, Linoleic Saturated FA (% of total) 6% Vitamins B complex (B5, B6, B12), C, D, E Minerals Phosphorus Sodium Magnesium Calcium Copper Iron Potassium Manganese Zinc | Protein 80% Unsaturated FA (% of fat) 70% Monounsaturated FA Oleic, palmitoleic, gadoleic Polyunsaturated FA DHA, Linoleic EPA Saturated FA (% of fat) 30% Vitamins A, D, B complex (B1, B2, B3) Minerals Phosphorus Calcium Potassium Magnesium Iron Zinc | Protein 65% Unsaturated FA (% of fat) 69% mostly EPA Polyunsaturated FA (% of fat) 45.4% Monounsaturated FA (% of fat) 23.6% Saturated FA (% of fat) 31% Phospholipid fraction 61.5% choline, ethanolamine phosphoglycerides and ceramide aminoethylphosphonates Vitamins B complex (B2, B5, B9, B12) Minerals Phosphorus Calcium Potassium Sodium Magnesium Iron Zinc Manganese Selenium Copper | Protein 65% L-Dopa 20 mg/gr extract Carbohydrates 66% Dietary fiber 30% Monounsaturated FA 29% (% of fat) Polyunsaturated FA 41.1% (% of fat) Saturated FA (% of fat) 29.8% Pigments Carotenoids Lutein, β-Carotenes, Epoxides, t-Zeaxanthin Phytosterols β-sitosterol, Campesterol Stigmasterol, Sitostanol, Cholesterol Vitamins B complex (B2, B3, B5, B6, B9), C, E, K Minerals Manganese Potassium Calcium Sodium Magnesium Iron Zinc Copper Selenium |
| LipoEsar | ||
|---|---|---|
| Study | Characteristics | Results–Conclusions |
| Lombardi, V.R.M. et al., 1999 [13]. | Sixty male Sprague-Dawley rats. Three groups of rats. One with normal diet, and two groups supplemented with E-SAR-94010, 1 mg and 5 mg. Experimental time 12 weeks. Biochemical study. Experimental group 10 healthy volunteers. Two sequential doses of E-SAR-94010 during two weeks each (1500 mg/day and 500 mg/day in soft gelatinous capsules). Biochemical and immunological analyses at baseline and at weeks 1 and 2. Participants continued their usual diet. | 1. No changes in rats body weight. 2. Significant changes in cholesterol and triglycerides levels comparing to rats control group at weeks 8 and 12. 3. In healthy volunteers, significant differences in HDL and LDL levels were observed at the end of the study between the group of 500 mg and 1500 mg. 4. The lymphocyte markers showed a significant increase in CD3, CD4 y CD25 antigen expression in both treatment groups. |
| Lombardi, V.R.M. et al., 2001 [18]. | Fourteen Sprague-Dawley rats. Two groups of 7 rats. Group 1 with standard diet. Group 2 supplemented with 50 mg/day of E-SAR-94010. Experimental time 12 weeks (6 weeks before pregnancy and 6 weeks after pregnancy). Intragastric treatment administration. Biochemical and immunological studies. | 1. E-SAR-94010 significantly reduced triglycerides levels in the pups and their mothers of the supplemented group with respect to control group. 2. No significant differences in the atherogenic index between both experimental groups. 3. E-SAR-94010 increased levels of CD25, CD28, CD54 and CD56 on lymphocytes from the supplemented group of rats and their pups. |
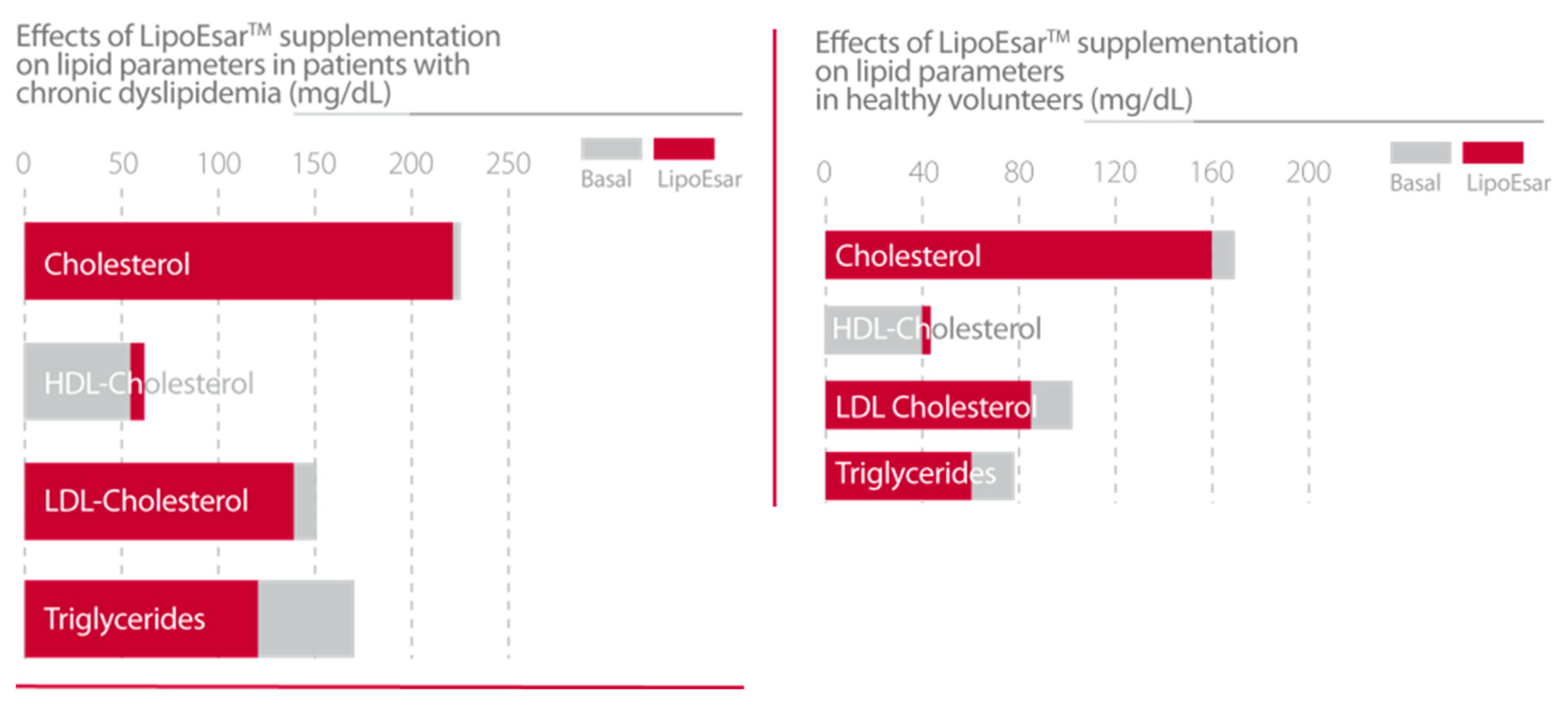
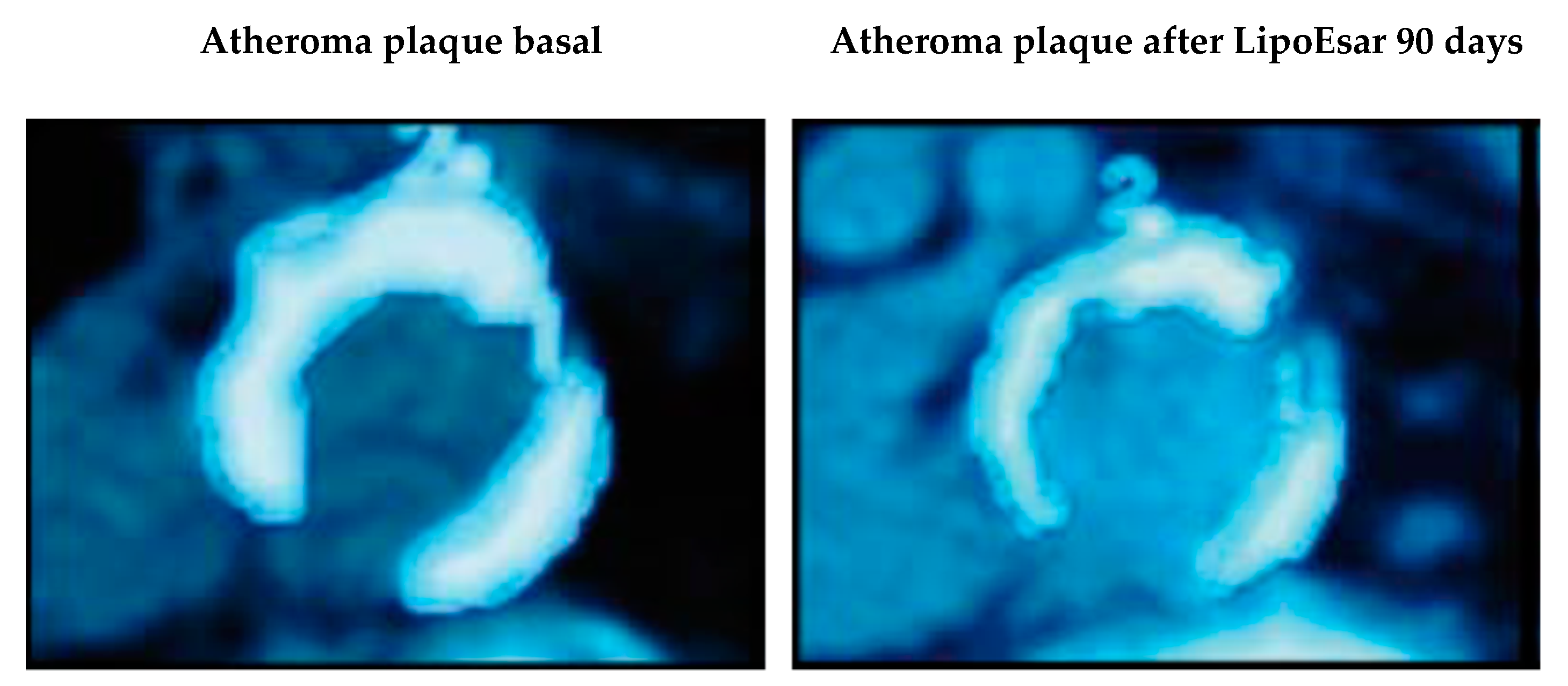
| Cacabelos, R. et al., 2004 [19]. | Description of the principal clinical effects of LipoEsar. Duration of treatment 1–3 months Daily dose of 250–500 mg. APOE genotype dependent effects of LipoEsar in 419 dyslipemic patients. | 1. LipoEsar reduced blood total cholesterol, glucose, uric acid, triglycerides, ALT and AST. 2. LipoEsar diminished the size of xanthelasma plaques after 6–9 months of treatment. 3. The therapeutic response of patients with dyslipemia to LipoEsar is APOE-related. |
| DefenVid | ||
|---|---|---|
| Study | Characteristics | Results–Conclusions |
| Lombardi, V.R. et al., 2002 [30]. | Experimental group 300 pigs. Fifteen groups of 20 pigs each. Five dietary treatments with different amount of fish extracts, sardine, mackerel and E-JUR-94013 (horse mackerel), in their habitual diet. Duration of the study 42 days. Anthropometric, biochemical and immunological analysis. | 1. E-JUR-94013 regulated IgA synthesis or release. 2. E-JUR-94013 reduced cholesterol levels in the sera of treated pigs. 3. E-JUR-94013 produced significant differences in feed intake and feed:gain ratio with respect to control group. 4. E-JUR-94013 increased the levels of IgA, IgM and IgE with respect to control group. |
| Lombardi, V.R. et al., 2005 [23]. | Experimental group 24 healthy volunteers. Evaluation of the effect of E-JUR-94013 on in vitro peripheral blood lymphocytes activation, FAS expression and apoptosis. | 1. E-JUR significantly increased the levels of CD25, CD8, CD38, CD19 and HLA-DR antigen expression. 2. E-JUR significantly reduced the percentages of apoptotic CD19 CD38 double positive lymphocytes. |
| Lombardi, V.R.M et al., 2018 [31]. | Three clinical studies to evaluate the effect of E-JUR-94013 in the improvement of immune system function. Study #1 50 patients. Study #2 205 patients. Study #3 1500 patients. Treatment regimen 750 mg/day of E-JUR-94013. Duration of treatment: Study #1 6 months; Study #2 and #3 3 months. In Study#2 patients were divided in 2 subgroups according to the number of leukocytes, high or low relative to reference ranges. In Study #3 a genetic analysis is incorporated to investigate the effect of genes in the response to DefenVid. | 1. Study #1 showed an increase in all leukocyte subclasses with a significant increase in the number of neutrophils and eosinophils. Serum immunoglobulins A, G and M were increased. A slight reduction in the concentration of IgE was found. 2. Study #2 showed an immunomodulatory effect. with an increase in white cells in the immunodeficient group and a decrease in white cells in the group with high leukocyte count at baseline, bringing the white cell count to normal ranges in both cases. 3. In Study #3 DefenVid was affected by polymorphic variations of genes involved in immune regulation of inflammation. |
| CabyMar | ||
|---|---|---|
| Study | Characteristics | Results - Conclusions |
| Lombardi, V.R. et al., 2005 [23]. | Experimental group 24 healthy volunteers. Evaluation of the effect of E-CAB-94011 on in vitro peripheral blood lymphocytes (PBL) activation, FAS expression and apoptosis. | 1. E-CAB significantly increased the levels of CD25, CD8, CD38, CD19 and HLA-DR antigen expression. 2. E-CAB significantly reduced the percentages of apoptotic CD19 CD38 double positive lymphocytes. |
| Lombardi, V.R. et al., 2005 [37]. | Experimental group 360 piglets divided in 4 groups (Group 1 normal diet, Group 2 supplemented diet with 0,1 g/Kg E-CAB, Group 3 supplemented with 0,25 g/Kg E-CAB, and Group 4 supplemented with 0,5 g/Kg). Anthropometric, biochemical and immunological analysis. Duration of the study 56 days. | 1. E-CAB-94011 increased total body weight. 2. E-CAB-94011 induced significant changes in total cholesterol, LDL, and hepatic transaminases. 3. E-CAB-94011 increases A and G immunoglobulin levels. 4. E-CAB-94011 increased CD4, CD8, CD25 and CD56 lymphocyte antigen expression. |
| Cacabelos, R. et al., 2018 [16]. | Mice treated with CabyMar. Biochemical, histological, and gene expression analysis. | 1. CabyMar significantly increased urea and creatinine levels and decreased glucose and lactate levels. 2. CabyMar significantly increased gene expression biomarkers in muscle and heart tissue. |
| AntiGan | ||
|---|---|---|
| Study | Characteristics | Results - Conclusions |
| Lombardi, V.R.M. et al., 2006 [47]. | Six Sprague-Dawley rats. Acute experimental model of immune system stress induced by exposure of rats to a hypothermic shock. Whole blood from rats was studied under two different concentrations of AntiGan (20 and 100 µg/mL) during 4, 16 and 24 hours. Analysis of T lymphocytes markers, and phagocytic activity. | 1. Increased CD4 and CD28 antigen expression in both groups of treatment. 2. Increased percentage of CD4 CD25 double positive T lymphocytes. 3. Increased phagocytic activity (monocytes and granulocytes) in both groups of treatment. |
| Lombardi, V.R.M. et al., 2018 [45]. | Evaluation of the apoptogenic activity of AntiGan after 24 h of incubation (10, 25 and 50 μL/mL AntiGan) of HL-60, Hs 313.T, SW-480, Caco-2 and HT-29 cell lines using growth inhibition and apoptosis activity assays. In vivo studies in mice (n = 56) by inducing colitis with oral administration of 2% dextran sulphate sodium (DSS) for 6 weeks. AntiGan was administered integrated in the diet as pellet biscuits (2,5%, 5%, 10% AntiGan). | 1. AntiGan treatment inhibited growth in tumor cell lines. 2. AntiGan induced apoptosis in tumor cell lines. 3. AntiGan downregulated Bcl-2 gene expression. 4. AntiGan displayed a powerful anti-inflammatory effect in DSS-induced colitis. |
| Lombardi, V.R.M. et al., 2019 [46]. | The effect of E-Congerine-10423 on tumor markers was studied in healthy subjects (n = 50) and in patients with different types of cancer (n = 156) at the time of diagnosis. AntiGan treatment 750 mg/day. | 1. AntiGan produced, in about 50% of the patients, a reduction in the levels of tumor markers, especially in patients with cancer. |
| MineraXin | ||
|---|---|---|
| Study | Characteristics | Results–Conclusions |
| Corzo, L. et al., 2017 [52]. | Experimental group 91 perimenopausal and postmenopausal women. MineraXin treatment left 750 mg/day during 3 months. Analysis of clinical symptoms, hormonal status, hypothalamic-pituitary-bone axis, markers of bone formation and resorption, antioxidant status, iron stores, cortisol, and BMI. | 1. Significant improvement in general status, hot flash frequency, emotional stability and musculoskeletal pain. 2. Increased serum estradiol and inhibin-A concentrations. 3. Decreased FSH and LH levels. 4. Increased serum GH and IGF-1 levels. 5. Slight increase in bone formation and a moderate decrease in bone resorption. 6. Significant increase of antioxidant status. 7. Increased ferritin concentrations. |
| AtreMorine | ||
|---|---|---|
| Study | Characteristics | Results–Conclusions |
| Cacabelos, R. et al., 2016 [62]. | Experimental group 119 patients with parkinsonian disorders. Two groups of patients, untreated vs. chronically treated patients. Oral dose of AtreMorine 5 g. Analysis of dopamine levels. APOE and CYP genetic studies. | 1. A single oral dose of AtreMorine (5g) induced a significant increase in dopamine levels after 1 hour. 2. The AtreMorine-induced dopamine response was different in APOE and CYP genetic variants. |
| Cacabelos, R. et al., 2016 [63]. | Experimental group, 119 patients with parkinsonian disorders. Two groups of patients, untreated vs. chronically treated patients. Oral dose of AtreMorine, 5 g. Analysis of neurotransmitters and hormones levels. | 1. A single oral dose of AtreMorine (5g) induced a significant increase in dopamine, noradrenalin and adrenaline levels after 1 hour. 2. AtreMorine induced a significant decrease in the levels of prolactin, GH, and cortisol. |
| Carrera, I. et al., 2017 [64]. | Experimental group, wild type C57BL6/J mice. MPTP-induced mice model of Parkinson Disease. Duration protocol 7 weeks. AtreMorine at doses of 2 mg and 4 mg was administered integrated in pellet biscuits. Immunohistochemical analysis. Motor function analysis. | 1. AtreMorine protected against MPTP-induced dopaminergic Neurodegeneration. 2. AtreMorine inhibited microglial activation and cell death in the Substantia Nigra. 3. AtreMorine improved motor and cognitive functions in MPTP mice model. |
| Cacabelos, R. et al., 2019 [65]. | Experimental group, 183 patients with parkinsonian disorders. Oral dose of AtreMorine, 5 g. Study of the influence of genes involved in pathogenic, metabolic, transporter, pleiotropic, and detoxifying functions. | Genetic variants in pathogenic genes (APOE, LRRK2), metabolic genes (CYP Family and NAT2) and detoxification genes (CYP1B1, GSTP1, and SOD2) influence the response of dopamine to AtreMorine. |
| Romero, A. et al., 2017 [61]. | Evaluation the neuroprotective and antiinflammatory effects of AtreMorine in in vitro models of Parkinson Disease and oxidative stress. 6- OHDA Parkinson model and rote/oligo, oxidative stress model in Neuroblastoma SH- SY5Y cells. 6- OHDA Parkinson model in rat striatal slices. OGD/Reox as an oxidative stress model in rat hippocampal slices. LPS as a model of neuroinflammation in BV2 microglia cells. | Atremorine showed neuroprotective and antiinflammatory effects in different in vitro models. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Corzo, L.; Fernández-Novoa, L.; Carrera, I.; Martínez, O.; Rodríguez, S.; Alejo, R.; Cacabelos, R. Nutrition, Health, and Disease: Role of Selected Marine and Vegetal Nutraceuticals. Nutrients 2020, 12, 747. https://doi.org/10.3390/nu12030747
Corzo L, Fernández-Novoa L, Carrera I, Martínez O, Rodríguez S, Alejo R, Cacabelos R. Nutrition, Health, and Disease: Role of Selected Marine and Vegetal Nutraceuticals. Nutrients. 2020; 12(3):747. https://doi.org/10.3390/nu12030747
Chicago/Turabian StyleCorzo, Lola, Lucía Fernández-Novoa, Iván Carrera, Olaia Martínez, Susana Rodríguez, Ramón Alejo, and Ramón Cacabelos. 2020. "Nutrition, Health, and Disease: Role of Selected Marine and Vegetal Nutraceuticals" Nutrients 12, no. 3: 747. https://doi.org/10.3390/nu12030747
APA StyleCorzo, L., Fernández-Novoa, L., Carrera, I., Martínez, O., Rodríguez, S., Alejo, R., & Cacabelos, R. (2020). Nutrition, Health, and Disease: Role of Selected Marine and Vegetal Nutraceuticals. Nutrients, 12(3), 747. https://doi.org/10.3390/nu12030747