Anti-Melanogenesis Effects of Lotus Seedpod In Vitro and In Vivo

 , , and
, , and 
Abstract

1. Introduction
2. Materials and Methods
2.1. Chemical and Reagents
2.2. Lotus Seedpod Extract (LSE) Preparation
2.3. Cell Culture
2.4. Cell Viability Assay
2.5. Tyrosinase Activity Assay
2.6. Melanin Content Assay
2.7. Cyclic AMP Assay
2.8. Real-Time PCR for Analyzing mRNA Levels
2.9. Electrophoretic Mobility Shift Assay (EMSA)
2.10. Animal and Experimental Design
2.11. Western Blotting
2.12. Statistical Analysis
3. Results
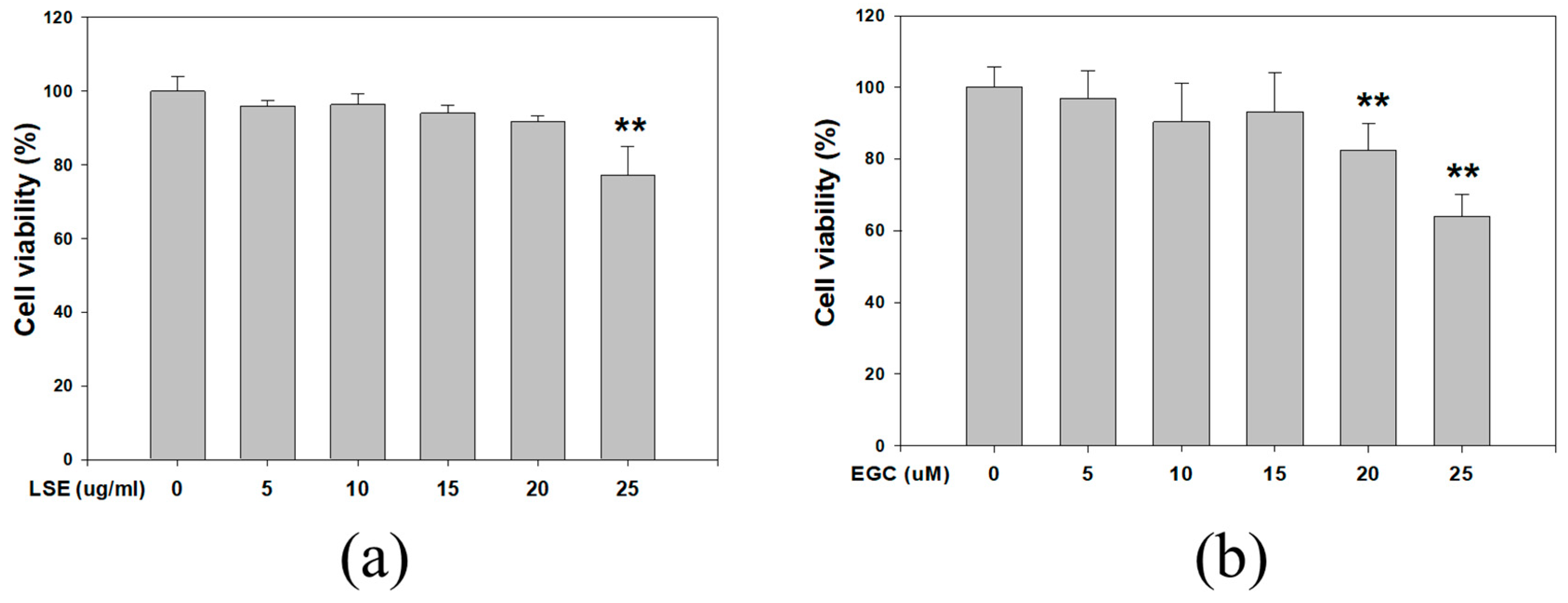
3.1. Effect of LSE and EGC on Cell Viability in B16F0 Cells
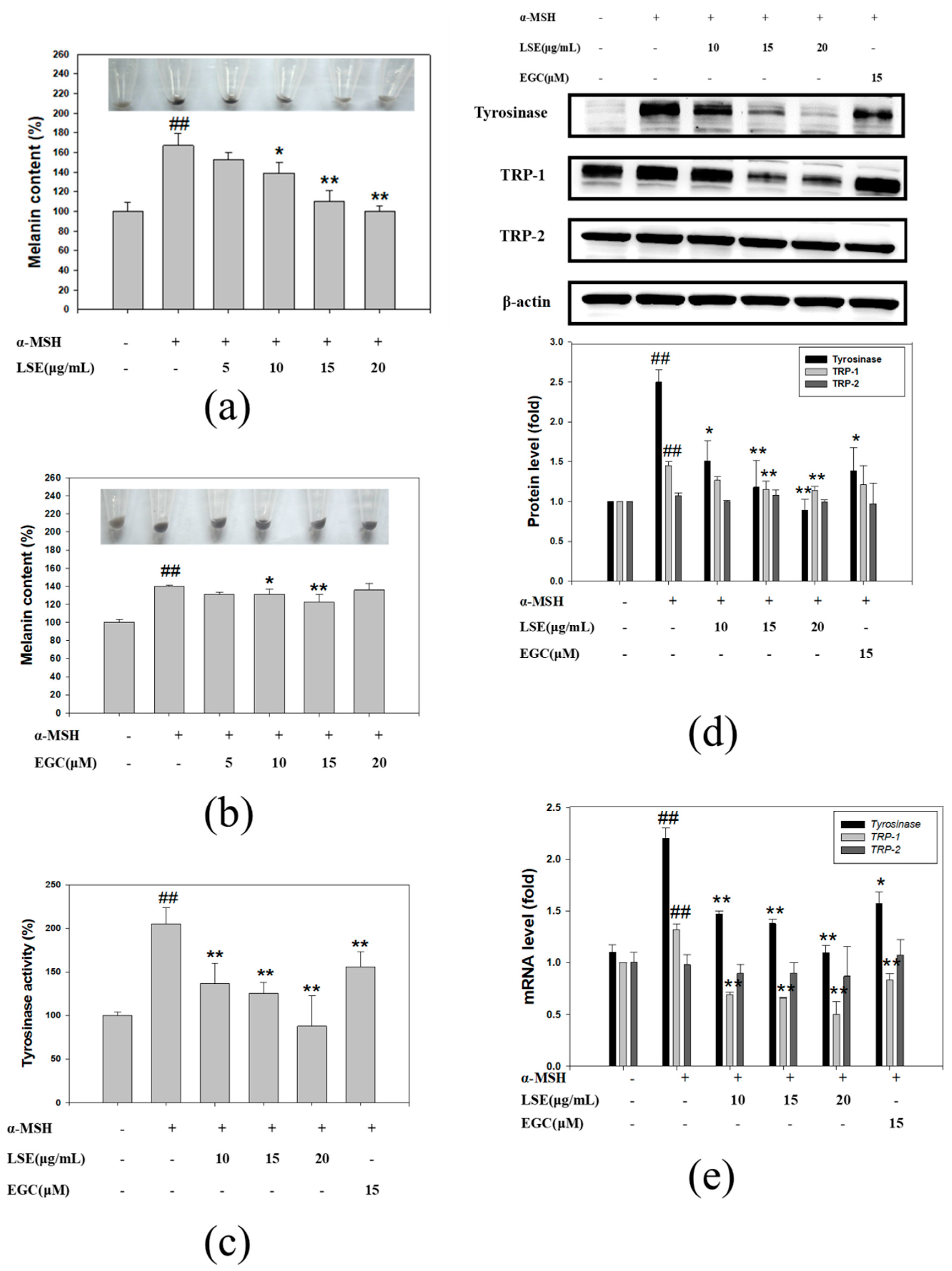
3.2. Effect of LSE and EGC on Melanin Formation and Tyrosinase Activity in α-MSH-Induced B16F0 Cells
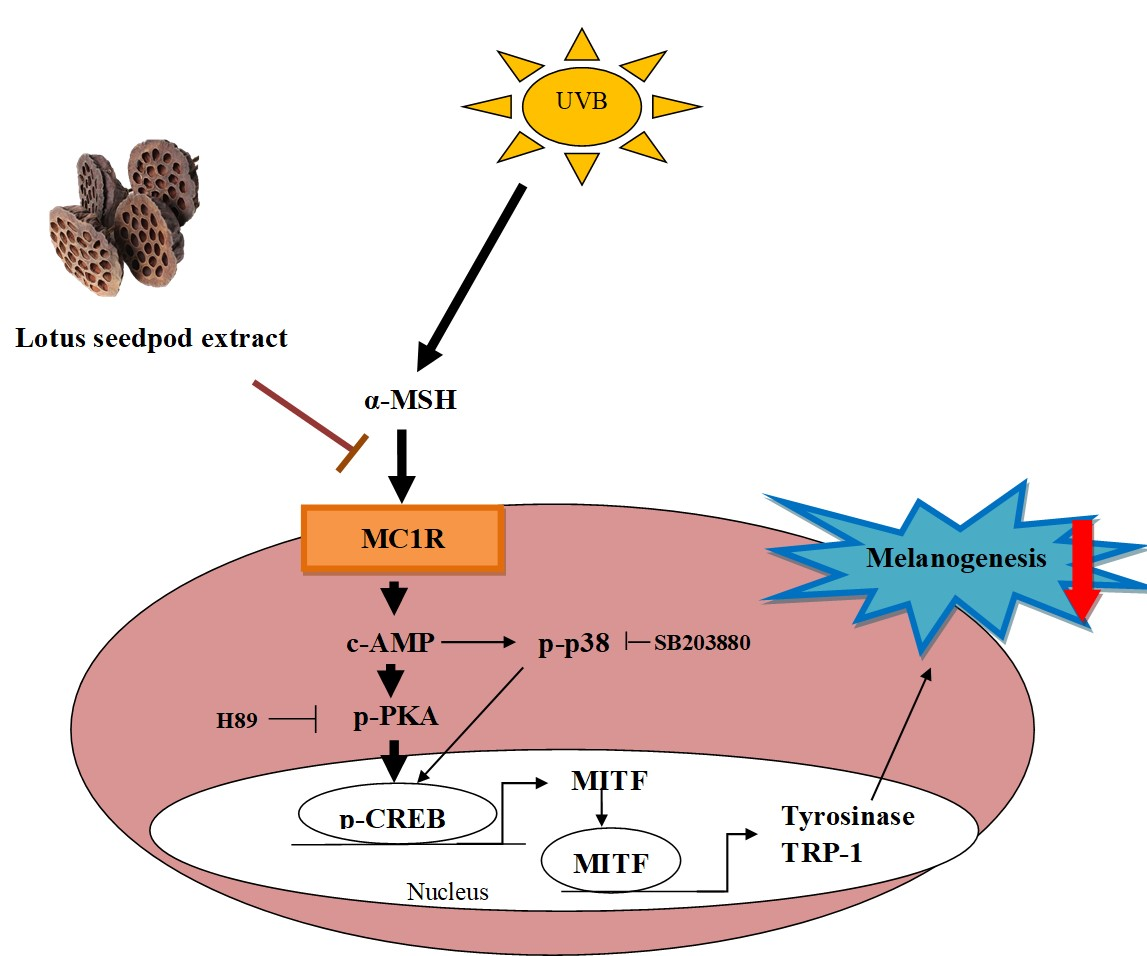
3.3. Effect of LSE and EGC on cAMP/PKA Signaling Pathway and MAPK Pathway in α-MSH-Induced B16F0 Cells
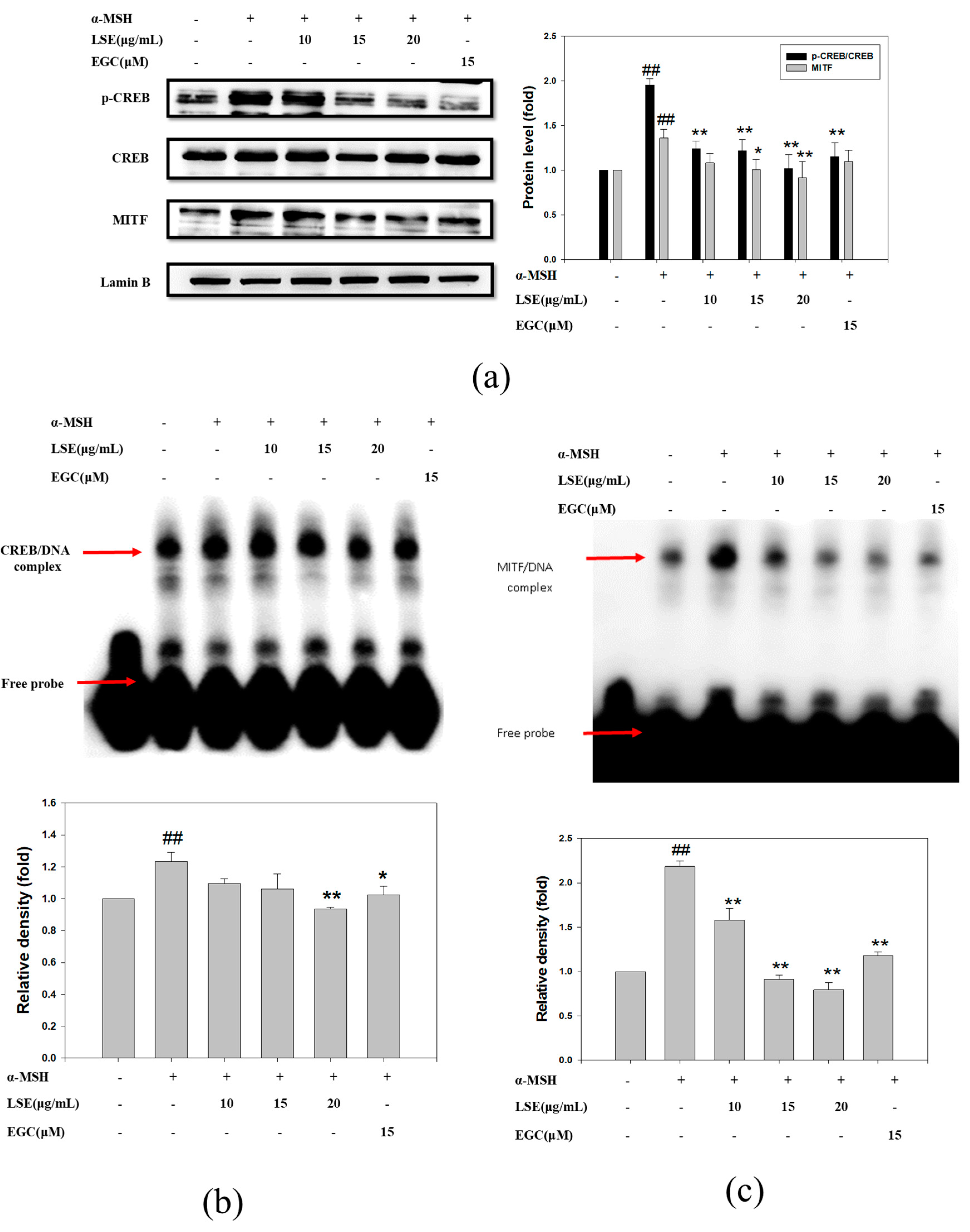
3.4. Effect of LSE and EGC on MITF and CREB in α-MSH-Induced B16F0 Cells
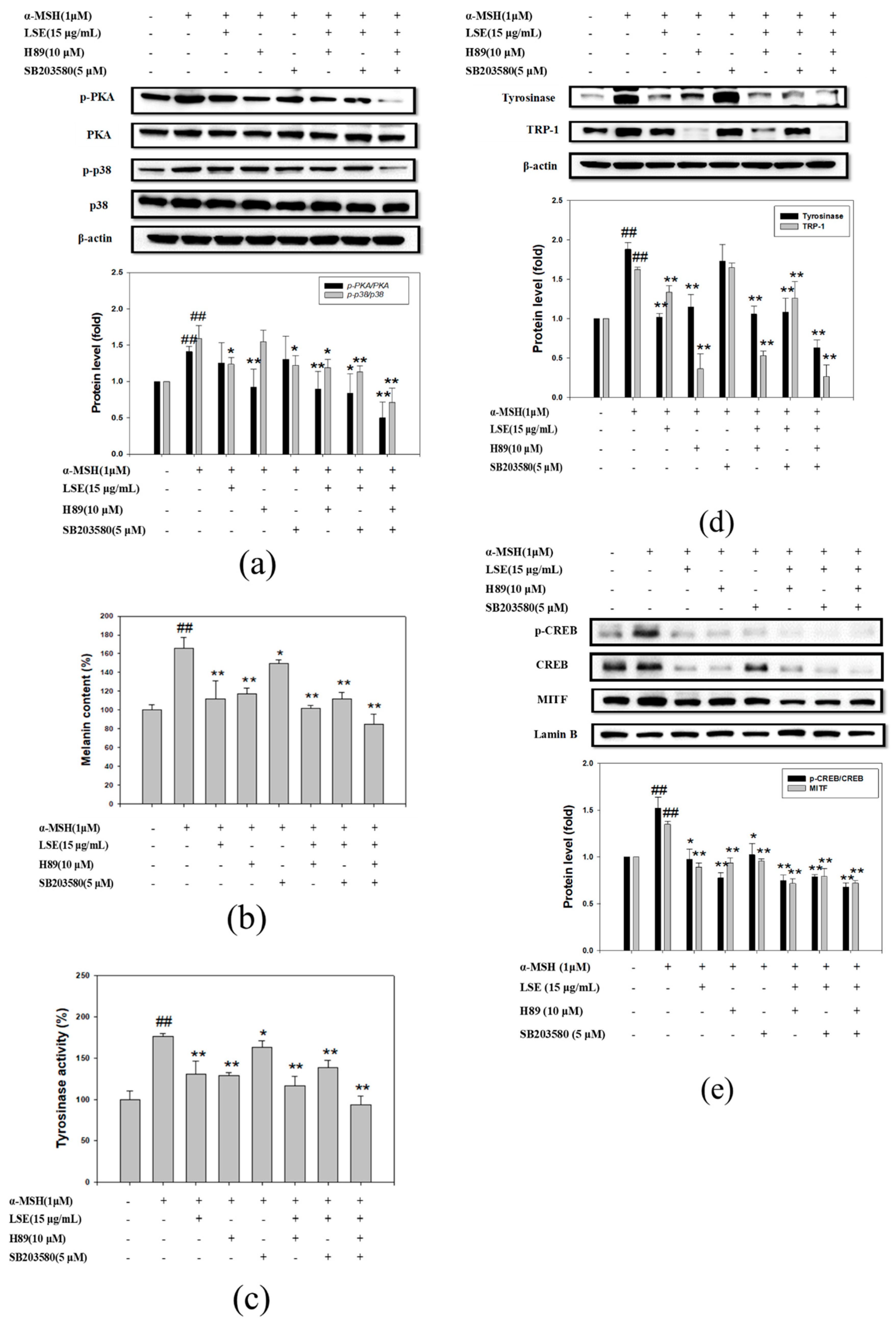
3.5. LSE Repressed Melanogenesis through Inhibiting PKA and p38 Signling Pathways
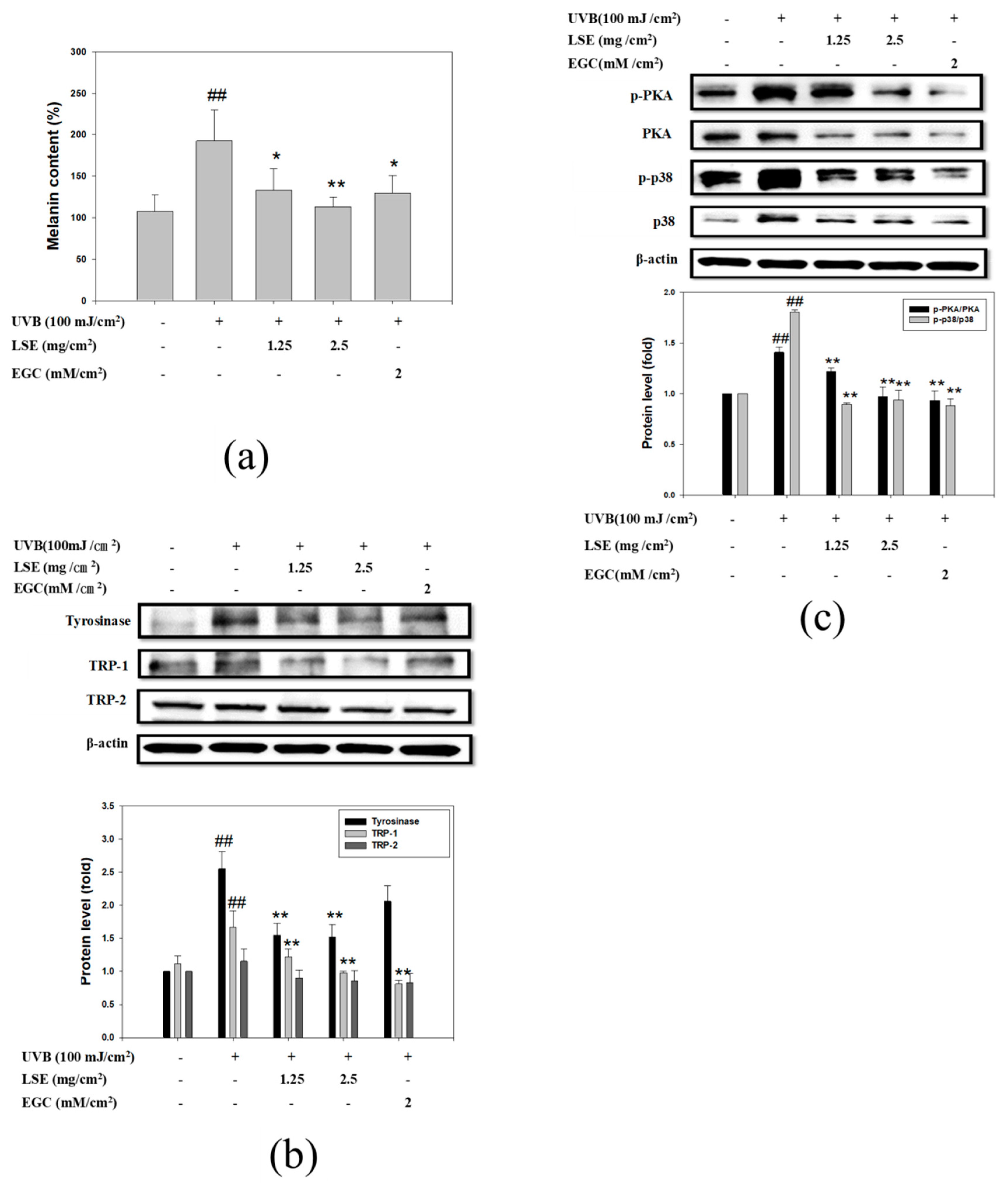
3.6. LSE and EGC Suppressed Melanin Synthesis Induced by UVB-Irridiation in Mice
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Yamaguchi, Y.; Hearing, V.J. Physiological factors that regulate skin pigmentation. Biofactors 2009, 35, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.Y.; Fisher, D.E. Melanocyte biology and skin pigmentation. Nature 2007, 445, 843–850. [Google Scholar] [CrossRef] [PubMed]
- Park, H.Y.; Kosmadaki, M.; Yaar, M.; Gilchrest, B.A. Cellular mechanisms regulating human melanogenesis. Cell. Mol. Life Sci. 2009, 66, 1493–1506. [Google Scholar] [CrossRef]
- Vance, K.W.; Goding, C.R. The transcription network regulating melanocyte development and melanoma. Pigment. Cell Res. 2004, 17, 318–325. [Google Scholar] [CrossRef]
- Kawakami, A.; Fisher, D.E. The master role of microphthalmia-associated transcription factor in melanocyte and melanoma biology. Lab. Investig. 2017, 97, 649–656. [Google Scholar] [CrossRef]
- D’Mello, S.A.N.; Finlay, G.J.; Baguley, B.C.; Askarian-Amiri, M.E. Signaling Pathways in Melanogenesis. Int. J. Mol. Sci. 2016, 17, 1144. [Google Scholar] [CrossRef]
- Wu, L.C.; Lin, Y.Y.; Yang, S.Y.; Weng, Y.T.; Tsai, Y.T. Antimelanogenic effect of c-phycocyanin through modulation of tyrosinase expression by upregulation of ERK and downregulation of p38 MAPK signaling pathways. J. Biomed. Sci. 2011, 18, 74. [Google Scholar] [CrossRef]
- Ling, Z.Q.; Xie, B.J.; Yang, E.L. Isolation, characterization, and determination of antioxidative activity of oligomeric procyanidins from the seedpod of Nelumbo nucifera Gaertn. J. Agric. Food Chem. 2005, 53, 2441–2445. [Google Scholar] [CrossRef]
- Duan, Y.; Zhang, H.; Xu, F.; Xie, B.; Yang, X.; Wang, Y.; Yan, Y. Inhibition effect of procyanidins from lotus seedpod on mouse B16 melanoma in vivo and in vitro. Food Chem. 2010, 122, 84–91. [Google Scholar] [CrossRef]
- Duan, Y.; Wang, Z.; Zhang, H.; He, Y.; Fan, R.; Cheng, Y.; Sun, G.; Sun, X. Extremely low frequency electromagnetic field exposure causes cognitive impairment associated with alteration of the glutamate level, MAPK pathway activation and decreased CREB phosphorylation in mice hippocampus: Reversal by procyanidins extracted from the lotus seedpod. Food Funct. 2014, 5, 2289–2297. [Google Scholar] [CrossRef]
- Wu, Q.; Li, S.; Li, X.; Sui, Y.; Yang, Y.; Dong, L.; Xie, B.; Sun, Z. Inhibition of Advanced Glycation Endproduct Formation by Lotus Seedpod Oligomeric Procyanidins through RAGE-MAPK Signaling and NF-kappaB Activation in High-Fat-Diet Rats. J. Agric. Food Chem. 2015, 63, 6989–6998. [Google Scholar] [CrossRef] [PubMed]
- Tseng, H.C.; Tsai, P.M.; Chou, Y.H.; Lee, Y.C.; Lin, H.H.; Chen, J.H. In Vitro and In Vivo Protective Effects of Flavonoid-Enriched Lotus Seedpod Extract on Lipopolysaccharide-Induced Hepatic Inflammation. Am. J. Chin. Med. 2019, 47, 153–176. [Google Scholar] [CrossRef]
- Liu, Y.T.; Lai, Y.H.; Lin, H.H.; Chen, J.H. Lotus Seedpod Extracts Reduced Lipid Accumulation and Lipotoxicity in Hepatocytes. Nutrients 2019, 11, 2895. [Google Scholar] [CrossRef]
- Xiao, J.S.; Xie, B.J.; Cao, Y.P.; Wu, H.; Sun, Z.D.; Xiao, D. Characterization of oligomeric procyanidins and identification of quercetin glucuronide from lotus (Nelumbo nucifera Gaertn.) seedpod. J. Agric. Food Chem. 2012, 60, 2825–2829. [Google Scholar] [CrossRef] [PubMed]
- Sato, K.; Toriyama, M. Depigmenting effect of catechins. Molecules 2009, 14, 4425–4432. [Google Scholar] [CrossRef]
- Shen, T.; Heo, S.I.; Wang, M.H. Involvement of the p38 MAPK and ERK signaling pathway in the anti-melanogenic effect of methyl 3,5-dicaffeoyl quinate in B16F10 mouse melanoma cells. Chem. Biol. Interact. 2012, 199, 106–111. [Google Scholar] [CrossRef] [PubMed]
- Bizzozero, L.; Cazzato, D.; Cervia, D.; Assi, E.; Simbari, F.; Pagni, F.; De Palma, C.; Monno, A.; Verdelli, C.; Querini, P.R.; et al. Acid sphingomyelinase determines melanoma progression and metastatic behaviour via the microphtalmia-associated transcription factor signalling pathway. Cell Death Differ. 2014, 21, 507–520. [Google Scholar] [CrossRef]
- Smalley, K.S.; Eisen, T.G. Differentiation of human melanoma cells through p38 MAP kinase is associated with decreased retinoblastoma protein phosphorylation and cell cycle arrest. Melanoma Res. 2002, 12, 187–192. [Google Scholar] [CrossRef]
- Park, S.Y.; Jin, M.L.; Kim, Y.H.; Kim, Y.; Lee, S.J. Aromatic-turmerone inhibits alpha-MSH and IBMX-induced melanogenesis by inactivating CREB and MITF signaling pathways. Arch. Dermatol. Res. 2011, 303, 737–744. [Google Scholar] [CrossRef]
- Mas, J.S.; Gerritsen, I.; Hahmann, C.; Jimenez-Cervantes, C.; Garcia-Borron, J.C. Rate limiting factors in melanocortin 1 receptor signalling through the cAMP pathway. Pigment. Cell Res. 2003, 16, 540–547. [Google Scholar] [CrossRef]
- Videira, I.F.; Moura, D.F.; Magina, S. Mechanisms regulating melanogenesis. An. Bras. Dermatol. 2013, 88, 76–83. [Google Scholar] [CrossRef] [PubMed]
- Del Marmol, V.; Beermann, F. Tyrosinase and related proteins in mammalian pigmentation. FEBS Lett. 1996, 381, 165–168. [Google Scholar] [CrossRef]
- Busca, R.; Ballotti, R. Cyclic AMP a key messenger in the regulation of skin pigmentation. Pigment. Cell Res. 2000, 13, 60–69. [Google Scholar] [CrossRef] [PubMed]
- Smalley, K.; Eisen, T. The involvement of p38 mitogen-activated protein kinase in the alpha-melanocyte stimulating hormone (alpha-MSH)-induced melanogenic and anti-proliferative effects in B16 murine melanoma cells. FEBS Lett. 2000, 476, 198–202. [Google Scholar] [CrossRef]
- Levy, C.; Khaled, M.; Fisher, D.E. MITF: Master regulator of melanocyte development and melanoma oncogene. Trends Mol. Med. 2006, 12, 406–414. [Google Scholar] [CrossRef] [PubMed]
- Abdel-Malek, Z.; Swope, V.B.; Suzuki, I.; Akcali, C.; Harriger, M.D.; Boyce, S.T.; Urabe, K.; Hearing, V.J. Mitogenic and melanogenic stimulation of normal human melanocytes by melanotropic peptides. Proc. Natl. Acad. Sci. USA 1995, 92, 1789–1793. [Google Scholar] [CrossRef]
- Maranduca, M.A.; Branisteanu, D.; Serban, D.N.; Branisteanu, D.C.; Stoleriu, G.; Manolache, N.; Serban, I.L. Synthesis and physiological implications of melanic pigments. Oncol Lett. 2019, 17, 4183–4187. [Google Scholar] [CrossRef]
- Sturm, R.A.; O’Sullivan, B.J.; Box, N.F.; Smith, A.G.; Smit, S.E.; Puttick, E.R.; Parsons, P.G.; Dunn, I.S. Chromosomal structure of the human TYRP1 and TYRP2 loci and comparison of the tyrosinase-related protein gene family. Genomics 1995, 29, 24–34. [Google Scholar] [CrossRef]
- Bolognia, J.; Murray, M.; Pawelek, J. UVB-induced melanogenesis may be mediated through the MSH-receptor system. J. Investig. Dermatol. 1989, 92, 651–656. [Google Scholar] [CrossRef]
- Lei, T.C.; Virador, V.M.; Vieira, W.D.; Hearing, V.J. A melanocyte-keratinocyte coculture model to assess regulators of pigmentation in vitro. Anal. Biochem. 2002, 305, 260–268. [Google Scholar] [CrossRef]
- Liu, S.H.; Chu, I.M.; Pan, I.H. Effects of hydroxybenzyl alcohols on melanogenesis in melanocyte-keratinocyte co-culture and monolayer culture of melanocytes. J. Enzym. Inhib. Med. Chem. 2008, 23, 526–534. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.L.; Wakamatsu, K.; Ito, S.; D’Orazio, J.A. Purification and growth of melanocortin 1 receptor (Mc1r)- defective primary murine melanocytes is dependent on stem cell factor (SFC) from keratinocyte-conditioned media. Vitr. Cell. Dev. Biol. Anim. 2009, 45, 577–583. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Bellei, B.; Flori, E.; Izzo, E.; Maresca, V.; Picardo, M. GSK3beta inhibition promotes melanogenesis in mouse B16 melanoma cells and normal human melanocytes. Cell. Signal. 2008, 20, 1750–1761. [Google Scholar] [CrossRef] [PubMed]
- Hwang, Y.S.; Oh, S.W.; Park, S.H.; Lee, J.; Yoo, J.A.; Kwon, K.; Park, S.J.; Kim, J.; Yu, E.; Cho, J.Y.; et al. Melanogenic Effects of Maclurin Are Mediated through the Activation of cAMP/PKA/CREB and p38 MAPK/CREB Signaling Pathways. Oxidative Med. Cell Longev. 2019, 2019, 9827519. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequence |
|---|---|
| Tyrosinase | Forward: 5′-CACCTGAGGGACCACTATTACG-3′ |
| Reverse: 5′-GGCAGTTCTATCCATTGATCCAG-3′ | |
| Trp-1 | Forward: 5′-ATGAAATCTTACAACGTCCTCCC-3′ |
| Reverse: 5′-TGGCACACTCTCGTGGAAACTGA-3′ | |
| Trp-2 | Forward: 5′-CTTCAACCGGACATGCAAATGC-3′ |
| Reverse: 5′-GCTTCTTCCGATTACAGTCGGG-3′ | |
| Mc1r | Forward: 5′-CTATGCGCTGCGTTATCACAG-3′ |
| Reverse: 5′-AAAGAAAGTGACGAGGCAGAG-3′ | |
| Gapdh | Forward: 5′-AGGTCGGTGTGAACGGATTTG-3′ |
| Reverse: 5′-GGGGTCGTTGATGGCAACAAT-3′ |
| Gene | Sequence |
|---|---|
| CREB | Forward: AGAGA TTGCC TGACG TCAGA GAGGT AG-5′ Biotin |
| Reverse: CTAGC TCTCT GACGT CAGGC AATCT CT-5′ Biotin | |
| MITF | Forward: ACTAC AACAC GTGTA GGCCA-5′ Biotin |
| Reverse: TGGCC TACAC GTGTT GTAGT 5′-Biotin |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, J.-Y.; Lin, H.-H.; Li, T.-S.; Tseng, C.-Y.; Wong, Y.; Chen, J.-H. Anti-Melanogenesis Effects of Lotus Seedpod In Vitro and In Vivo. Nutrients 2020, 12, 3535. https://doi.org/10.3390/nu12113535
Hsu J-Y, Lin H-H, Li T-S, Tseng C-Y, Wong Y, Chen J-H. Anti-Melanogenesis Effects of Lotus Seedpod In Vitro and In Vivo. Nutrients. 2020; 12(11):3535. https://doi.org/10.3390/nu12113535
Chicago/Turabian StyleHsu, Jen-Ying, Hui-Hsuan Lin, Ting-Shuan Li, Chaio-Yun Tseng, Yueching Wong, and Jing-Hsien Chen. 2020. "Anti-Melanogenesis Effects of Lotus Seedpod In Vitro and In Vivo" Nutrients 12, no. 11: 3535. https://doi.org/10.3390/nu12113535
APA StyleHsu, J.-Y., Lin, H.-H., Li, T.-S., Tseng, C.-Y., Wong, Y., & Chen, J.-H. (2020). Anti-Melanogenesis Effects of Lotus Seedpod In Vitro and In Vivo. Nutrients, 12(11), 3535. https://doi.org/10.3390/nu12113535