Significant Decreased Expressions of CaN, VEGF, SLC39A6 and SFRP1 in MDA-MB-231 Xenograft Breast Tumor Mice Treated with Moringa oleifera Leaves and Seed Residue (MOLSr) Extracts



Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of Moringa Mixtures
2.2. Phytochemical Screening, Total Flavonoid, Phenolic Content and Antioxidant Assays
2.3. Metabolite Profiling
2.4. Cell Culture Growth Conditions
2.5. Cell Viability Assessment
2.6. Xenograft Mice Model
2.7. In Vivo Tumorigenicity Study
2.8. Protein and Gene Expression Analysis
2.9. Gene Function and Gene Network Analysis
2.10. Statistical Analysis
3. Results
3.1. M1S9 Extracts Detected the Presence of Saponins, Amino Acids and High Glucosinolate Content but with Low Antioxidant Activity
3.2. M1S9 Extracts Exhibited the Cytotoxic Effect on MCF-7 Breast Cancer Cell Line
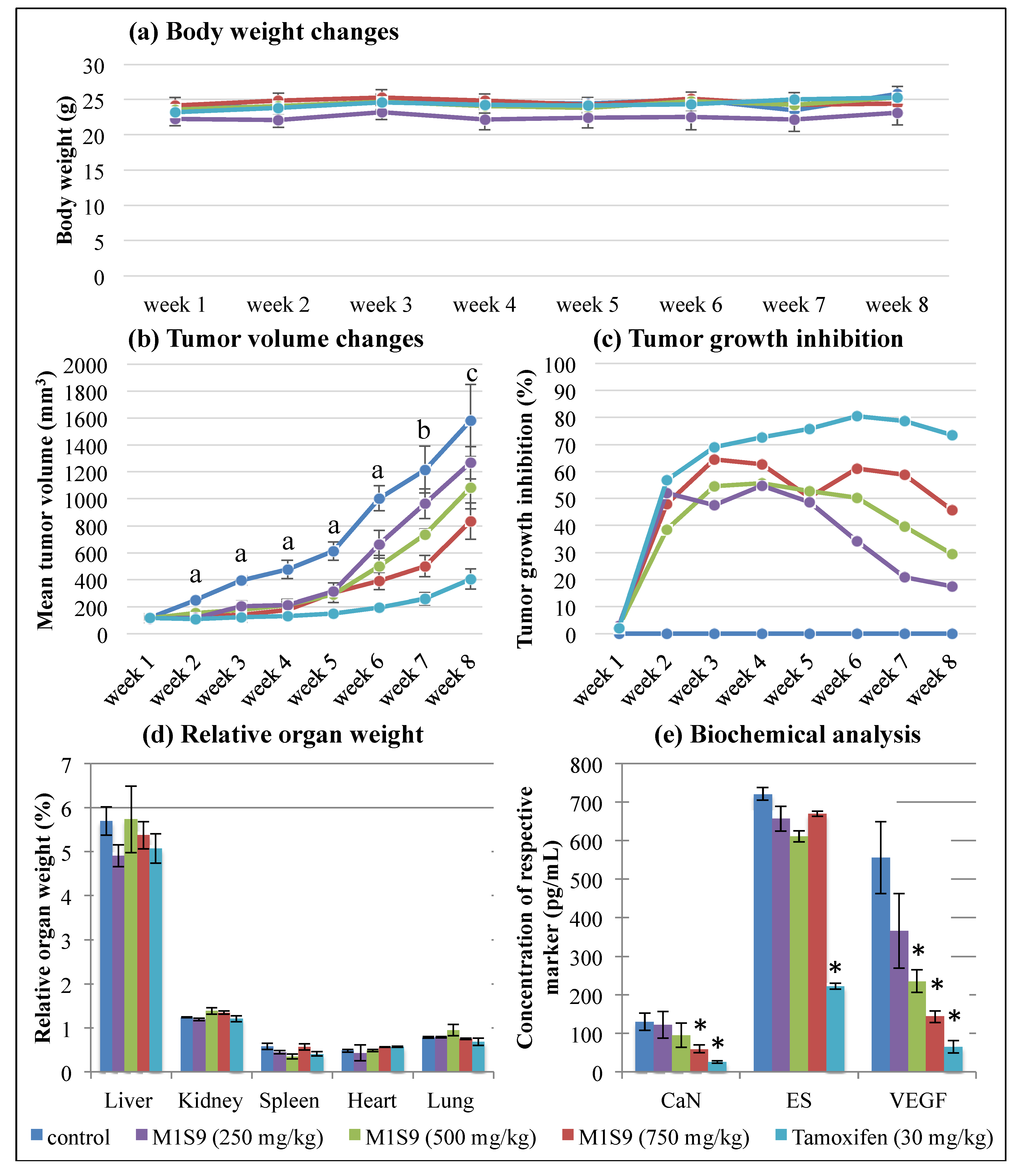
3.3. High-Dose M1S9 Extracts Suppressed Tumor Growth in MDA-MB-231 Xenograft Tumor
3.4. High-Dose M1S9 Extracts Downregulated the Expression of CaN and VEGF Proteins as Well as SFRP1 and SLC39A6 Genes in MDA-MB-231 Xenograft Tumor
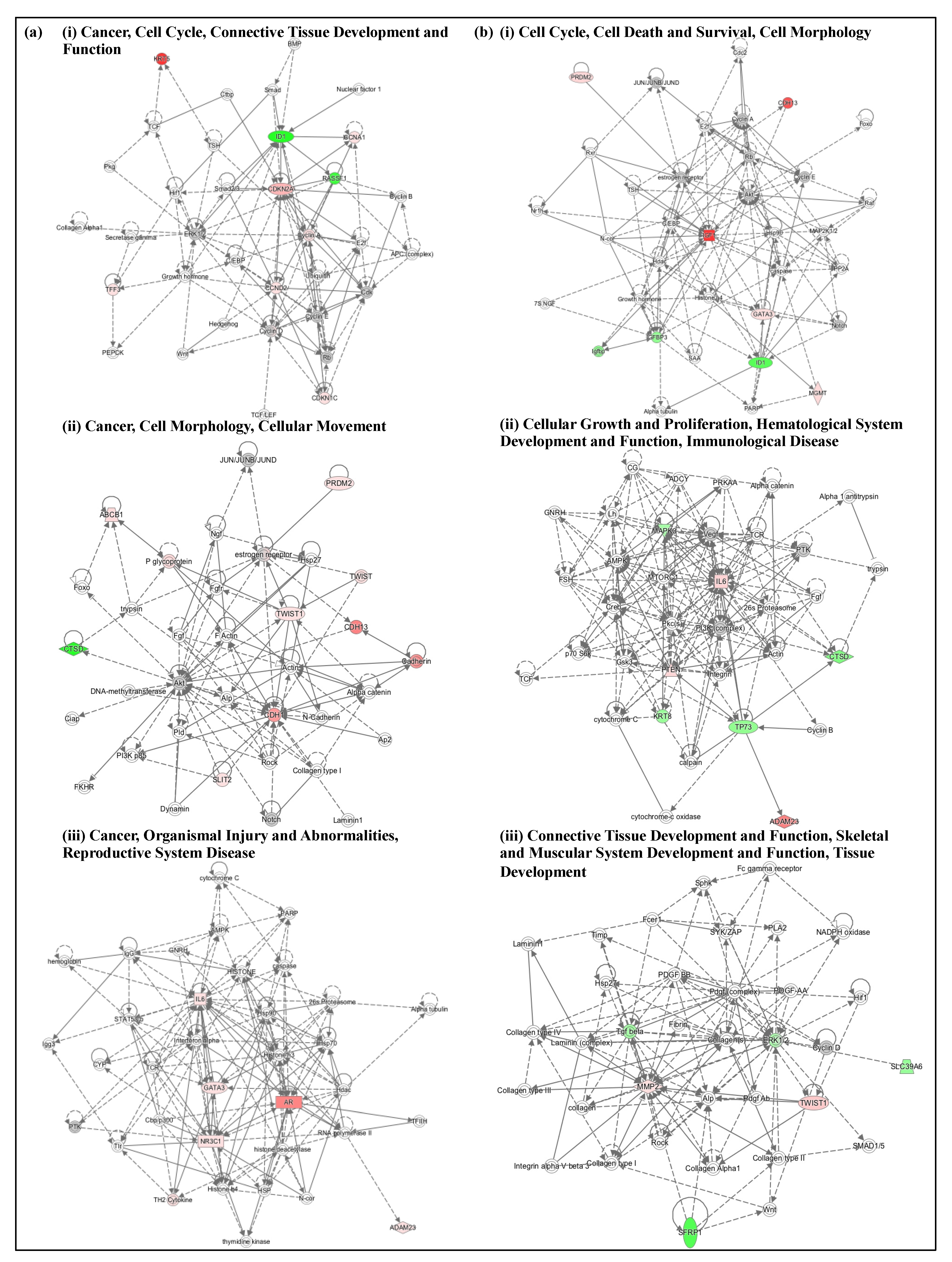
3.5. High-Dose M1S9 Regulated Genes Enriched in Specific Biological Processes, Pathways and Gene Networks
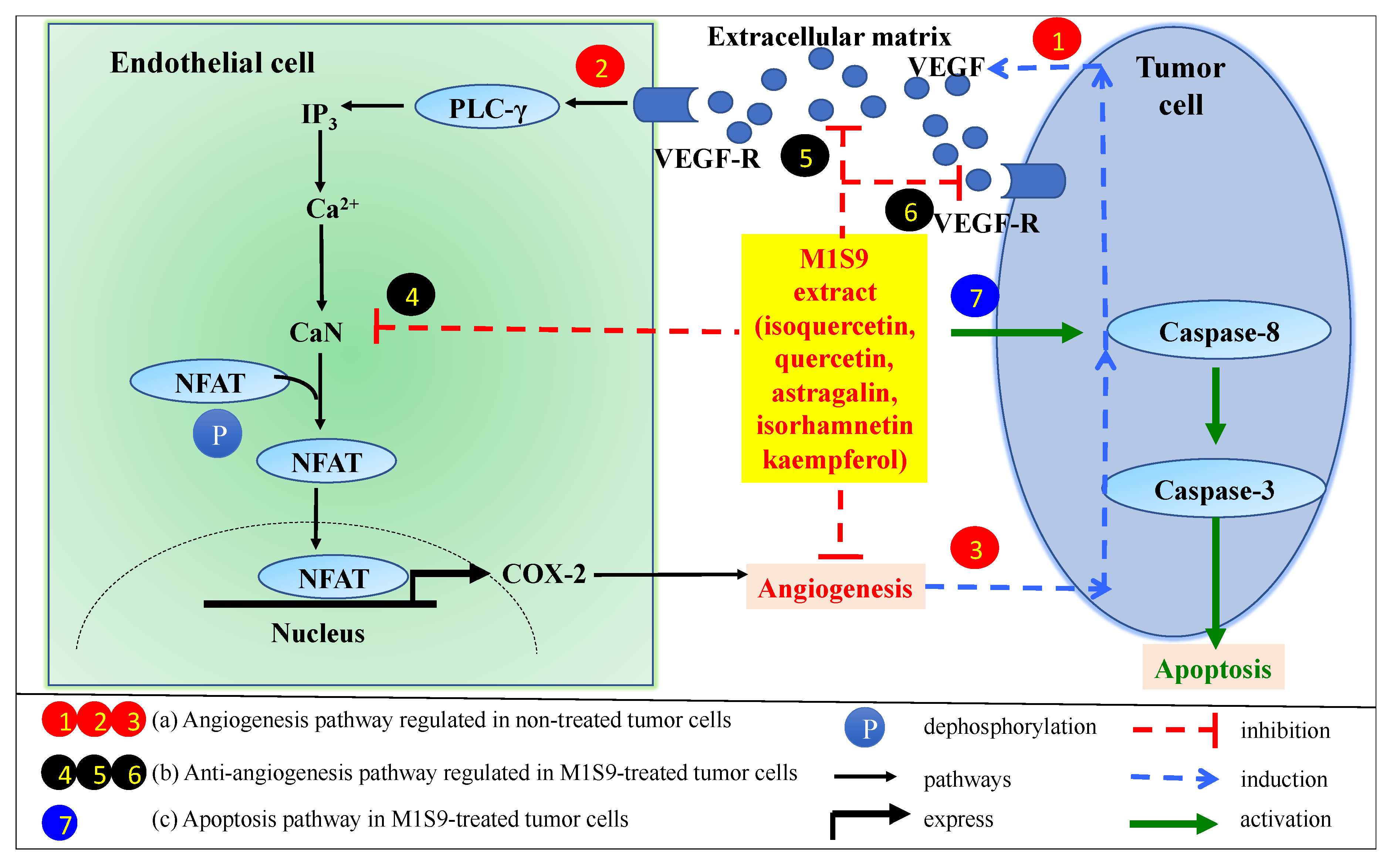
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cancer. Available online: https://www.who.int/news-room/fact-sheets/detail/cancer (accessed on 9 July 2020).
- Housman, G.; Byler, S.; Heerboth, S.; Lapinska, K.; Longacre, M.; Snyder, N.; Sarkar, S. Drug resistance in cancer: An overview. Cancers 2014, 6, 1769–1792. [Google Scholar] [CrossRef] [PubMed]
- Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [Google Scholar] [CrossRef]
- Sporn, M.B.; Dunlop, N.M.; Newton, D.L.; Smith, J.M. Prevention of chemical carcinogenesis by vitamin A and its synthetic analogs (retinoids). Fed. Proc. 1976, 35, 1332–1338. [Google Scholar] [PubMed]
- Hong, W.K.; Sporn, M.B. Recent advances in chemoprevention of cancer. Science 1997, 278, 1073–1077. [Google Scholar] [CrossRef]
- Aggarwal, B.B.; Takada, Y.; Oommen, O.V. From chemoprevention to chemotherapy: Common targets and common goals. Expert Opin. Investig. Drugs 2004, 13, 1327–1338. [Google Scholar] [CrossRef] [PubMed]
- Karim, N.A.; Ibrahim, M.D.; Kntayya, S.B.; Rukayadi, Y.; Hamid, H.A.; Razis, A.F. Moringa oleifera Lam: Targeting chemoprevention. Asian Pac. J. Cancer Prev. 2016, 17, 3675–3686. [Google Scholar]
- Kou, X.; Li, B.; Olayanju, J.B.; Drake, J.M.; Chen, N. Nutraceutical or pharmacological potential of Moringa oleifera Lam. Nutrients 2018, 10, 343. [Google Scholar] [CrossRef]
- Al-Asmari, A.K.; Albalawi, S.M.; Athar, M.T.; Khan, A.Q.; Al-Shahrani, H.; Islam, M. Moringa oleifera as an anti-cancer agent against breast and colorectal cancer cell Lines. PLoS ONE 2015, 10. [Google Scholar] [CrossRef]
- Abd-Rabou, A.A.; Abdalla, A.M.; Ali, N.A.; Zoheir, K.M. Moringa oleifera root induces cancer apoptosis more effectively than leave nanocomposites and its free counterpart. Asian Pac. J. Cancer Prev. 2017, 18, 2141–2149. [Google Scholar] [CrossRef]
- Adebayo, I.A.; Arsad, H.; Samian, M.R. Antiproliferative effect on breast cancer (Mcf7) of Moringa oleifera seed extracts. Afr. J. Tradit. Complement. Altern. Med. 2017, 14, 282–287. [Google Scholar] [CrossRef]
- Paikra, B.K.; Dhongade, H.K.J.; Gidwani, B. Phytochemistry and pharmacology of Moringa oleifera Lam. J. Pharmacopunct. 2017, 20, 194–200. [Google Scholar] [CrossRef]
- Jaja-Chimedza, A.; Graf, B.L.; Simmler, C.; Kim, Y.; Kuhn, P.; Pauli, G.F.; Raskin, I. Biochemical characterization and anti-inflammatory properties of an isothiocyanate-enriched moringa (Moringa oleifera) seed extract. PLoS ONE 2017, 12, e0182658. [Google Scholar] [CrossRef] [PubMed]
- Fahey, J.W.; Olson, M.K.; Stephenson, K.K.; Wade, K.L.; Chodur, G.M.; Odee, D.; Nouman, W.; Massiah, M.; Alt, J.; Egner, P.A.; et al. The diversity of chemoprotective glucosinolates in Moringaceae (Moringa spp.). Sci. Rep. 2018, 8. [Google Scholar] [CrossRef] [PubMed]
- Förster, N.; Ulrichs, C.; Schreiner, M.; Muller, C.T.; Mewis, I. Development of a reliable extraction and quantification method for glucosinolates in Moringa oleifera. Food Chem. 2015, 166, 456–464. [Google Scholar] [CrossRef]
- Brunelli, D.; Tavecchio, M.; Falcioni, C.; Frapolli, R.; Erba, E.; Iori, R.; Rollin, P.; Barillari, J.; Manzotti, C.; Morazzoni, P.; et al. The isothiocyanate produced from glucomoringin inhibits NF-kB and reduces myeloma growth in nude mice In Vivo. Biochem. Pharmacol. 2010, 79, 1141–1148. [Google Scholar] [CrossRef]
- Anwar, F.; Bhanger, M.I. Analytical characterization of Moringa oleifera seed oil grown in temperate regions of Pakistan. J. Agric. Food Chem. 2003, 51, 6558–6563. [Google Scholar] [CrossRef]
- Leone, A.; Spada, A.; Battezzati, A.; Schiraldi, A.; Aristil, J.; Bertoli, S. Moringa oleifera Seeds and Oil: Characteristics and Uses for Human Health. Int. J. Mol. Sci. 2016, 17, 2141. [Google Scholar] [CrossRef]
- Amante, B.; López-Grimau, V.; Smith, T. Valuation of oil extraction residue from Moringa oleifera seeds for water purification in Burkina Faso. Desalin. Water Treat. 2015, 57, 2743–2749. [Google Scholar] [CrossRef]
- Eman, N.A.; Tan, C.S.; Makky, E.A. Impact of Moringa oleifera cake residue application on waste water treatment: A case study. J. Water Res. Prot. 2014, 6, 677–687. [Google Scholar] [CrossRef][Green Version]
- Eman, N.A.; Hong, T. Heavy metals (Fe, Cu, and Cr) removal from wastewater by Moringa oleifera press cake. MATEC Web Conf. 2018, 150, 02008. [Google Scholar] [CrossRef][Green Version]
- Sreelatha, S.; Jeyachitra, A.; Padma, P.R. Antiproliferation and induction of apoptosis by Moringa oleifera leaf extract on human cancer cells. Food Chem. Toxicol. 2011, 49, 1270–1275. [Google Scholar] [CrossRef]
- Jung, I.L. Soluble extraction from Moringa oleifera leaves with a new anticancer activity. PLoS ONE 2014, 9, e95492. [Google Scholar] [CrossRef]
- Madi, N.; Dany, M.; Abdoun, S.; Usta, J. Moringa oleifera’s nutritious aqueous leaf extract has anticancerous effects by compromising mitochondrial viability in an ROS-dependent manner. J. Am. Coll. Nutr. 2016, 35, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Rahaman, M.H.A.; Kadir, N.H.A.; Amin, N.M.; Omar, W.B.W. Cytotoxicity effect of water extracts of Moringa oleifera leaves and seeds against MCF-7 cells. Acta Hortic. 2017, 1, 279–286. [Google Scholar] [CrossRef]
- Vongsak, B.; Sithisarn, P.; Mangmool, S.; Thongpraditchote, S.; Wongkrajang, Y.; Gritsanapan, W. Maximizing total phenolics, total flavonoids contents and antioxidant activity of Moringa oleifera Leaf extract by the appropriate extraction method. Ind. Crop. Prod. 2013, 44, 566–571. [Google Scholar] [CrossRef]
- Harborne, J.B. Phytochemical Methods: A Guide to Modern Techniques of Plant Analysis, 3rd ed.; Chapman and Hall: New York, NY, USA, 1998. [Google Scholar]
- Kokate, C.K. Practical Pharmacognosy, 5th ed.; Vallabh Prakashan: New Delhi, India, 2005. [Google Scholar]
- Yang, H.; Dong, Y.; Du, H.; Shi, H.; Peng, Y.; Li, X. Antioxidant compounds from propolis collected in Anhui, China. Molecules 2011, 16, 3444–3455. [Google Scholar] [CrossRef] [PubMed]
- Hisam, E.E.A.; Rofiee, M.S.; Khalid, A.M.; Jalaluddin, A.F.; Yusof, M.I.M.; Idris, M.H.; Ramli, S.; James, R.J.; Yoeng, W.J.; Lay Kek, T.; et al. Combined extract of Moringa oleifera and Centella asiatica modulates oxidative stress and senescence in hydrogen peroxide-induced human dermal fibroblasts. Turk. J. Biol. 2018, 42, 33–44. [Google Scholar] [CrossRef]
- Yoneya, T.; Taniguchi, K.; Nakamura, R.; Tsunenari, T.; Ohizumi, I.; Kanbe, Y.; Morikawa, K.; Kaiho, S.-I.; Yamada-Okabe, H. Thiochroman derivative CH4986399, A new nonsteroidal estrogen receptor down-regulator, is effective in breast cancer models. Anticancer Res. 2010, 30, 873–878. [Google Scholar]
- Saini, R.K.; Sivanesan, I.; Keum, Y.S. Phytochemicals of Moringa oleifera: A review of their nutritional, therapeutic and industrial significance. 3 Biotech. 2016, 6, 203. [Google Scholar] [CrossRef]
- Iorns, E.; Drews-Elger, K.; Ward, T.M.; Dean, S.; Clarke, J.; Berry, D.; Ashry, D.E.; Lippman, M. A new mouse model for the study of human breast cancer metastasis. PLoS ONE 2012, 7, e47995. [Google Scholar] [CrossRef]
- Peters, J.M.; Boyd, E.M. Organ weights and water levels of the rat following reduced food intake. J. Nutr. 1966, 90, 354–360. [Google Scholar] [CrossRef] [PubMed]
- Michael, B.; Yano, B.; Sellers, R.S.; Perry, R.; Morton, D.; Roome, N.; Johnson, J.K.; Schafer, K. Evaluation of organ weights for rodent and non-rodent toxicity studies: A review of regulatory guidelines and a survey of current practices. Toxicol. Pathol. 2007, 35, 742–750. [Google Scholar] [CrossRef]
- Zong, J.; Wang, R.; Bao, G.; Ling, T.; Zhang, L.; Zhang, X.; Hou, R. Novel triterpenoid saponins from residual seed cake of Camellia oleifera Abel. show anti-proliferative activity against tumor cells. Fitoterapia 2015, 104, 7–13. [Google Scholar] [CrossRef]
- Anhwange, B.A.; Ajibola, V.O.; Oniye, S.J. Amino acid composition of the seeds of Moringa oleifera (Lam), Detarium microcarpum (Guill & Sperr) and Bauhinia monandra (Linn.). ChemClass J. 2004, 1, 9–13. [Google Scholar]
- Jahan, I.A.; Hossain, M.H.; Ahmed, K.S.; Sultana, Z.; Biswas, P.K.; Nada, K. Antioxidant activity of Moringa oleifera seed extracts. Orient. Pharm. Exp. Med. 2018, 18, 299–307. [Google Scholar] [CrossRef]
- Geran, R.I.; Greenberg, N.H.; Macdonald, M.M.; Schumacher, A.M.; Abbott, B.J. Protocols for screening chemical agents and natural products against animal tumors and other biological systems. Cancer Chemother. Rep. 1972, 3, 1–102. [Google Scholar]
- Araújo, L.C.; Aguiar, J.S.; Napoleao, T.H.; Mota, F.V.; Barros, A.L.; Moura, M.C.; Coriolano, M.C.; Coelho, L.C.; Silva, T.G.; Paiva, P.M. Evaluation of cytotoxic and anti-inflammatory activities of extracts and lectins from Moringa oleifera seeds. PLoS ONE 2013, 8, e81973. [Google Scholar] [CrossRef] [PubMed]
- Ragasa, C.Y.; Levida, R.M.; Don, M.-J.; Shen, C.-C. Cytotoxic isothiocyanates from Moringa oleifera Lam seeds. Philipp. Sci. Lett. 2012, 5, 46–52. [Google Scholar]
- Khalil, W.K.B.; Ghaly, I.S.; Diab, K.A.E.; ELmakawy, A.I. Antitumor activity of Moringa oleifera leaf extract against Ehrlich solid tumor. Int. J. Pharm. 2014, 4, 68–82. [Google Scholar]
- Ferrara, N. Vascular endothelial growth factor a target for anticancer therapy. Oncologist 2004, 9, 2–10. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, Q.; Yang, S.; Chen, C.; Li, X.; Liu, J.; Zou, Z.; Cai, D. Quercetin inhibits angiogenesis by targeting calcineurin in the xenograft model of human breast cancer. Eur. J. Pharmacol. 2016, 781, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Taylor, K.M.; Nicholson, R.I. The LZT proteins; the LIV-1 subfamily of zinc transporters. Biochim. Biophys. Acta 2003, 1611, 16–30. [Google Scholar] [CrossRef]
- Taylor, K.M.; Morgan, H.E.; Smart, K.; Zahari, N.M.; Pumford, S.; Ellis, I.O.; Robertson, J.F.; Nicholson, R.I. The emerging role of the LIV-1 subfamily of zinc transporters in breast cancer. Mol. Med. 2007, 13, 396–406. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Wei, L.; Huang, X.; Zheng, J.; Shao, M.; Feng, T.; Li, J.; Han, Y.; Tan, W.; Tan, W.; et al. Solute carrier family 39 member 6 gene promotes aggressiveness of esophageal carcinoma cells by increasing intracellular levels of zinc, activating phosphatidylinositol 3-kinase signaling, and up-regulating genes that regulate metastasis. Gastroenterology 2017, 152, 1985–1997. [Google Scholar] [CrossRef]
- Surana, R.; Sikka, S.; Cai, W.; Shin, E.M.; Warrier, S.R.; Tan, H.J.; Arfuso, F.; Fox, S.A.; Dharmarajan, A.M.; Kumar, A.P. Secreted frizzled related proteins: Implications in cancers. Biochim. Biophys. Acta 2014, 1845, 53–65. [Google Scholar] [CrossRef]
- Matsuda, Y.; Schlange, T.; Oakeley, E.J.; Boulay, A.; Hynes, N.E. WNT signaling enhances breast cancer cell motility and blockade of the WNT pathway by sFRP1 suppresses MDA-MB-231 xenograft growth. Breast Cancer Res. 2009, 11, R32. [Google Scholar] [CrossRef]
- Martin-Manso, G.; Calzada, M.J.; Chuman, Y.; Sipes, J.M.; Xavier, C.P.; Wolf, V.; Kuznetsova, S.A.; Rubin, J.S.; Roberts, D.D. sFRP-1 binds via its netrin-related motif to the N-module of thrombospondin-1 and blocks thrombospondin-1 stimulation of MDA-MB-231 breast carcinoma cell adhesion and migration. Arch. Biochem. Biophys. 2011, 509, 147–156. [Google Scholar] [CrossRef]
- Qu, Y.; Ray, P.S.; Li, J.; Cai, Q.; Bagaria, S.P.; Moran, C.; Sim, M.S.; Zhang, J.; Turner, R.R.; Zhu, Z.; et al. High levels of secreted frizzled-related protein 1 correlate with poor prognosis and promote tumourigenesis in gastric cancer. Eur. J. Cancer 2013, 49, 3718–3728. [Google Scholar] [CrossRef]
- Barba, F.J.; Nikmaram, N.; Roohinejad, S.; Khelfa, A.; Zhu, Z.; Koubaa, M. Bioavailability of glucosinolates and their breakdown products: Impact of processing. Front. Nutr. 2016, 3, 24. [Google Scholar] [CrossRef]
- Soundararajan, P.; Kim, J.S. Anti-carcinogenic glucosinolates in cruciferous vegetables and their antagonistic effects on prevention of cancers. Molecules 2018, 23, 2983. [Google Scholar] [CrossRef]
- Jurkowska, H.; Wróbel, M.; Szlęzak, D.; Jasek-Gajda, E. New aspects of antiproliferative activity of 4-hydroxybenzyl isothiocyanate, a natural H2S-donor. Amino Acids 2018, 50, 699–709. [Google Scholar] [CrossRef] [PubMed]
- Gong, X.; Smith, J.R.; Swanson, H.M.; Rubin, L.P. Carotenoid lutein selectively inhibits breast cancer cell growth and potentiates the effect of chemotherapeutic agents through ROS-mediated mechanisms. Molecules 2018, 23, 905. [Google Scholar] [CrossRef] [PubMed]
- Hermawan, A.; Nur, K.A.; Sarmoko, D.; Putri, P.; Meiyanto, E. Ethanolic extract of Moringa oleifera increased cytotoxic effect of doxorubicin on HeLa cancer cells. J. Nat. Rem. 2012, 12, 108–114. [Google Scholar]
- Berkovich, L.; Earon, G.; Ron, I.; Rimmon, A.; Vexler, A.; Lev-Ari, S. Moringa oleifera aqueous leaf extract down-regulates nuclear factor-kappaB and increases cytotoxic effect of chemotherapy in pancreatic cancer cells. BMC Complement. Altern. Med. 2013, 13, 212. [Google Scholar] [CrossRef] [PubMed]
- Yaacob, N.S.; Kamal, N.N.; Norazmi, M.N. Synergistic anticancer effects of a bioactive subfraction of Strobilanthes crispus and tamoxifen on MCF-7 and MDA-MB-231 human breast cancer cell lines. BMC Complement. Altern. Med. 2014, 14, 252. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | MOLSr | ||||
|---|---|---|---|---|---|
| Negative | Positive | M1S9 | M1S1 | M9S1 | |
| (a) Qualitative Phytochemical Analysis | |||||
| Tannins | − | ++++ | + | +++ | +++ |
| Triterpenoids | − | ++++ | − | − | − |
| Flavonoids | − | ++++ | − | + | ++ |
| Saponins | − | +++ | ++ | − | − |
| Anthraquinone glycosides | − | ++++ | − | − | − |
| Phenols | − | ++++ | + | +++ | ++++ |
| Steroids | − | ++++ | − | − | − |
| Alkaloids | − | ++++ | − | − | − |
| Proteins | − | + | + | + | + |
| Amino acids | − | ++++ | + | − | − |
| Fats and fixed oils | − | ++++ | +++ | ++ | ++ |
| Carbohydrates | − | ++++ | +++ | ++ | ++ |
| (b) Quantitative Phytochemical Analysis | |||||
| TFC (µg QE/mg) | NR | NR | 0.88 ± 0.09 | 2.58 ± 0.11 | 4.72 ± 0.06 |
| TPC (µg GAE/mg) | NR | NR | 57.7 ± 1.03 | 68.0 ± 0.53 | 82.2 ± 4.57 |
| (c) Antioxidant Assays | |||||
| DPPH (IC50, µg/mL) | 1.2 ± 0.10 * | − | 113.43 ± 25.45 | 12.50 ± 3.67 | 11.65 ± 4.99 |
| FRAP (µg Trolox/mg) | − | 172.41 ± 12.6 $ | 1.37 ± 0.56 | 2.04 ± 0.67 | 5.60 ± 0.73 |
| Compound Group | Compound Name | Retention Time | Molecular Formula | Molecular Weight | Percentage of Relative Abundance (%) | ||||
|---|---|---|---|---|---|---|---|---|---|
| MOL | MOSr | M1S9 | M1S1 | M9S1 | |||||
| Glucosinolates | Glucosinalbin | 1.351 | C14H19NO10S2 | 425.0419 | ND | 0.107 | 0.046 | 0.029 | ND |
| Glucosinolates | Glucotropaeolin | 1.395 | C14H19NO9S2 | 409.0502 | ND | 0.016 | 0.004 | ND | ND |
| Miscellaneous | Cinnamic acid | 1.667 | C9H8O2 | 148.0516 | 0.090 | 0.020 | 0.060 | 0.060 | 0.090 |
| Miscellaneous | Quinoline | 2.501 | C9H7N | 129.0575 | 0.007 | ND | 0.004 | 0.005 | 0.008 |
| Miscellaneous | Benzoic acid | 5.913 | C7H6O2 | 122.037 | ND | 0.010 | 0.007 | ND | ND |
| Glycoside | Niazirin | 8.212 | C14H17NO5 | 279.1121 | ND | 0.009 | 0.009 | 0.006 | 0.005 |
| Flavonoid | isoquercetin | 10.422 | C21H20O12 | 464.0956 | 1.030 | ND | 0.630 | 0.930 | 1.230 |
| Flavonoid | Quercetin | 10.442 | C15H10O7 | 302.0436 | 0.560 | ND | 0.350 | 0.510 | 0.680 |
| Flavonoid | Astragalin | 11.478 | C21H20O11 | 448.1009 | 0.370 | ND | 0.240 | 0.340 | 0.510 |
| Steroid | Strophanthidin | 11.703 | C23H32O6 | 404.2181 | 0.050 | 0.01 | 0.020 | 0.030 | 0.040 |
| Flavonoid | Isorhamnetin | 11.772 | C16H12O7 | 316.0584 | 0.040 | ND | 0.020 | 0.030 | 0.040 |
| Flavonoid | Kaempferol | 12.291 | C15H10O6 | 286.0479 | 0.060 | ND | 0.020 | 0.040 | 0.060 |
| Glycoside | Niazimicin | 14.802 | C16H23NO6S | 357.1252 | 0.050 | ND | 0.030 | 0.040 | 0.050 |
| Phenolic acid | p-coumaric acid | 20.79 | C9H8O3 | 164.0475 | ND | 0.005 | 0.005 | ND | ND |
| Miscellaneous | Vanillin | 15.878 | C8H8O3 | 152.0479 | ND | 0.010 | 0.008 | ND | ND |
| Miscellaneous | Heptadecane | 26.47 | C17H36 | 240.2828 | ND | 0.003 | ND | ND | 0.002 |
| Miscellaneous | Palmitic acid | 34.17 | C16H32O2 | 256.2414 | 0.004 | ND | ND | 0.003 | 0.003 |
| Cell Lines | Time Points (h) | IC50 (μg/mL) | |||
|---|---|---|---|---|---|
| TAM | M1S9 | M1S1 | M9S1 | ||
| Primary mammary epithelial cells (PMECs) | 24 | 4.0 | 70.0 | 82.0 | >100.0 |
| 48 | 4.0 | 33.5 | 78.5 | >100.0 | |
| 72 | 3.0 | 17.5 | 51.0 | >100.0 | |
| Non-malignant Chang’s liver cells | 24 | 4.0 | >100.0 | >100.0 | >100.0 |
| 48 | 6.0 | 93.5 | >100.0 | >100.0 | |
| 72 | 3.0 | >100.0 | >100.0 | >100.0 | |
| Hepatocellular carcinoma (HepG2) | 24 | 7.0 | >100.0 | >100.0 | >100.0 |
| 48 | 4.0 | >100.0 | >100.0 | >100.0 | |
| 72 | 3.0 | >100.0 | >100.0 | >100.0 | |
| Colorectal carcinoma cells (HCT-116) | 24 | 9.0 | >100.0 | >100.0 | >100.0 |
| 48 | 5.5 | >100.0 | >100.0 | >100.0 | |
| 72 | 4.5 | >100.0 | >100.0 | >100.0 | |
| Cervical adenocarcinoma cells (HeLa) | 24 | 5.0 | 91.0 | >100.0 | >100.0 |
| 48 | 3.5 | 75.0 | >100.0 | >100.0 | |
| 72 | 3.0 | 81.0 | 91.0 | >100.0 | |
| Breast adenocarcinoma cells (MCF-7) | 24 | 15.5 | 97.5 | >100.0 | >100.0 |
| 48 | 7.5 | 53.0 | >100.0 | >100.0 | |
| 72 | 8.0 | 38.5 | 57.0 | >100.0 | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lim, W.F.; Mohamad Yusof, M.I.; Teh, L.K.; Salleh, M.Z. Significant Decreased Expressions of CaN, VEGF, SLC39A6 and SFRP1 in MDA-MB-231 Xenograft Breast Tumor Mice Treated with Moringa oleifera Leaves and Seed Residue (MOLSr) Extracts. Nutrients 2020, 12, 2993. https://doi.org/10.3390/nu12102993
Lim WF, Mohamad Yusof MI, Teh LK, Salleh MZ. Significant Decreased Expressions of CaN, VEGF, SLC39A6 and SFRP1 in MDA-MB-231 Xenograft Breast Tumor Mice Treated with Moringa oleifera Leaves and Seed Residue (MOLSr) Extracts. Nutrients. 2020; 12(10):2993. https://doi.org/10.3390/nu12102993
Chicago/Turabian StyleLim, Wai Feng, Mohd Izwan Mohamad Yusof, Lay Kek Teh, and Mohd Zaki Salleh. 2020. "Significant Decreased Expressions of CaN, VEGF, SLC39A6 and SFRP1 in MDA-MB-231 Xenograft Breast Tumor Mice Treated with Moringa oleifera Leaves and Seed Residue (MOLSr) Extracts" Nutrients 12, no. 10: 2993. https://doi.org/10.3390/nu12102993
APA StyleLim, W. F., Mohamad Yusof, M. I., Teh, L. K., & Salleh, M. Z. (2020). Significant Decreased Expressions of CaN, VEGF, SLC39A6 and SFRP1 in MDA-MB-231 Xenograft Breast Tumor Mice Treated with Moringa oleifera Leaves and Seed Residue (MOLSr) Extracts. Nutrients, 12(10), 2993. https://doi.org/10.3390/nu12102993