Serum Metabolomics Profiling of Commercially Mixed Functional Foods—Effects in Beta-Amyloid Induced Rats Measured Using 1H NMR Spectroscopy

 ,
,  ,
,
Abstract
:1. Introduction
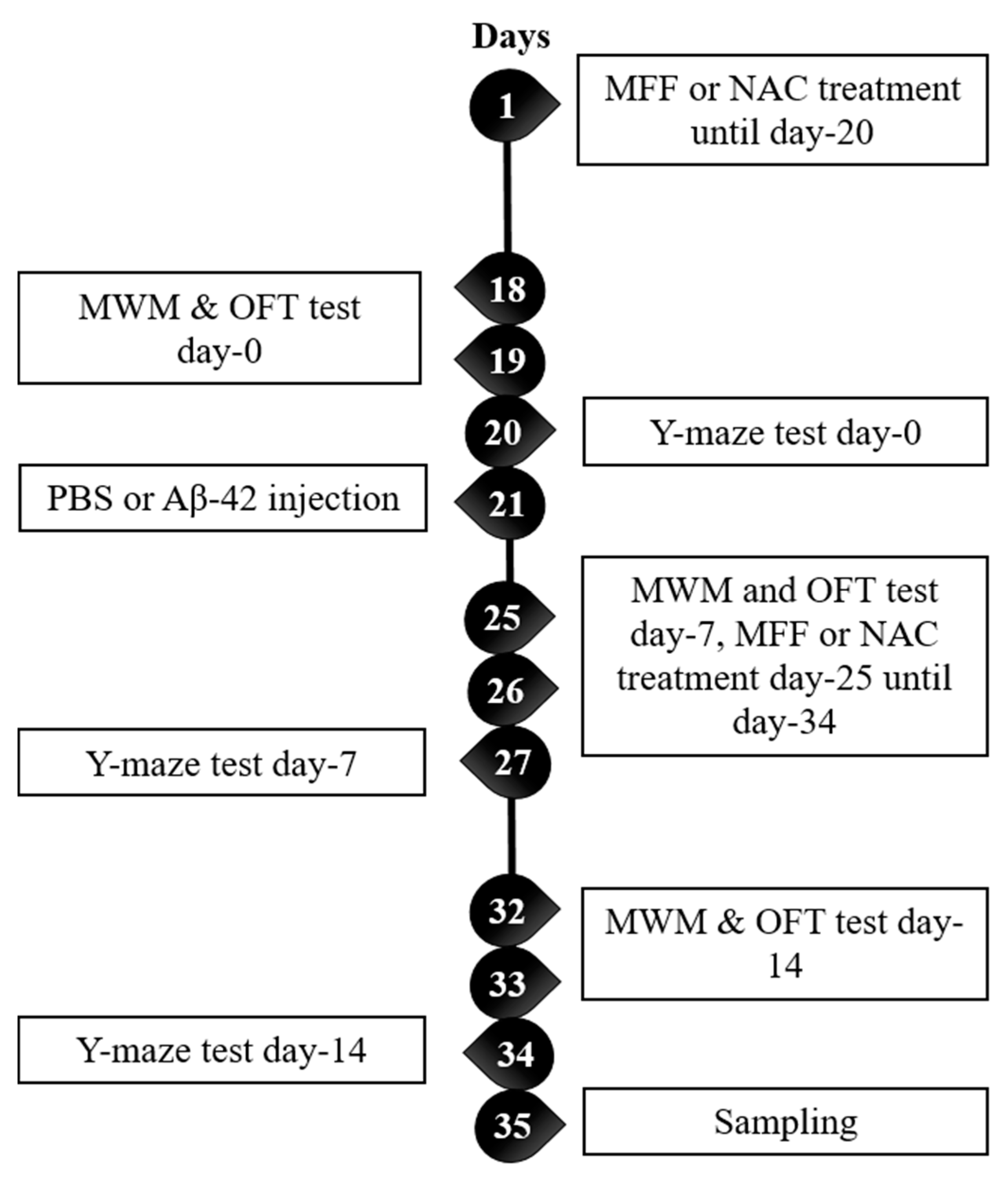
2. Materials and Methods
2.1. Animals
2.2. Preparation of Aβ-42 and Surgery
2.3. Morris Water Maze (MWM) Test
2.4. Open Field Test (OFT)
2.5. Y-Maze Test
2.6. Statistical Analysis for Behavioural Study
2.7. H NMR Spectroscopy
2.8. Statistical Analysis of 1H NMR Spectra
2.9. Pathway Analysis of 1H NMR Spectra
3. Results
3.1. Morris Water Maze (MWM) Test
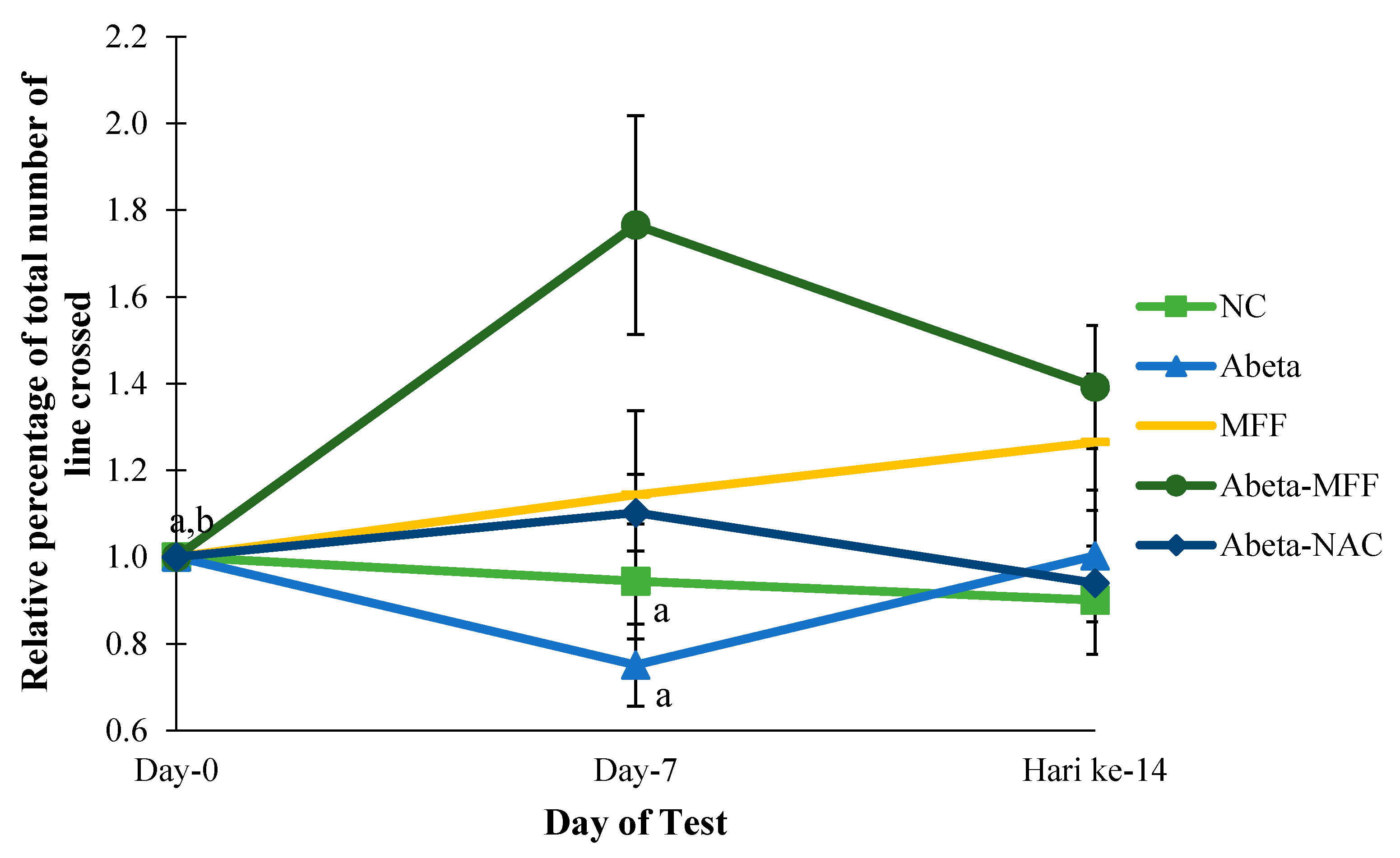
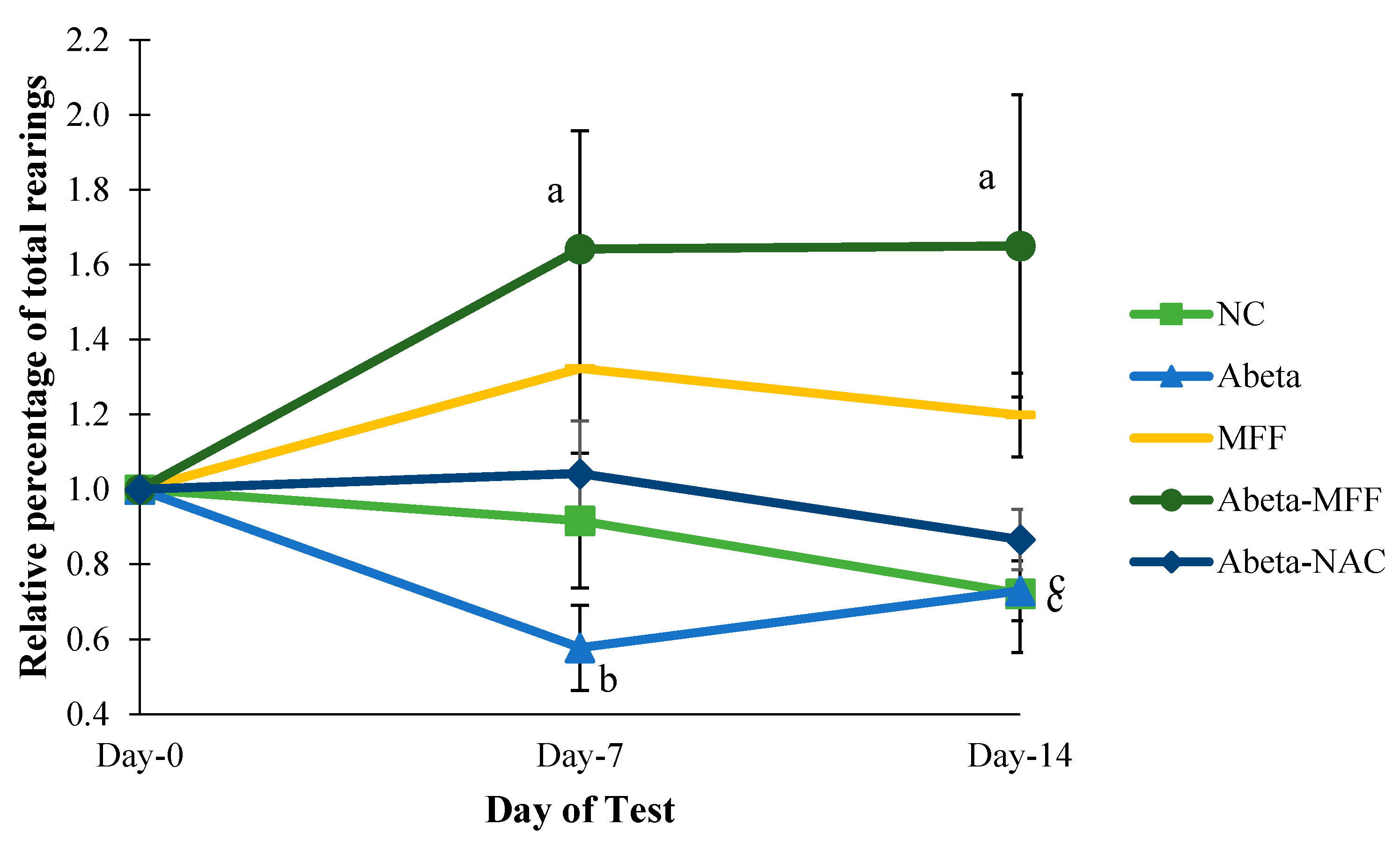
3.2. Open Field Test (OFT)
3.3. Y-Maze Test
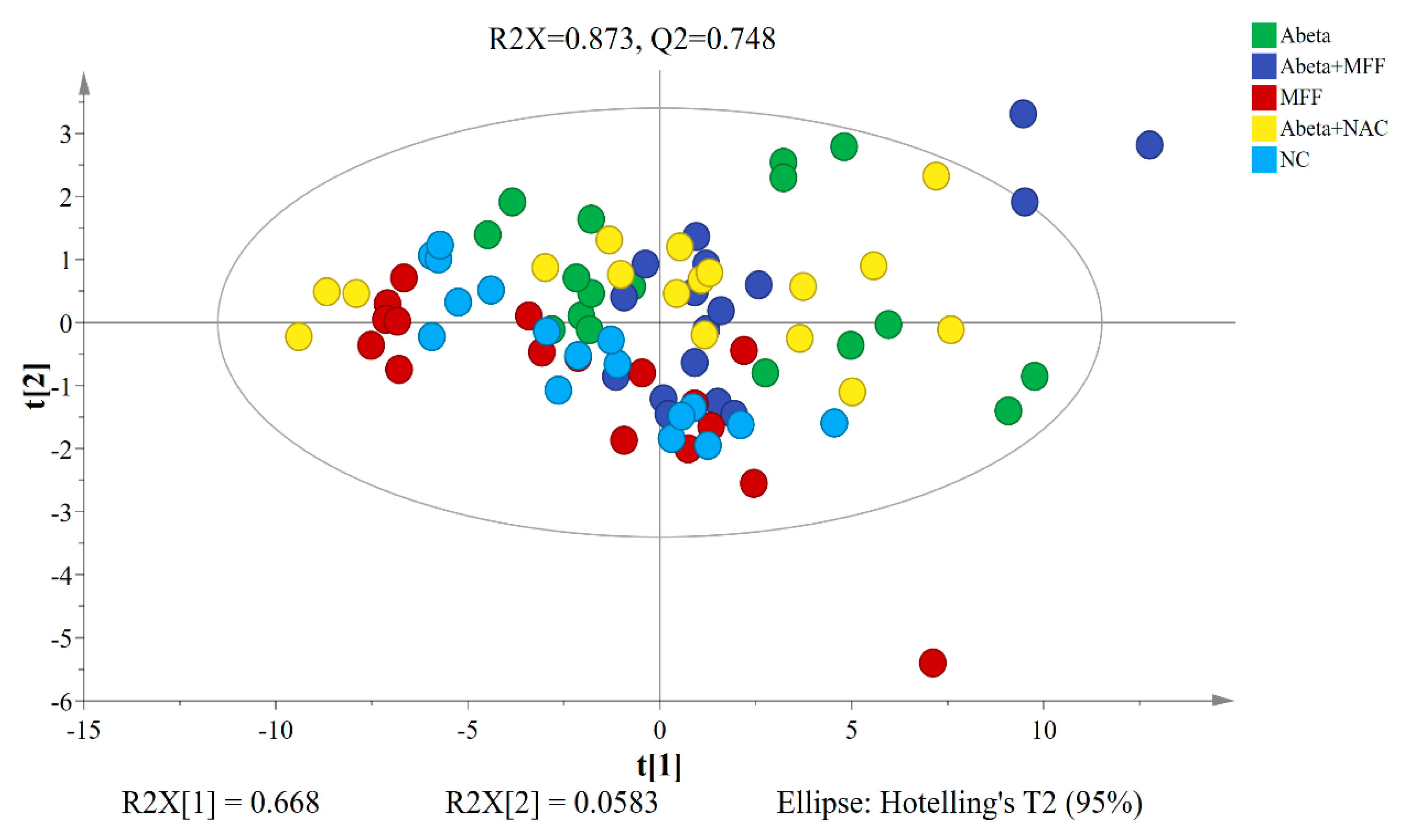
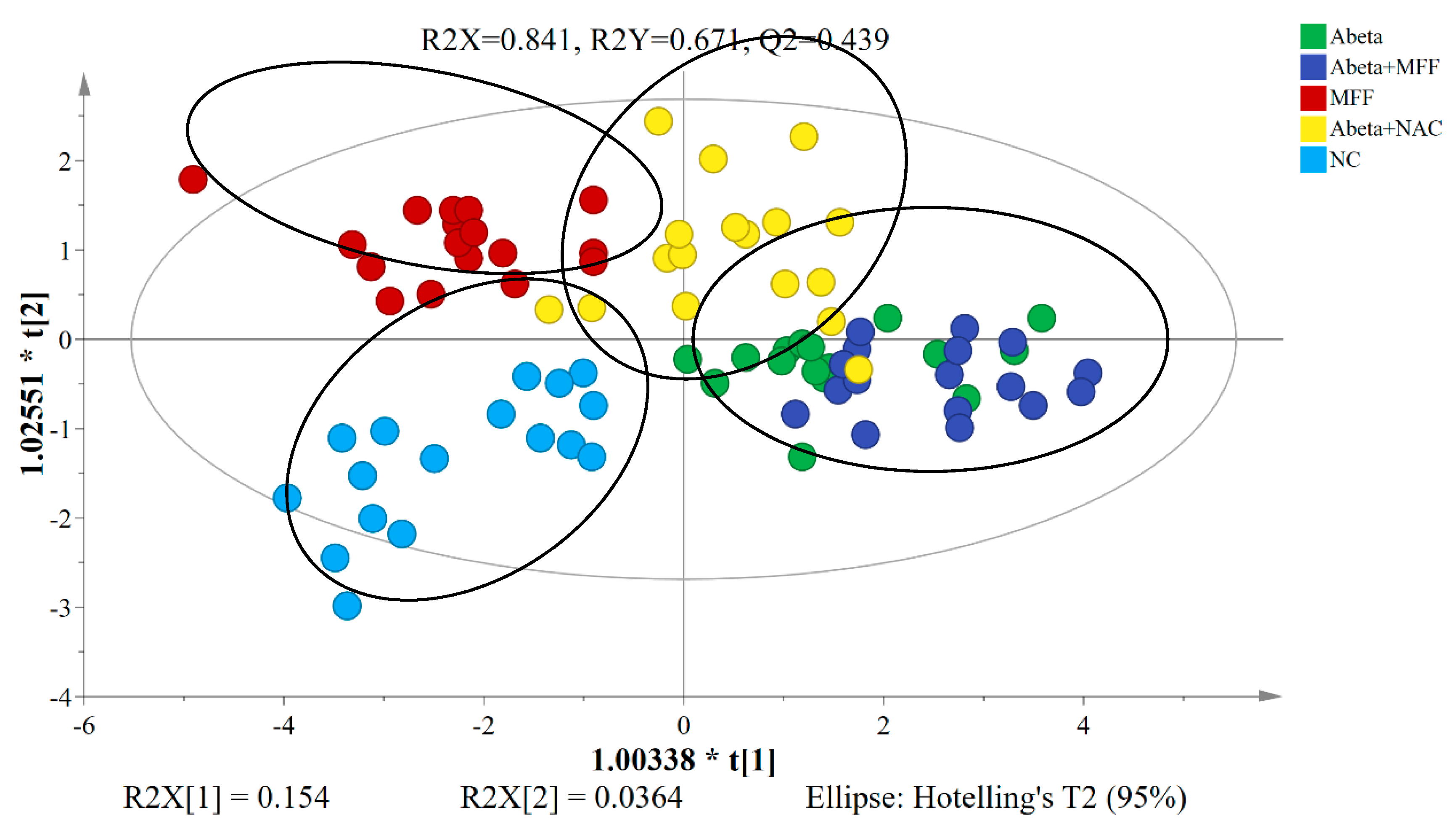
3.4. H NMR Metabolomic Analysis
3.5. Identification of Affected Metabolites
3.6. Metabolic Pathway Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jellinger, K.A.; Janetzky, B.; Attems, J.; Kienzl, E. Biomarkers for early diagnosis of Alzheimer disease: ‘ALZheimer ASsociated gene’—A new blood biomarker? J. Cell. Mol. Med. 2008, 12, 1094–1117. [Google Scholar] [CrossRef] [Green Version]
- Iriti, M.; Vitalini, S.; Fico, G.; Faoro, F. Neuroprotective Herbs and Foods from Different Traditional Medicines and Diets. Molecules 2010, 15, 3517–3555. [Google Scholar] [CrossRef] [Green Version]
- Reitz, C.; Brayne, C.; Mayeux, R. Epidemiology of Alzheimer disease. Nat. Rev. Neurol. 2011, 7, 137. [Google Scholar] [CrossRef]
- Gold, C.A.; Budson, A.E. Memory loss in Alzheimer’s disease: Implications for development of therapeutics. Expert Rev. Neurother. 2008, 8, 1879–1891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- García-Mesa, Y.; Colie, S.; Corpas, R.; Cristòfol, R.; Comellas, F.; Nebreda, A.R.; Giménez-Llort, L.; Sanfeliu, C. Oxidative Stress Is a Central Target for Physical Exercise Neuroprotection Against Pathological Brain Aging. J. Gerontol. Ser. A 2016, 71, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Corlier, F.; Hafzalla, G.; Faskowitz, J.; Kuller, L.H.; Becker, J.T.; Lopez, O.L.; Thompson, P.M.; Braskie, M.N. Systemic inflammation as a predictor of brain aging: Contributions of physical activity, metabolic risk, and genetic risk. NeuroImage 2018, 172, 118–129. [Google Scholar] [CrossRef] [PubMed]
- Bishop, N.A.; Lu, T.; Yankner, B.A. Neural mechanisms of ageing and cognitive decline. Nature 2010, 464, 529. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Golpich, M.; Amini, E.; Mohamed, Z.; Azman Ali, R.; Mohamed Ibrahim, N.; Ahmadiani, A. Mitochondrial Dysfunction and Biogenesis in Neurodegenerative diseases: Pathogenesis and Treatment. CNS Neurosci. Ther. 2017, 23, 5–22. [Google Scholar] [CrossRef]
- Akintola, A.; van Heemst, D. Chapter 65—Glucose, Insulin, and Human Brain Aging. In Conn’s Handbook of Models for Human Aging, 2nd ed.; Ram, J.L., Conn, P.M., Eds.; Academic Press: Cambridge, MA, USA, 2018; pp. 889–898. [Google Scholar] [CrossRef]
- De la Monte, S.M. Type 3 diabetes is sporadic Alzheimer’s disease: Mini-review. Eur. Neuropsychopharmacol. 2014, 24, 1954–1960. [Google Scholar] [CrossRef] [Green Version]
- Sheng, B.; Wang, X.; Su, B.; Lee, H.-G.; Casadesus, G.; Perry, G.; Zhu, X. Impaired mitochondrial biogenesis contributes to mitochondrial dysfunction in Alzheimer’s disease. J. Neurochem. 2012, 120, 419–429. [Google Scholar] [CrossRef]
- Li, J.; Zhao, X.-L.; Liu, Y.-X.; Peng, X.-H.; Zhu, S.-F.; Guo, H.; Liu, Y.-L.; Wan, M.-H.; Tang, W.-F. 1H NMR-based metabolomic profile of rats with experimental acute pancreatitis. BMC Gastroenterol. 2014, 14, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahmad Azam, A.; Ismail, I.S.; Shaikh, M.F.; Shaari, K.; Abas, F. Effects of Clinacanthus nutans leaf extract on lipopolysaccharide -induced neuroinflammation in rats: A behavioral and (1)H NMR-based metabolomics study. Avicenna J. Phytomedicine 2019, 9, 164–186. [Google Scholar]
- Du, Y.; Zheng, H.; Xia, H.; Zhao, L.; Hu, W.; Bai, G.; Yan, Z.; Gao, H. Early Effect of Amyloid β-Peptide on Hippocampal and Serum Metabolism in Rats Studied by an Integrated Method of NMR-Based Metabolomics and ANOVA-Simultaneous Component Analysis. Biomed Res. Int. 2017, 2017, 3262495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilkins, J.M.; Trushina, E. Application of Metabolomics in Alzheimer’s Disease. Front. Neurol. 2018, 8, 719. [Google Scholar] [CrossRef] [Green Version]
- Santos, C.Y.; Snyder, P.J.; Wu, W.-C.; Zhang, M.; Echeverria, A.; Alber, J. Pathophysiologic relationship between Alzheimer’s disease, cerebrovascular disease, and cardiovascular risk: A review and synthesis. Alzheimer’s Dement. Diagn. Assess. Dis. Monit. 2017, 7, 69–87. [Google Scholar] [CrossRef] [Green Version]
- Heppner, F.L.; Ransohoff, R.M.; Becher, B. Immune attack: The role of inflammation in Alzheimer disease. Nat. Rev. Neurosci. 2015, 16, 358. [Google Scholar] [CrossRef]
- Gil, M.I.; Tomás-Barberán, F.A.; Hess-Pierce, B.; Holcroft, D.M.; Kader, A.A. Antioxidant activity of pomegranate juice and its relationship with phenolic composition and processing. J. Agric. Food Chem. 2000, 48, 4581–4589. [Google Scholar] [CrossRef]
- Benmeddour, Z.; Mehinagic, E.; Meurlay, D.L.; Louaileche, H. Phenolic composition and antioxidant capacities of ten Algerian date (Phoenix dactylifera L.) cultivars: A comparative study. J. Funct. Foods 2013, 5, 346–354. [Google Scholar] [CrossRef]
- Socha, R.; Juszczak, L.; Pietrzyk, S.; Gałkowska, D.; Fortuna, T.; Witczak, T. Phenolic profile and antioxidant properties of Polish honeys. Int. J. Food Sci. Technol. 2011, 46, 528–534. [Google Scholar] [CrossRef]
- Poyrazoğlu, E.; Gökmen, V.; Artιk, N. Organic Acids and Phenolic Compounds in Pomegranates (Punica granatum L.) Grown in Turkey. J. Food Compos. Anal. 2002, 15, 567–575. [Google Scholar] [CrossRef]
- Ismail, N.I.; Abdul Kadir, M.R.; Mahmood, N.H.; Singh, O.P.; Iqbal, N.; Zulkifli, R.M. Apini and Meliponini foraging activities influence the phenolic content of different types of Malaysian honey. J. Apic. Res. 2016, 55, 137–150. [Google Scholar] [CrossRef]
- Manach, C.; Williamson, G.; Morand, C.; Scalbert, A.; Rémésy, C. Bioavailability and bioefficacy of polyphenols in humans. I. Review of 97 bioavailability studies. Am. J. Clin. Nutr. 2005, 81, 230S–242S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, W.; Breksa Iii, A.P.; Pan, Z.; Ma, H. Quantitative determination of major polyphenol constituents in pomegranate products. Food Chem. 2012, 132, 1585–1591. [Google Scholar] [CrossRef] [PubMed]
- González-Sarrías, A.; García-Villalba, R.; Núñez-Sánchez, M.Á.; Tomé-Carneiro, J.; Zafrilla, P.; Mulero, J.; Tomás-Barberán, F.A.; Espín, J.C. Identifying the limits for ellagic acid bioavailability: A crossover pharmacokinetic study in healthy volunteers after consumption of pomegranate extracts. J. Funct. Foods 2015, 19, 225–235. [Google Scholar] [CrossRef]
- Schaffer, S.; Halliwell, B. Do polyphenols enter the brain and does it matter? Some theoretical and practical considerations. Genes Nutr. 2012, 7, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Spencer, J.P.E. The impact of fruit flavonoids on memory and cognition. Br. J. Nutr. 2010, 104, S40–S47. [Google Scholar] [CrossRef] [Green Version]
- Adams, L.S.; Seeram, N.P.; Aggarwal, B.B.; Takada, Y.; Sand, D.; Heber, D. Pomegranate Juice, Total Pomegranate Ellagitannins, and Punicalagin Suppress Inflammatory Cell Signaling in Colon Cancer Cells. J. Agric. Food Chem. 2006, 54, 980–985. [Google Scholar] [CrossRef]
- Essa, M.M.; Subash, S.; Akbar, M.; Al-Adawi, S.; Guillemin, G.J. Long-Term Dietary Supplementation of Pomegranates, Figs and Dates Alleviate Neuroinflammation in a Transgenic Mouse Model of Alzheimer’s Disease. PLoS ONE 2015, 10, e0120964. [Google Scholar] [CrossRef] [Green Version]
- Abdulmajeed, W.I.; Sulieman, H.B.; Zubayr, M.O.; Imam, A.; Amin, A.; Biliaminu, S.A.; Oyewole, L.A.; Owoyele, B.V. Honey prevents neurobehavioural deficit and oxidative stress induced by lead acetate exposure in male wistar rats—A preliminary study. Metab. Brain Dis. 2016, 31, 37–44. [Google Scholar] [CrossRef]
- Erejuwa, O.O.; Sulaiman, S.A.; Wahab, M.S.; Sirajudeen, K.N.S.; Salleh, M.S.M.; Gurtu, S. Antioxidant protection of Malaysian tualang honey in pancreas of normal and streptozotocin-induced diabetic rats. Ann. D’endocrinologie 2010, 71, 291–296. [Google Scholar] [CrossRef]
- Feng, Y.; Yang, S.-G.; Du, X.-T.; Zhang, X.; Sun, X.-X.; Zhao, M.; Sun, G.-Y.; Liu, R.-T. Ellagic acid promotes Aβ42 fibrillization and inhibits Aβ42-induced neurotoxicity. Biochem. Biophys. Res. Commun. 2009, 390, 1250–1254. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.-J.; Jung, J.-S.; Kim, T.-K.; Hasan, M.A.; Hong, C.-W.; Nam, J.-S.; Song, D.-K. Protective Effects of Ferulic Acid in Amyloid Precursor Protein Plus Presenilin-1 Transgenic Mouse Model of Alzheimer Disease. Biol. Pharm. Bull. 2013, 36, 140–143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, R.H. Health benefits of fruit and vegetables are from additive and synergistic combinations of phytochemicals. Am. J. Clin. Nutr. 2003, 78, 517S–520S. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Jha, A.; Kumar, A.; Hettiarachchy, N.; Rai, A.K.; Sharma, D. Influence of the solvents on the extraction of major phenolic compounds (punicalagin, ellagic acid and gallic acid) and their antioxidant activities in pomegranate aril. J. Food Sci. Technol. 2014, 51, 2070–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Şenol, N.; Nazıroğlu, M.; Yürüker, V. N-Acetylcysteine and Selenium Modulate Oxidative Stress, Antioxidant Vitamin and Cytokine Values in Traumatic Brain Injury-Induced Rats. Neurochem. Res. 2014, 39, 685–692. [Google Scholar] [CrossRef] [PubMed]
- Ding, B.J.; Ma, W.W.; He, L.L.; Zhou, X.; Yuan, L.H.; Yu, H.L.; Feng, J.F.; Xiao, R. Soybean isoflavone alleviates β-amyloid 1-42 induced inflammatory response to improve learning and memory ability by down regulation of Toll-like receptor 4 expression and nuclear factor-κB activity in rats. Int. J. Dev. Neurosci. 2011, 29, 537–542. [Google Scholar] [CrossRef]
- Beheshti, S.; Soleimanipour, A. Prophylactic effect of all-trans retinoic acid in an amyloid-beta rat model of Alzheimer’s disease. Physiol. Pharmacol. 2017, 21, 34–43. [Google Scholar]
- Murphy, K.J.; Miller, A.-M.; Thelma, R.; Cowley, F.; Fionnuala Cox, F.; Lynch, M.A. The age- and amyloid-β-related increases in Nogo B contribute to microglial activation. Neurochem. Int. 2011, 58, 161–168. [Google Scholar] [CrossRef]
- Huang, T.-C.; Lu, K.-T.; Wo, Y.-Y.P.; Wu, Y.-J.; Yang, Y.-L. Resveratrol protects rats from Aβ-induced neurotoxicity by the reduction of iNOS expression and lipid peroxidation. PLoS ONE 2011, 6, e29102. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Wang, C.; Zhang, J.H.; Cai, J.-M.; Cao, Y.-P.; Sun, X.-J. Hydrogen-rich saline improves memory function in a rat model of amyloid-beta-induced Alzheimer’s disease by reduction of oxidative stress. Brain Res. 2010, 1328, 152–161. [Google Scholar] [CrossRef] [Green Version]
- Sturman, O.; Germain, P.-L.; Bohacek, J. Exploratory rearing: A context- and stress-sensitive behavior recorded in the open-field test. Stress 2018, 21, 443–452. [Google Scholar] [CrossRef] [PubMed]
- Bagheri, M.; Joghataei, M.-T.; Mohseni, S.; Roghani, M. Genistein ameliorates learning and memory deficits in amyloid β(1–40) rat model of Alzheimer’s disease. Neurobiol. Learn. Mem. 2011, 95, 270–276. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Le Guennec, A.; Tayyari, F.; Edison, A.S. Alternatives to Nuclear Overhauser Enhancement Spectroscopy Presat and Carr–Purcell–Meiboom–Gill Presat for NMR-Based Metabolomics. Anal. Chem. 2017, 89, 8582–8588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, S.F.; Holscher, C.; McClean, P.; Elliott, C.T.; Green, B.D. 1H NMR metabolomics investigation of an Alzheimer’s disease (AD) mouse model pinpoints important biochemical disturbances in brain and plasma. Metabolomics 2013, 9, 974–983. [Google Scholar] [CrossRef]
- Oskouie, A.A.; Yekta, R.F.; Tavirani, M.R.; Kashani, M.S.; Goshadrou, F. Lavandula angustifolia Effects on Rat Models of Alzheimer’s Disease Through the Investigation of Serum Metabolic Features Using NMR Metabolomics. Avicenna J. Med. Biotechnol. 2018, 10, 83–92. [Google Scholar] [PubMed]
- González-Domínguez, R.; Sayago, A.; Fernández-Recamales, Á. Metabolomics in Alzheimer’s disease: The need of complementary analytical platforms for the identification of biomarkers to unravel the underlying pathology. J. Chromatogr. B 2017, 1071, 75–92. [Google Scholar] [CrossRef]
- Trushina, E.; Mielke, M.M. Recent advances in the application of metabolomics to Alzheimer’s Disease. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2014, 1842, 1232–1239. [Google Scholar] [CrossRef] [Green Version]
- Barba, I.; Fernandez-Montesinos, R.; Garcia-Dorado, D.; Pozo, D. Alzheimer’s disease beyond the genomic era: Nuclear magnetic resonance (NMR) spectroscopy-based metabolomics. J. Cell. Mol. Med. 2008, 12, 1477–1485. [Google Scholar] [CrossRef] [Green Version]
- Butterfield, D.A.; Swomley, A.M.; Sultana, R. Amyloid β-Peptide (1–42)-Induced Oxidative Stress in Alzheimer Disease: Importance in Disease Pathogenesis and Progression. Antioxid. Redox Signal. 2012, 19, 823–835. [Google Scholar] [CrossRef] [Green Version]
- Oeckl, P.; Otto, M. A Review on MS-Based Blood Biomarkers for Alzheimer’s Disease. Neurol. Ther. 2019, 8, 113–127. [Google Scholar] [CrossRef] [Green Version]
- Lin, C.-N.; Huang, C.-C.; Huang, K.-L.; Lin, K.-J.; Yen, T.-C.; Kuo, H.-C. A metabolomic approach to identifying biomarkers in blood of Alzheimer’s disease. Ann. Clin. Transl. Neurol. 2019, 6, 537–545. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Humpel, C.; Hochstrasser, T. Cerebrospinal fluid and blood biomarkers in Alzheimer’s disease. World J. Psychiatry 2011, 1, 8. [Google Scholar] [CrossRef]
- Rasoolijazi, H.; Joghataie, M.T.; Roghani, M.; Nobakht, M. The Beneficial Effect of (-)-Epigallocatechin-3-Gallate in an Experimental Model of Alzheimer’s disease in Rat: A Behavioral Analysis. Iran. Biomed. J. 2007, 11, 237–243. [Google Scholar]
- Holcomb, L.; Gordon, M.N.; McGowan, E.; Yu, X.; Benkovic, S.; Jantzen, P.; Wright, K.; Saad, I.; Mueller, R.; Morgan, D.; et al. Accelerated Alzheimer-type phenotype in transgenic mice carrying both mutant amyloid precursor protein and presenilin 1 transgenes. Nat. Med. 1998, 4, 97–100. [Google Scholar] [CrossRef]
- Reisel, D.; Bannerman, D.M.; Schmitt, W.B.; Deacon, R.M.J.; Flint, J.; Borchardt, T.; Seeburg, P.H.; Rawlins, J.N.P. Spatial memory dissociations in mice lacking GluR1. Nat. Neurosci. 2002, 5, 868. [Google Scholar] [CrossRef]
- Arendash, G.W.; King, D.L.; Gordon, M.N.; Morgan, D.; Hatcher, J.M.; Hope, C.E.; Diamond, D.M. Progressive, age-related behavioral impairments in transgenic mice carrying both mutant amyloid precursor protein and presenilin-1 transgenes. Brain Res. 2001, 891, 42–53. [Google Scholar] [CrossRef]
- Prediger, R.D.S.; Franco, J.L.; Pandolfo, P.; Medeiros, R.; Duarte, F.S.; Di Giunta, G.; Figueiredo, C.P.; Farina, M.; Calixto, J.B.; Takahashi, R.N.; et al. Differential susceptibility following β-amyloid peptide-(1–40) administration in C57BL/6 and Swiss albino mice: Evidence for a dissociation between cognitive deficits and the glutathione system response. Behav. Brain Res. 2007, 177, 205–213. [Google Scholar] [CrossRef]
- Lever, C.; Burton, S.; Ο’Keefe, J. Rearing on Hind Legs, Environmental Novelty, and the Hippocampal Formation. Rev. Neurosci. 2006, 17, 111. [Google Scholar] [CrossRef]
- Bailey, K.R.; Crawley, J.N. Anxiety-related behaviors in mice. In Methods of Behavior Analysis in Neuroscience, 2nd ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis: Abingdon, UK, 2009. [Google Scholar]
- Griffin, J.W.D.; Bradshaw, P.C. Amino Acid Catabolism in Alzheimer’s Disease Brain: Friend or Foe? Oxidative Med. Cell. Longev. 2017, 2017, 5472792. [Google Scholar] [CrossRef] [Green Version]
- De Simone, R.; Vissicchio, F.; Mingarelli, C.; De Nuccio, C.; Visentin, S.; Ajmone-Cat, M.A.; Minghetti, L. Branched-chain amino acids influence the immune properties of microglial cells and their responsiveness to pro-inflammatory signals. Biochim. Et Biophys. Acta (BBA) Mol. Basis Dis. 2013; 1832, 650–659. [Google Scholar] [CrossRef] [Green Version]
- Parrella, E.; Maxim, T.; Maialetti, F.; Zhang, L.; Wan, J.; Wei, M.; Cohen, P.; Fontana, L.; Longo, V.D. Protein restriction cycles reduce IGF-1 and phosphorylated Tau, and improve behavioral performance in an Alzheimer’s disease mouse model. Aging Cell 2013, 12, 257–268. [Google Scholar] [CrossRef]
- Chen, J.; Zhang, C.; Wu, X.; Ji, H.; Ma, W.; Wei, S.; Zhang, L.; Chen, J. 1H NMR-based nontargeted metabonomics study of plasma and urinary biochemical changes in Kudouzi treated rats. Rev. Bras. Farmacogn. 2018, 28, 474–480. [Google Scholar] [CrossRef]
- Bonvallot, N.; Canlet, C.; Blas-Y-Estrada, F.; Gautier, R.; Tremblay-Franco, M.; Chevolleau, S.; Cordier, S.; Cravedi, J.-P. Metabolome disruption of pregnant rats and their offspring resulting from repeated exposure to a pesticide mixture representative of environmental contamination in Brittany. PLoS ONE 2018, 13, e0198448. [Google Scholar] [CrossRef]
- Chen, J.; Herrup, K. Glutamine Acts as a Neuroprotectant against DNA Damage, Beta-Amyloid and H2O2-Induced Stress. PLoS ONE 2012, 7, e33177. [Google Scholar] [CrossRef] [Green Version]
- Burns, C.M.; Chen, K.; Kaszniak, A.W.; Lee, W.; Alexander, G.E.; Bandy, D.; Fleisher, A.S.; Caselli, R.J.; Reiman, E.M. Higher serum glucose levels are associated with cerebral hypometabolism in Alzheimer regions. Neurology 2013, 80, 1557–1564. [Google Scholar] [CrossRef] [Green Version]
- Furst, A.J.; Rabinovici, G.D.; Rostomian, A.H.; Steed, T.; Alkalay, A.; Racine, C.; Miller, B.L.; Jagust, W.J. Cognition, glucose metabolism and amyloid burden in Alzheimer’s disease. Neurobiol. Aging 2012, 33, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Enche Ady, C.N.A.; Lim, S.M.; Teh, L.K.; Salleh, M.Z.; Chin, A.-V.; Tan, M.P.; Poi, P.J.H.; Kamaruzzaman, S.B.; Abdul Majeed, A.B.; Ramasamy, K. Metabolomic-guided discovery of Alzheimer’s disease biomarkers from body fluid. J. Neurosci. Res. 2017, 95, 2005–2024. [Google Scholar] [CrossRef]
- González-Domínguez, R.; García-Barrera, T.; Gómez-Ariza, J.L. Application of a novel metabolomic approach based on atmospheric pressure photoionization mass spectrometry using flow injection analysis for the study of Alzheimer’s disease. Talanta 2015, 131, 480–489. [Google Scholar] [CrossRef] [Green Version]
- Azam, A.A.; Pariyani, R.; Ismail, I.S.; Ismail, A.; Khatib, A.; Abas, F.; Shaari, K. Urinary metabolomics study on the protective role of Orthosiphon stamineus in Streptozotocin induced diabetes mellitus in rats via 1H NMR spectroscopy. BMC Complement. Altern. Med. 2017, 17, 278. [Google Scholar] [CrossRef] [Green Version]
- Jonckheere, A.I.; Smeitink, J.A.M.; Rodenburg, R.J.T. Mitochondrial ATP synthase: Architecture, function and pathology. J. Inherit. Metab. Dis. 2012, 35, 211–225. [Google Scholar] [CrossRef] [Green Version]
- Chen, R.; Wang, J.; Liao, C.; Ma, N.; Zhang, L.; Wang, X. 1H NMR studies on serum metabonomic changes over time in a kidney-Yang deficiency syndrome model. RSC Adv. 2017, 7, 34251–34261. [Google Scholar] [CrossRef] [Green Version]
- Ammar, N.M.; Farag, M.A.; Kholeif, T.E.; Metwally, N.S.; El-Sheikh, N.M.; El Gendy, A.N.; Abdel-Hamid, A.-H.Z. Serum metabolomics reveals the mechanistic role of functional foods and exercise for obesity management in rats. J. Pharm. Biomed. Anal. 2017, 142, 91–101. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, Y.; Lu, L.; Yang, Z.; Xue, W.; Tian, X.; Zhang, X. 1H NMR Based Serum Metabolomics Study to Investigate Hepatoprotective Effect of Qin-Jiao on Carbon Tetrachloride-Induced Acute Hepatotoxicity in Rats. Evid. Based Complement. Altern. Med. 2017, 2017, 6091589. [Google Scholar] [CrossRef]
- Suzuki, M.; Suzuki, M.; Sato, K.; Dohi, S.; Sato, T.; Matsuura, A.; Hiraide, A. Effect of β-Hydroxybutyrate, a Cerebral Function Improving Agent, on Cerebral Hypoxia, Anoxia and Ischemia in Mice and Rats. Jpn. J. Pharmacol. 2001, 87, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Kashiwaya, Y.; Takeshima, T.; Mori, N.; Nakashima, K.; Clarke, K.; Veech, R.L. d-β-Hydroxybutyrate protects neurons in models of Alzheimer’s and Parkinson’s disease. Proc. Natl. Acad. Sci. USA 2000, 97, 5440. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gasior, M.; Rogawski, M.A.; Hartman, A.L. Neuroprotective and disease-modifying effects of the ketogenic diet. Behav. Pharm. 2006, 17, 431–439. [Google Scholar] [CrossRef] [Green Version]
- Czech, C.; Berndt, P.; Busch, K.; Schmitz, O.; Wiemer, J.; Most, V.; Hampel, H.; Kastler, J.; Senn, H. Metabolite profiling of Alzheimer’s disease cerebrospinal fluid. PLoS ONE 2012, 7, e31501. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metabolites | Fold Change | |||
|---|---|---|---|---|
| NC vs. Abeta | MFF vs. Abeta | Abeta–MFF vs. Abeta | Abeta–NAC vs. Abeta | |
| Succinate | −0.83 * | −0.70 ** | +1.08 | −0.89 |
| Glutamine | −0.87 | −0.89 | +1.18 * | +1.03 |
| Pantothenate | −0.48 *** | −0.47 *** | +1.04 | −0.85 |
| Pyruvate | +1.29 * | −0.98 | +1.31 * | +1.07 |
| Citrate | +2.22 | +1.03 | +1.23 * | +1.03 |
| 3-hydroxybutyrate | +1.03 | +1.18 | +1.25 * | +1.2 * |
| Leucine | −0.64 | −0.55 * | −0.72 | −0.62 |
| Alanine | 1.00 | −0.72 *** | +1.08 | −0.90 |
| Choline | −0.95 | −0.60 ** | +1.03 | −0.46 *** |
| Lactate | −0.91 | −0.70 ** | +1.07 | −0.9 |
| Glucose | −0.72 ** | −0.83 | +1.07 | −0.98 |
| O-acetylcholine | −0.91 | −0.72 ** | +1.11 | −0.8 * |
| Pathways | P (Raw P) | −Log (p) | * FDR | Impact |
|---|---|---|---|---|
| Valine, leucine and isoleucine biosynthesis | 0.00354 | 5.64 | 0.0717 | 0.33 |
| Alanine, aspartate and glutamate metabolism | 0.0000299 | 10.4 | 0.00242 | 0.15 |
| Citrate cycle (TCA cycle) | 0.000504 | 7.59 | 0.0136 | 0.15 |
| Pyruvate metabolism | 0.0141 | 4.26 | 0.225 | 0.19 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rosli, N.H.M.; Yahya, H.M.; Ibrahim, F.W.; Shahar, S.; Ismail, I.S.; Azam, A.A.; Rajab, N.F. Serum Metabolomics Profiling of Commercially Mixed Functional Foods—Effects in Beta-Amyloid Induced Rats Measured Using 1H NMR Spectroscopy. Nutrients 2020, 12, 3812. https://doi.org/10.3390/nu12123812
Rosli NHM, Yahya HM, Ibrahim FW, Shahar S, Ismail IS, Azam AA, Rajab NF. Serum Metabolomics Profiling of Commercially Mixed Functional Foods—Effects in Beta-Amyloid Induced Rats Measured Using 1H NMR Spectroscopy. Nutrients. 2020; 12(12):3812. https://doi.org/10.3390/nu12123812
Chicago/Turabian StyleRosli, Nur Hasnieza Mohd, Hanis Mastura Yahya, Farah Wahida Ibrahim, Suzana Shahar, Intan Safinar Ismail, Amalina Ahmad Azam, and Nor Fadilah Rajab. 2020. "Serum Metabolomics Profiling of Commercially Mixed Functional Foods—Effects in Beta-Amyloid Induced Rats Measured Using 1H NMR Spectroscopy" Nutrients 12, no. 12: 3812. https://doi.org/10.3390/nu12123812
APA StyleRosli, N. H. M., Yahya, H. M., Ibrahim, F. W., Shahar, S., Ismail, I. S., Azam, A. A., & Rajab, N. F. (2020). Serum Metabolomics Profiling of Commercially Mixed Functional Foods—Effects in Beta-Amyloid Induced Rats Measured Using 1H NMR Spectroscopy. Nutrients, 12(12), 3812. https://doi.org/10.3390/nu12123812