Subacute Ingestion of Caffeine and Oolong Tea Increases Fat Oxidation without Affecting Energy Expenditure and Sleep Architecture: A Randomized, Placebo-Controlled, Double-Blinded Cross-Over Trial


Abstract
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Experimental Beverages
2.3. Protocol
2.4. Energy Metabolism Measurements
2.5. Activity and Sleep Recording
2.6. Thermometry
2.7. Autonomic Nervous System Activity
2.8. Statistical Analysis
3. Results
3.1. Physical Characteristics of the Subjects
3.2. Energy Metabolism
3.3. Core Body Temperature, Autonomic Nervous System Activity, and Sleep Architecture
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Liu, P.P.; Yin, J.F.; Chen, G.S.; Wang, F.; Xu, Y.Q. Flavor characteristics and chemical compositions of oolong tea processed using different semi-fermentation times. J. Food Sci. Technol. 2018, 55, 1185–1195. [Google Scholar] [CrossRef]
- Dulloo, A.G.; Geissler, C.A.; Horton, T.; Collins, A.; Miller, D.S. Normal caffeine consumption: Influence on thermogenesis and daily energy expenditure in lean and postobese human volunteers. Am. J. Clin. Nutr. 1989, 49, 44–50. [Google Scholar] [CrossRef]
- Bracco, D.; Ferrarra, J.M.; Arnaud, M.J.; Jéquier, E.; Schutz, Y. Effects of caffeine on energy metabolism, heart rate, and methylxanthine metabolism in lean and obese women. Am. J. Physiol. 1995, 269, E671–E678. [Google Scholar] [CrossRef]
- Rumpler, W.; Seale, J.; Clevidence, B.; Judd, J.; Wiley, E.; Yamamoto, S.; Komatsu, T.; Sawaki, T.; Ishikura, Y.; Hosoda, K. Oolong tea increases metabolic rate and fat oxidation in men. J. Nutr. 2001, 131, 2848–4852. [Google Scholar] [CrossRef]
- Dulloo, A.G.; Duret, C.; Rohrer, D.; Girardier, L.; Mensi, N.; Fathi, M.; Chantre, P.; Vandermander, J. Efficacy of a green tea extract rich in catechin polyphenols and caffeine in increasing 24-h energy expenditure and fat oxidation in humans. Am. J. Clin. Nutr. 1999, 70, 1040–1045. [Google Scholar] [CrossRef]
- Robertson, D.; Wade, D.; Workman, R.; Woosley, R.L.; Oates, J.A. Tolerance to the humoral and hemodynamic effects of caffeine in man. J. Clin. Investig. 1981, 67, 1111–1117. [Google Scholar] [CrossRef]
- Lazarus, M.; Shen, H.Y.; Cherasse, Y.; Qu, W.M.; Huang, Z.L.; Bass, C.E.; Winsky-Sommerer, R.; Semba, K.; Fredholm, B.B.; Boison, D.; et al. Arousal effect of caffeine depends on adenosine A2A receptors in the shell of the nucleus accumbens. J. Neurosci. 2011, 31, 10067–10075. [Google Scholar] [CrossRef]
- Lazarus, M.; Oishi, Y.; Bjorness, T.E.; Greene, R.W. Gating and the need for sleep: Dissociable effects of adenosine A1and A2A receptors. Front. Neurosci. 2019, 13, 740. [Google Scholar] [CrossRef]
- Nielsen, L.S.; Danielsen, K.V.; Sørensen, T.I. Short sleep duration as a possible cause of obesity: Critical analysis of the epidemiological evidence. Obes. Rev. 2011, 12, 78–92. [Google Scholar] [CrossRef]
- Chaput, J.P.; Leblanc, C.; Pérusse, L.; Després, J.P.; Bouchard, C.; Angelo Tremblay, A. Risk factors for adult overweight and obesity in the Quebec Family Study: Have we been barking up the wrong tree? Obesity 2009, 17, 1964–1970. [Google Scholar] [CrossRef]
- Cappuccio, F.P.; Taggart, F.M.; Kandala, N.B.; Currie, A.; Peile, E.; Stranges, S.; Miller, M.A. Meta-analysis of short sleep duration and obesity in children and adults. Sleep 2008, 31, 619–626. [Google Scholar] [CrossRef]
- Taheri, S.; Lin, L.; Austin, D.; Young, T.; Mignot, E. Short sleep duration is associated with reduced leptin, elevated ghrelin, and increased body mass index. PLoS Med. 2004, 1, e62. [Google Scholar] [CrossRef]
- Gangwisch, J.E.; Malaspina, D.; Boden-Albala, B.; Heymsfield, S.B. Inadequate sleep as a risk factor for obesity: Analysis of the NHANES I. Sleep 2005, 28, 1289–1296. [Google Scholar] [CrossRef]
- Knutson, K.L.; Spiegel, K.; Penev, P.; Van Cauter, E. The metabolic consequences of sleep deprivation. Sleep Med. Rev. 2007, 11, 163–178. [Google Scholar] [CrossRef]
- Nicolaidis, S. Metabolic mechanism of wakefulness (and hunger) and sleep (and satiety). Metabolism 2006, 55, S24–S29. [Google Scholar] [CrossRef]
- Sakurai, T. The role of orexin in motivated behaviours. Nat. Rev. Neurosci. 2014, 15, 719–731. [Google Scholar] [CrossRef]
- Tokuyama, K.; Ogata, H.; Katayose, Y.; Satoh, M. Algorithm for transient response of whole body indirect calorimeter: Deconvolution with a regularization parameter. J. Appl. Physiol. 2009, 106, 640–650. [Google Scholar] [CrossRef]
- Nakai, M.; Fukui, Y.; Asami, S.; Toyoda-Ono, Y.; Iwashita, T.; Shibata, H.; Mitsunaga, T.; Hashimoto, F.; Kiso, Y. Inhibitory effects of oolong tea polyphenols on pancreatic lipase in vitro. J. Agric. Food Chem. 2005, 53, 4593–4598. [Google Scholar] [CrossRef]
- Ministry of Health, Labour and Welfare of Japan. The National Health and Nutrition Survey. 2015. Available online: https://www.mhlw.go.jp/file/04-Houdouhappyou-10904750-Kenkoukyoku-Gantaisakukenkouzoushinka/0000041955.pdf (accessed on 11 September 2020). (In Japanese)
- Seol, J.; Fujii, Y.; Park, I.; Suzuki, Y.; Kawana, F.; Yajima, K.; Fukusumi, S.; Okura, T.; Satoh, M.; Tokuyama, K.; et al. Distinct effects of orexin receptor antagonist and GABAA agonist on sleep and physical/cognitive functions after forced awakening. Proc. Natl. Acad. Sci. USA 2019, 116, 24353–24358. [Google Scholar] [CrossRef]
- Park, I.; Ochiai, R.; Ogata, H.; Kayaba, M.; Hari, S.; Hibi, M.; Katsuragi, Y.; Satoh, M.; Tokuyama, K. Effects of subacute ingestion of chlorogenic acids on sleep architecture and energy metabolism through activity of the autonomic nervous system: A randomised, placebo-controlled, double-blinded crossover trial. Br. J. Nutr. 2017, 117, 979–984. [Google Scholar] [CrossRef]
- Ferrannini, E. The theoretical bases of indirect calorimetry: A review. Metabolism 1988, 37, 287–301. [Google Scholar] [CrossRef]
- Matsuo, M.; Masuda, F.; Sumi, Y.; Takahashi, M.; Yamada, N.; Hasegawa-Ohira, M.; Koichi Fujiwara, K.; Kanemura, T.; Kadotani, H. Comparisons of portable sleep monitors of different modalities: Potential as naturalistic aleep recorders. Front. Neurol. 2016, 7, 110. [Google Scholar] [CrossRef]
- Zhang, S.; Osumi, H.; Uchizawa, A.; Hamada, H.; Park, I.; Suzuki, Y.; Tanaka, Y.; Ishihara, A.; Yajima, K.; Seol, J.; et al. Changes in sleeping energy metabolism and thermoregulation during menstrual cycle. Physiol. Rep. 2020, 8, e14353. [Google Scholar] [CrossRef]
- Berry, R.B.; Albertario, C.L.; Harding, S.M.; Lloyd, R.M.; Plante, D.T.; Quan, S.F.; Troester, M.M.; Vaughn, B.V.; American Academy of Sleep Medicine. The AASM Manual for the Scoring of Sleep and Associated Events: Rules, Terminology and Technical Specifications; Version 2.5; American Academy of Sleep Medicine: Darien, IL, USA, 2018. [Google Scholar]
- Ogata, H.; Horie, M.; Kayaba, M.; Tanaka, Y.; Ando, A.; Park, I.; Zhang, S.; Yajima, K.; Shoda, J.; Omi, N.; et al. Skipping breakfast for 6 days delayed the circadian rhythm of the body temperature but did not alter the peak time of clock gene expression in human leukocytes. Nutrients 2020, 12, 2797. [Google Scholar] [CrossRef]
- Task Force of the European Society of Cardiology and the North American Society of Pacing and Electrophysiology. Heart rate variability: Standards of measurement, physiological interpretation and clinical use. Circulation 1996, 93, 1043–1065. [Google Scholar] [CrossRef]
- Acheson, K.J.; Gremaud, G.; Meirim, I.; Montigon, F.; Krebs, Y.; Fay, L.B.; Gay, L.-J.; Schneiter, P.; Schindler, C.; Tappy, L. Metabolic effects of caffeine in humans: Lipid oxidation or futile cycling? Am. J. Clin. Nutr. 2004, 79, 40–46. [Google Scholar] [CrossRef]
- LeBlanc, J.; Jobin, M.; Côté, J.; Samson, P.; Labrie, A. Enhanced metabolic response to caffeine in exercise-trained human subjects. J. Appl. Physiol. 1985, 59, 832–837. [Google Scholar] [CrossRef]
- Poehlman, E.T.; Després, J.P.; Bessette, H.; Fontaine, E.; Tremblay, A.; Bouchard, C. Influence of caffeine on the resting metabolic rate of exercise-trained and inactive subjects. Med. Sci. Sports Exerc. 1985, 17, 689–694. [Google Scholar] [CrossRef]
- Astrup, A.; Toubro, S.; Cannon, S.; Hein, P.; Breum, L.; Madsen, J. Caffeine: A double-blind, placebo-controlled study of its thermogenic, metabolic, and cardiovascular effects in healthy volunteers. Am. J. Clin. Nutr. 1990, 51, 759–767. [Google Scholar] [CrossRef]
- Acheson, K.J.; Zahorska-Markiewicz, B.; Pittet, P.; Anantharaman, K.; Jéquier, E. Caffeine and coffee: Their influence on metabolic rate and substrate utilization in normal weight and obese individuals. Am. J. Clin. Nutr. 1980, 33, 989–997. [Google Scholar] [CrossRef]
- Jung, R.T.; Shetty, P.S.; James, W.P.; Barrand, M.A.; Callingham, B.A. Caffeine: Its effect on catecholamines and metabolism in lean and obese humans. Clin. Sci. 1981, 60, 527–535. [Google Scholar] [CrossRef] [PubMed]
- Harpaz, E.; Tamir, S.; Weinstein, A.; Weinstein, Y. The effect of caffeine on energy balance. J. Basic Clin. Physiol. Pharmacol. 2017, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Landolt, H.P. Caffeine, the circadian clock, and sleep. Science 2015, 349, 1289. [Google Scholar] [CrossRef] [PubMed]
- Zwyghuizen-Doorenbos, A.; Roehrs, T.A.; Lipschutz, L.; Timms, V.; Roth, T. Effects of caffeine on alertness. Psychopharmacology 1990, 100, 36–39. [Google Scholar] [CrossRef]
- Yamada, M.; Sasaki, S.; Murakami, K.; Takahashi, Y.; Okubo, H.; Hirota, N.; Notsu, A.; Todoriki, H.; Miura, A.; Fukui, M.; et al. Estimation of caffeine intake in Japanese adults using 16 d weighed diet records based on a food composition database newly developed for Japanese populations. Public Health Nutr. 2009, 13, 663–672. [Google Scholar] [CrossRef]
- Komatsu, T.; Nakamori, M.; Komatsu, K.; Hosoda, K.; Okamura, M.; Toyama, K.; Ishikura, Y.; Sakai, T.; Kunii, D.; Yamamoto, S. Oolong tea increases energy metabolism in Japanese females. J. Med. Investig. 2003, 50, 170–175. [Google Scholar]
- Bernard, E.; Statland, T.J. Demas. Serum caffeine half-lives: Healthy subjects vs. patients having alcoholic hepatic disease. Am. J. Clin. Pathol. 1980, 73, 390–393. [Google Scholar]
- Begaye, B.; Vinales, K.L.; Hollstein, T.; Ando, T.; Walter, M.; Bogardus, C.; Krakoff, J.; Piaggi, P. Impaired metabolic flexibility to high-fat overfeeding predicts future weight gain in healthy adults. Diabetes 2020, 69, 181–192. [Google Scholar] [CrossRef]
- Mynatt, R.L.; Noland, R.C.; Elks, C.M.; Vandanmagsar, B.; Bayless, D.S.; Stone, A.C.; Ghosh, S.; Ravussin, E.; Warfel, J.D. The RNA binding protein HuR influences skeletal muscle metabolic flexibility in rodents and humans. Metabolism 2019, 97, 40–49. [Google Scholar] [CrossRef]
- Kräuchi, K.; Konieczka, K.; Roescheisen-Weich, C.; Gompper, B.; Hauenstein, D.; Schoetzau, A.; Fraenkl, S.; Flammer, J. Diurnal and menstrual cycles in body temperature are regulated differently: A 28-day ambulatory study in healthy women with thermal discomfort of cold extremities and controls. Chronobiol. Intern. 2014, 31, 102–113. [Google Scholar] [CrossRef]
- Lonac, M.C.; Richards, J.C.; Schweder, M.M.; Johnson, T.K.; Bell, C. Influence of short-term consumption of the caffeine-free, epigallocatechin-3-gallate supplement, Teavigo, on resting metabolism and the thermic effect of feeding. Obesity 2011, 19, 298–304. [Google Scholar] [CrossRef] [PubMed]
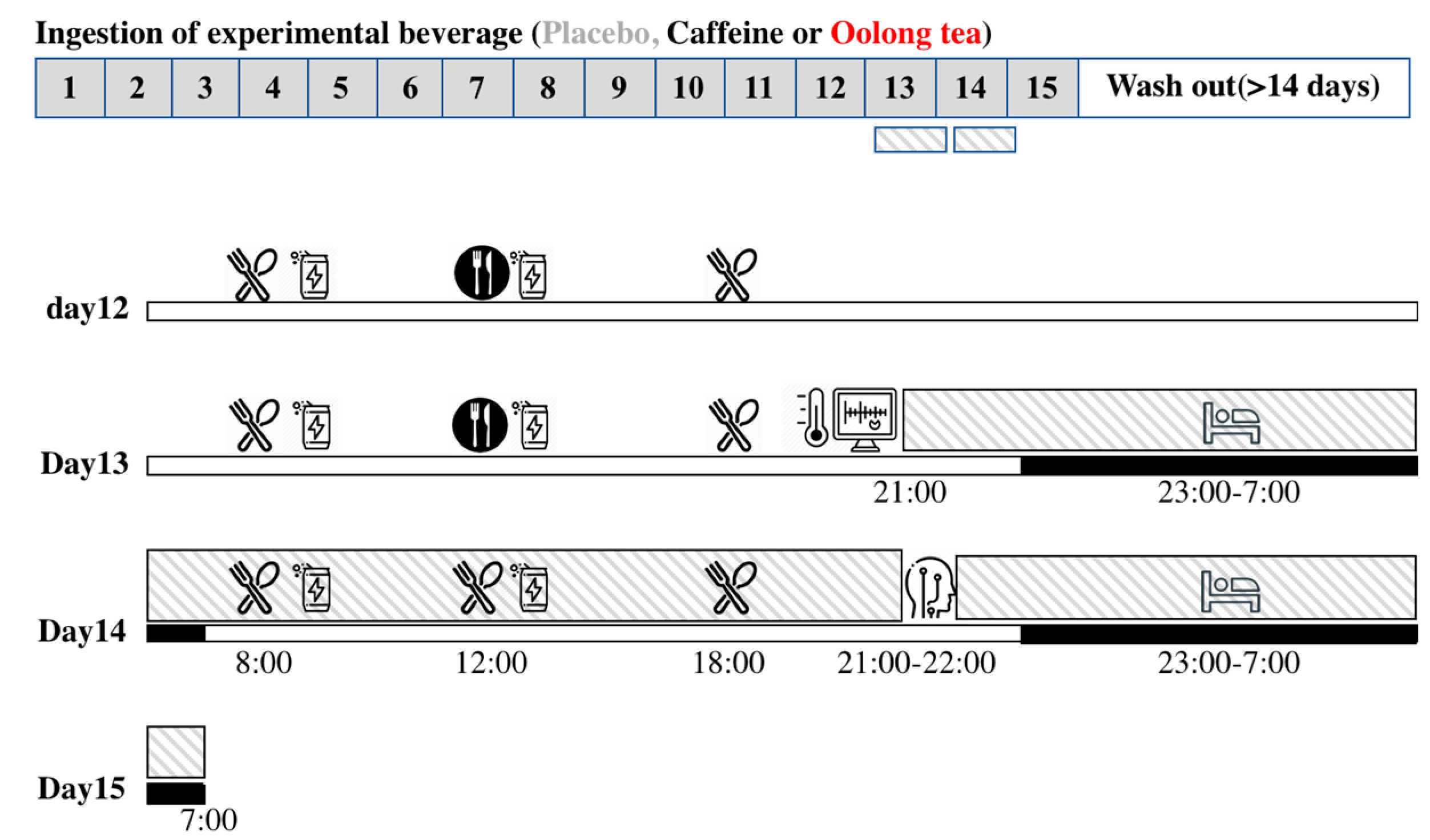
 ), but the lunch menu was not controlled (
), but the lunch menu was not controlled ( ). On day 13 of the session, the subjects wore sensors for heart rate, swallowed a core body temperature sensor, and then entered the whole room indirect calorimeter (
). On day 13 of the session, the subjects wore sensors for heart rate, swallowed a core body temperature sensor, and then entered the whole room indirect calorimeter ( ), where they slept for 8 h from 23:00 to 7:00 (
), where they slept for 8 h from 23:00 to 7:00 ( ). On day 14 of the session, experimental meals were provided as breakfast at 8:00, lunch at 12:00, and dinner at 18:00. Indirect calorimetry was interrupted from 21:00 of the 14th day of the session for preparing polysomnographic measurement.
), but the lunch menu was not controlled (). On day 13 of the session, the subjects wore sensors for heart rate, swallowed a core body temperature sensor, and then entered the whole room indirect calorimeter (), where they slept for 8 h from 23:00 to 7:00 (). On day 14 of the session, experimental meals were provided as breakfast at 8:00, lunch at 12:00, and dinner at 18:00. Indirect calorimetry was interrupted from 21:00 of the 14th day of the session for preparing polysomnographic measurement.
). On day 14 of the session, experimental meals were provided as breakfast at 8:00, lunch at 12:00, and dinner at 18:00. Indirect calorimetry was interrupted from 21:00 of the 14th day of the session for preparing polysomnographic measurement.
), but the lunch menu was not controlled (). On day 13 of the session, the subjects wore sensors for heart rate, swallowed a core body temperature sensor, and then entered the whole room indirect calorimeter (), where they slept for 8 h from 23:00 to 7:00 (). On day 14 of the session, experimental meals were provided as breakfast at 8:00, lunch at 12:00, and dinner at 18:00. Indirect calorimetry was interrupted from 21:00 of the 14th day of the session for preparing polysomnographic measurement.




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Placebo | Caffeine | Oolong Tea | |
|---|---|---|---|
| Caffeine | 0.0 | 51.8 | 51.8 |
| Gallic acid | 0.0 | 0.0 | 10.7 |
| Catechins | |||
| Cathechin | 0.0 | 0.0 | 3.4 |
| Gallocatechin | 0.0 | 0.0 | 12.3 |
| Catechin gallate | 0.0 | 0.0 | 2.9 |
| Gallocatechin gallate | 0.0 | 0.0 | 10.5 |
| Epicatechin | 0.0 | 0.0 | 1.8 |
| Epigallocatechin | 0.0 | 0.0 | 5.2 |
| Epicatechin gallate | 0.0 | 0.0 | 2.4 |
| Epigallocatechin gallate | 0.0 | 0.0 | 10.0 |
| Other polyphenols including | |||
| polymerized polyphenols | 0.0 | 0.0 | 62.3 |
| Placebo | Caffeine | Oolong Tea | |
|---|---|---|---|
| Age (years) | 37.1 ± 4.3 | ||
| Height (cm) | 168.2 ± 1.7 | ||
| PSQI-J | 1.4 ± 0.5 | ||
| MEQ-J | 53.0 ± 2.5 | ||
| Weight (kg) | 62.0 ± 1.5 | 61.6 ± 1.6 | 61.6 ± 1.5 |
| BMI (kg/m2) | 21.9 ± 0.6 | 21.8 ± 0.5 | 21.8± 0.5 |
| Body fat (%) | 18.5 ± 1.0 | 17.8± 1.0 | 17.5± 1.1 |
| Body water (%) | 54.9 ± 1.1 | 55.5 ± 1.1 | 56.5 ± 1.4 |
| Physical activity (kcal/day) | 1843 ± 63 | 1850 ± 49 | 1817 ± 50 |
| Placebo | Caffeine | Oolong | Df # | F # | p-Value # | |
|---|---|---|---|---|---|---|
| 24 h | ||||||
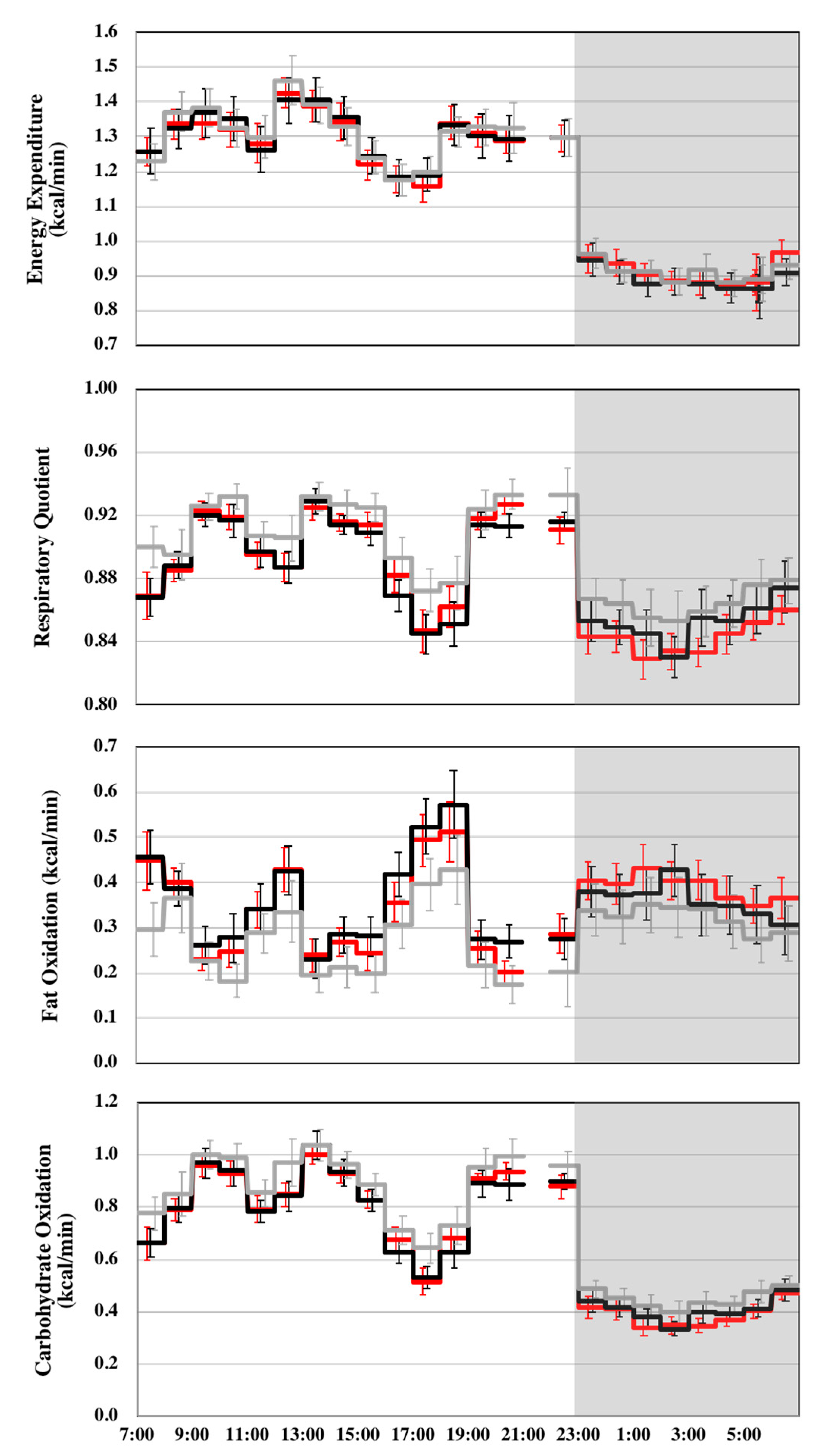
| Energy expenditure (kcal/min) | 1.17 ± 0.26 | 1.16 ± 0.27 | 1.16 ± 0.24 | 748.0 | 1.175 | p = 0.309 |
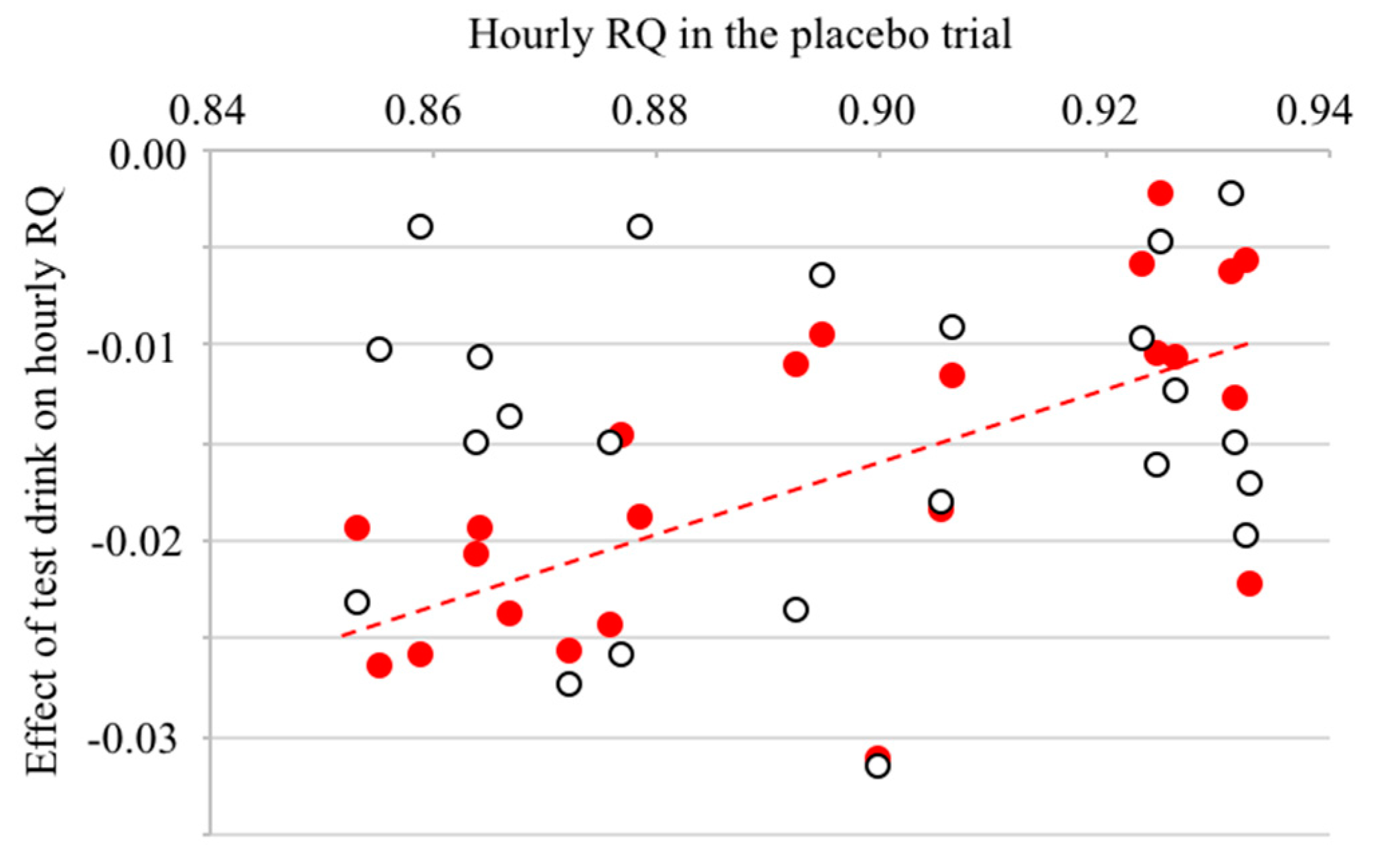
| RQ | 0.90 ± 0.05 | 0.88 ± 0.05 ** | 0.88 ± 0.047 ** | 748.0 | 24.792 | p < 0.001 |
| Fat oxidation (kcal/min) | 0.29 ± 0.20 | 0.36 ± 0.20 ** | 0.35 ± 0.17 ** | 748.0 | 26.484 | p < 0.001 |
| Carbohydrate oxidation (kcal/min) | 0.74 ± 0.29 | 0.68 ± 0.27 ** | 0.67 ± 0.27 ** | 748.0 | 18.437 | p < 0.001 |
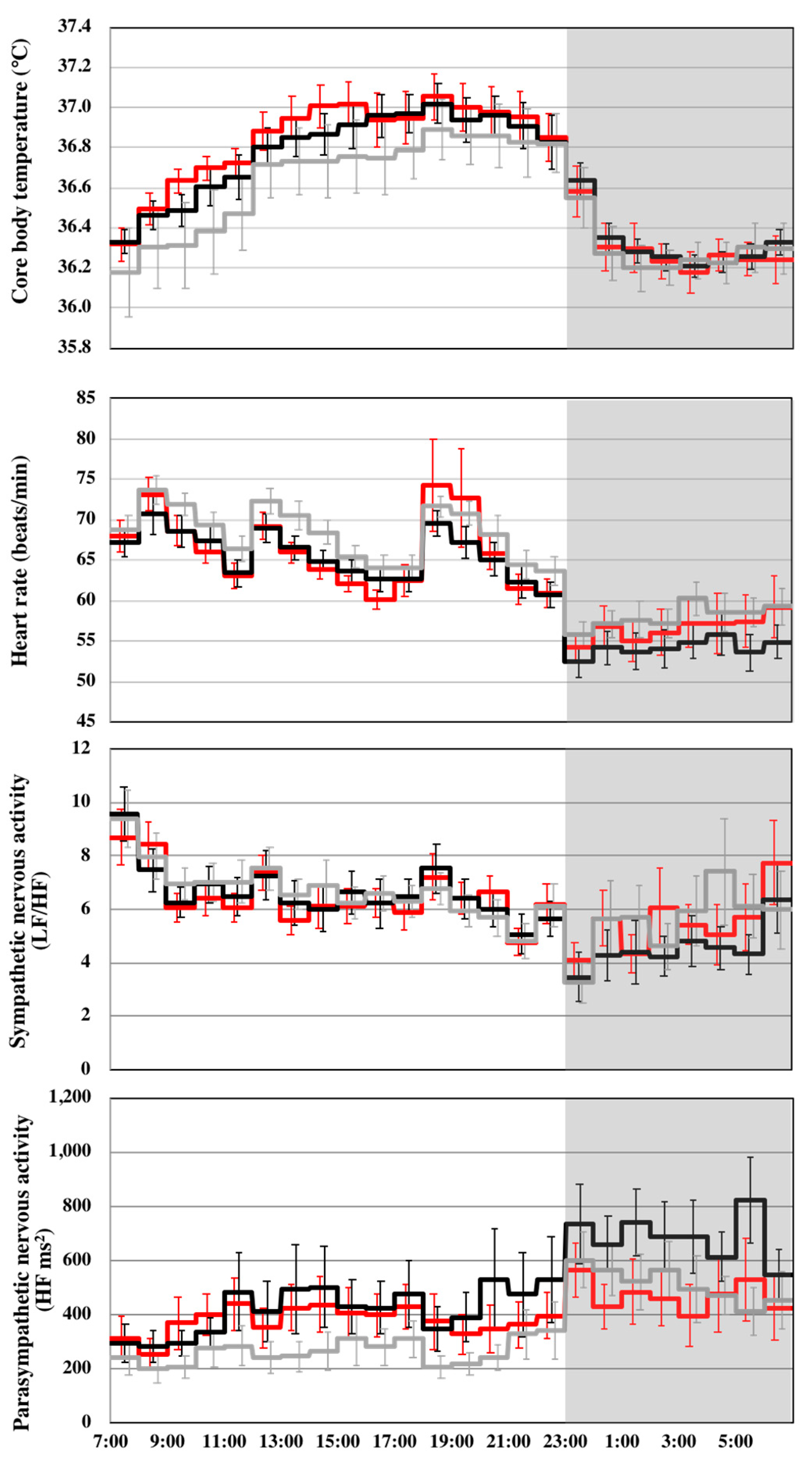
| Core body temperature (°C) | 36.5 ± 0.5 | 36.6 ± 0.4 ** | 36.7 ± 0.5 ** | 721.3 | 11.048 | p < 0.001 |
| Heart rate (beats/min) | 65 ± 8 | 62 ± 8 ** | 64 ± 10 †† | 599.2 | 10.217 | p < 0.001 |
| LF/HF | 6.85 ± 3.03 | 5.88 ± 2.86 * | 6.31 ± 2.79 | 599.5 | 3.396 | p = 0.34 |
| HF (ms2) | 315 ± 259 | 502 ± 394 ** | 393 ± 296 †† | 598.4 | 19.312 | p < 0.001 |
| Wake time | ||||||
| Energy expenditure (kcal/min) | 1.31 ± 0.20 | 1.30 ± 0.21 | 1.30 ± 0.17 | 484.0 | 0.903 | p = 0.406 |
| RQ | 0.91 ± 0.05 | 0.90 ± 0.04 ** | 0.90 ± 0.04 ** | 484.0 | 16.950 | p < 0.001 |
| Fat oxidation (kcal/min) | 0.27 ± 0.20 | 0.35 ± 0.20 ** | 0.33 ± 0.18 ** | 484.0 | 21.930 | p < 0.001 |
| Carbohydrate oxidation (kcal/min) | 0.89 ± 0.24 | 0.82 ± 0.22 ** | 0.82 ± 0.20 ** | 484.0 | 12.820 | p < 0.001 |
| Core body temperature (°C) | 36.6 ± 0.6 | 36.8 ± 0.4 ** | 36.8 ± 0.4 ** | 481.0 | 12.329 | p < 0.001 |
| Heart rate (beats/min) | 68 ± 6 | 66 ± 6 ** | 67 ± 9 | 395.6 | 5.083 | p = 0.007 |
| LF/HF | 7.21 ± 2.41 | 6.49 ± 2.52 | 6.58 ± 2.37 | 394.5 | 3.239 | p = 0.040 |
| HF (ms2) | 252 ± 201 | 427 ± 372 ** | 366 ± 266 ** | 393.7 | 21.153 | p < 0.001 |
| Sleep | ||||||
| Energy expenditure (kcal/min) | 0.91 ± 0.13 | 0.89 ± 0.14 | 0.91 ± 0.11 | 253.0 | 2.312 | p = 0.101 |
| RQ | 0.86 ± 0.05 | 0.85 ± 0.05 ** | 0.84 ± 0.04 **,† | 253.0 | 15.420 | p < 0.001 |
| Fat oxidation (kcal/min) | 0.32 ± 0.20 | 0.36 ± 0.20 * | 0.39 ± 0.15 ** | 253.0 | 12.391 | p < 0.001 |
| Carbohydrate oxidation (kcal/min) | 0.45 ± 0.13 | 0.41 ± 0.13 ** | 0.39 ± 0.12 ** | 253.0 | 11.093 | p < 0.001 |
| Core body temperature (°C) | 36.3 ± 0.4 | 36.3 ± 0.3 | 36.3 ± 0.4 | 230.8 | 0.413 | p = 0.662 |
| Heart rate (beats/min) | 58 ± 6 | 54 ± 7 ** | 56 ± 9 †† | 194.1 | 9.963 | p < 0.001 |
| LF/HF | 5.59 ± 3.95 | 4.58 ± 3.01 | 5.28 ± 3.44 | 196.3 | 3.664 | p = 0.027 |
| HF (ms2) | 511 ± 290 | 684 ± 380 * | 472 ± 346 †† | 200.1 | 8.800 | p < 0.001 |
| Placebo | Caffeine | Oolong Tea | |
|---|---|---|---|
| Time in bed (min) | 480.0 | 480.0 | 480.0 |
| Sleep period (min) | 468.6 ± 3.8 | 464.6 ± 5.8 | 460.0 ± 6.8 |
| Total sleep time (min) | 419.4 ± 11.4 | 424.9 ± 11.1 | 411.6 ± 14.9 |
| Sleep efficiency (%) | 87.4 ± 2.4 | 88.6 ± 2.3 | 85.7 ± 3.1 |
| Sleep latency (min) | 11.0 ± 3.8 | 6.1 ± 1.7 | 12.5 ± 6.2 |
| Wake after sleep onset (min) | 49.6 ± 12.1 | 48.5 ± 11.4 | 56.0 ± 15.9 |
| REM latency * (min) | 92.4 ± 7.6 | 84.7 ± 11.5 | 107.8 ± 15.5 |
| N1 (min) | 73 ± 9.8 | 68.8 ± 10.6 | 56.7 ± 7.3 |
| N2 (min) | 199.3 ± 9.7 | 216.0 ± 12.1 | 200.1 ± 10.0 |
| N3 (min) | 61.3 ± 13.9 | 55.5 ± 14.6 | 69.5 ± 15.7 |
| R (min) | 85.8 ± 5.4 | 84.6 ± 3.2 | 85.4 ± 7.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, S.; Takano, J.; Murayama, N.; Tominaga, M.; Abe, T.; Park, I.; Seol, J.; Ishihara, A.; Tanaka, Y.; Yajima, K.; et al. Subacute Ingestion of Caffeine and Oolong Tea Increases Fat Oxidation without Affecting Energy Expenditure and Sleep Architecture: A Randomized, Placebo-Controlled, Double-Blinded Cross-Over Trial. Nutrients 2020, 12, 3671. https://doi.org/10.3390/nu12123671
Zhang S, Takano J, Murayama N, Tominaga M, Abe T, Park I, Seol J, Ishihara A, Tanaka Y, Yajima K, et al. Subacute Ingestion of Caffeine and Oolong Tea Increases Fat Oxidation without Affecting Energy Expenditure and Sleep Architecture: A Randomized, Placebo-Controlled, Double-Blinded Cross-Over Trial. Nutrients. 2020; 12(12):3671. https://doi.org/10.3390/nu12123671
Chicago/Turabian StyleZhang, Simeng, Jiro Takano, Norihito Murayama, Morie Tominaga, Takashi Abe, Insung Park, Jaehoon Seol, Asuka Ishihara, Yoshiaki Tanaka, Katsuhiko Yajima, and et al. 2020. "Subacute Ingestion of Caffeine and Oolong Tea Increases Fat Oxidation without Affecting Energy Expenditure and Sleep Architecture: A Randomized, Placebo-Controlled, Double-Blinded Cross-Over Trial" Nutrients 12, no. 12: 3671. https://doi.org/10.3390/nu12123671
APA StyleZhang, S., Takano, J., Murayama, N., Tominaga, M., Abe, T., Park, I., Seol, J., Ishihara, A., Tanaka, Y., Yajima, K., Suzuki, Y., Suzuki, C., Fukusumi, S., Yanagisawa, M., Kokubo, T., & Tokuyama, K. (2020). Subacute Ingestion of Caffeine and Oolong Tea Increases Fat Oxidation without Affecting Energy Expenditure and Sleep Architecture: A Randomized, Placebo-Controlled, Double-Blinded Cross-Over Trial. Nutrients, 12(12), 3671. https://doi.org/10.3390/nu12123671