Citrus Flavonoids as Promising Phytochemicals Targeting Diabetes and Related Complications: A Systematic Review of In Vitro and In Vivo Studies

 ,
,  ,
,  , , , , and
, , , , and
Abstract
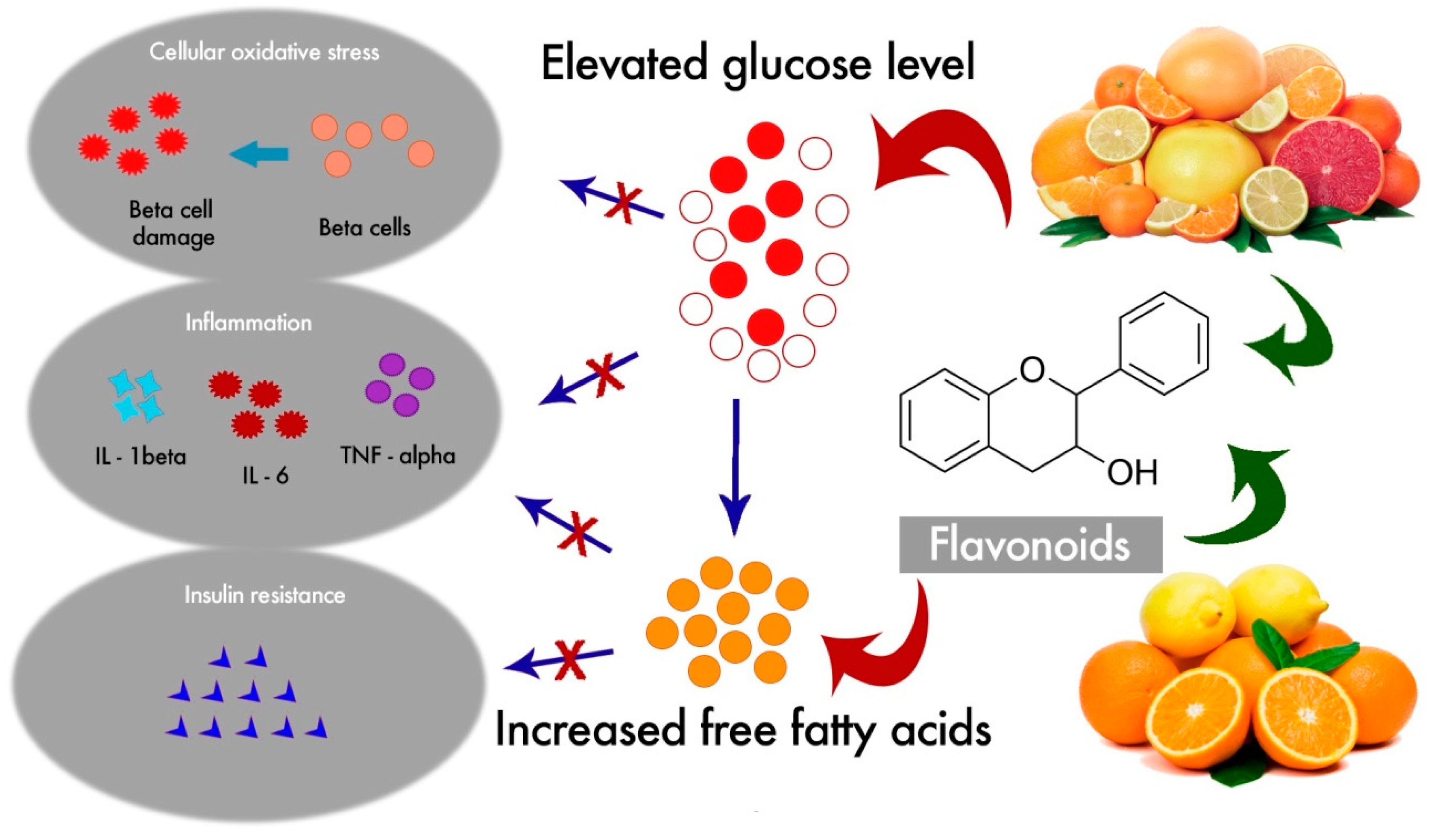
1. Introduction
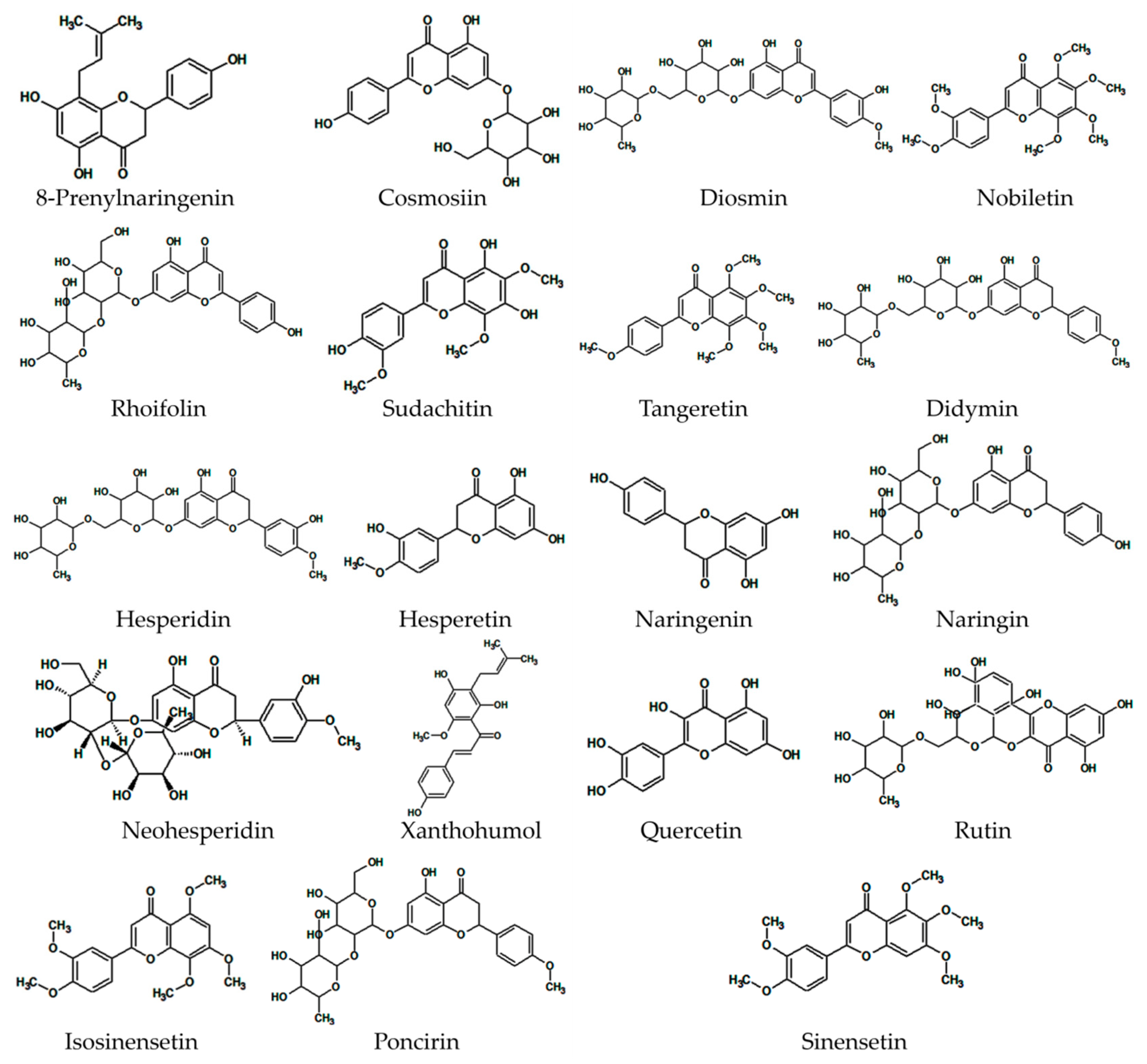
2. Main Citrus Flavonoids with Antidiabetic Effects
2.1. 8-Prenylnaringenin
2.2. Cosmosiin
2.3. Diosmin
2.4. Nobiletin
2.5. Rhoifolin

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Flavonoids | Class | Concentrations and Duration of the Treatment | In Vitro Models | Effects and Molecular Mechanisms | Ref. |
|---|---|---|---|---|---|
| Nobiletin | Flavone | 1, 2.5, 5, 10, and 20 µM; 24 h | HepG2 cells (human hepatoma cells) | Nobiletin activated mitogen-activated protein kinase-extracellular signal-related kinase (MAPK/ERK), resulting in the marked inhibition of apolipoprotein B100 secretion. It neither induced the phosphorylation of the insulin receptor (IR) or insulin receptor substrate-1(IRS-1) tyrosine nor triggered lipogenesis associated with insulin resistance. | [41] |
| Rhoifolin and cosmosiin from Citrus grandis (L.) Osbeck leaves | Flavone | Rhoifolin: 0.001–5 μM; cosmosiin: 1–20 μM; 24 h | 3T3-L1 adipocyte cells | Rhoifolin and cosmosiin exerted antidiabetic effects by promoting adiponectin secretion, the tyrosine phosphorylation of IR-β, and glucose transporter 4 (GLUT4) translocation. These bioactive molecules may help in insulin resistance-related treatment for diabetic complications. | [30] |
| Tangeretin and nobiletin | Flavone | 5–50 mM; 24 h | 3T3-F442A preadipocytes | Tangeretin and nobiletin induced increased glucose uptake in murine adipocytes, suggesting that the action was mediated by phosphatidylinositol 3-kinase (PI3K) as well as protein kinase B (Akt) and protein kinase A (PKA)/cAMP-response element-binding protein (CREB) signaling-dependent pathways. | [42] |
| Flavonoids from Citrus aurantium Linn. include naringin, esperidin, poncirin, sosiennsetin, sineesytin, nobiletin, and tangeretin | Flavone and flavanone | 0, 10, and 50 μg/mL; 0–6 days | 3T3-L1 preadipocytes | C. aurantium containing flavonoids decreased the expression of key adipocyte differentiation regulators, including CCAAT-enhancer binding protein family (C/EBPβ and C/EBPα) and peroxisome proliferator-activated receptor gamma (PPARγ); it reduced adipogenesis and the accumulation of cytoplasmic lipid droplets during differentiation in 3T3-L1 cells. | [53] |
| Nobiletin | Flavone | 0, 1, 10, and 100 μM; 7 days | 3T3-L1 preadipocytes | Nobiletin suppressed the differentiation of 3T3-L1 preadipocytes into adipocytes by down-regulating the expression of the gene coding for PPARγ2. In addition, nobiletin reduced the phosphorylation of CREB and strongly improved the phosphorylation of signal transducer and activation of transcription (STAT)5. | [43] |
| Sudachitin | Flavone | 30 mmol/L; 48 h | Primary myoblasts | Sudachitin increased mitochondrial biogenesis and improved mitochondrial function, leading to an improvement in lipid and glucose metabolism mediated via the sirtuin (Sirt) 1-AMP-activated protein kinase (AMPK)-peroxisome proliferator-activated receptor gamma coactivator-1- alpha (PGC-1α) pathway. | [54] |
| Naringenin | Flavanone | 0, 10, and 50 μM; 3 h | RAW 264 (macrophages) cells and 3T3-L1 adipocytes | Naringenin inhibited the monocyte chemoattractant protein-1 (MCP-1)’s mRNA expression and secretion in the adipocytes in a dose-dependent manner. It also prevented the MCP-1 production stimulated by the interaction between the adipocytes and the infiltrated macrophages. | [55] |
| Hesperidin and naringin | Flavanone | 0.25, 0.5, 1, and 2 mg/mL; 1 and 24 h | Pancreatic islets | Hesperidin and naringin increased the production and the release of insulin from the islet cells and decreased the intestinal glucose absorption. | [56] |
| Quercetin | Flavanol | 10 and 100 mM; 24 h | L6 myotubes | Quercetin activated the adenosine monophosphate kinase (AMPK)-P38 MAPK pathway and up-regulated glucose transporter type 4 (GLUT4)/AKT mRNA expression to induce glucose uptake in skeletal muscle cell lines. | [57] |
| Diosmin | Flavone | 0.01–1 μmol/L; 24 h | Transfected imidazoline receptor (I-R) gene in CHO-K1 cells (Chinese hamster ovary cell) | Diosmin enhanced calcium influx in I-R gene-transfected CHO-K1 cells. Diosmin effectively activated the I-R gene via inducing opioid secretion, showing utility as an antidiabetic drug. | [35] |
| Hesperidin | Flavanone | 12.5, 25, and 50 μmol/L; 6 h | RGC-5 cells (retinal ganglial cells) | Hesperidin protected against a high level of glucose-induced cell apoptosis by down-regulating caspase-9, caspase-3, and Bax/Bcl-2. Furthermore, it significantly inhibited the phosphorylation of c-Jun N-terminal kinases (JNK) and activated p38 MAPK in high glucose-fed RGC-5 cells. | [58] |
| Hesperidin and hesperetin | Flavanone | 40, 80, 120, 160, and 200 μM; 24 h | Rat liver cells | Flavonoids hesperidin and hesperetin inhibited the activities of two gluconeogenesis enzymes, alanine aminotransferase (ALT) and aspartate aminotransferase (AST), indicating their effectiveness in treating AST and ALT-mediated metabolic disorders, including in diabetes mellitus. | [59] |
| Tangeretin | Flavone | 0, 2.5, 5, and 10 μM; 24 h | Human glomerular mesangial cells (MCs) | Tangeretin very effectively inhibited high glucose (HG)-induced cell proliferation, oxidative stress, and extracellular matrix (ECM) expression in the human glomerular mesangial cells (MCs) via inactivating the extracellular signal-regulated kinase (ERK) signaling pathway. It also displayed therapeutic potential in the management of diabetic nephropathy. | [60] |
| Didymin | Flavanone | 10 and 20 μM; 6 and 24 h | Human umbilical vein endothelial cells (HUVECs) | Didymin protected against high glucose (HG)-induced human umbilical vein endothelial cells by modulating the expression of intercellular adhesion molecule (ICAM)-1 and vascular cell adhesion protein (VCAM)-1, and regulating nuclear factor kappa B (NF-κB)-mediated inflammatory cytokines and chemokines. Didymin prevented HG-induced endothelial dysfunction and death via antioxidative and anti-inflammatory activities. | [61] |
| Didymin | Flavanone | 10–30 μM; 15 and 30 min, 1 and 24 h, 28 days | HepG2 (human hepatocarcinoma) cell line | Didymin inhibited α-glucosidase, activated the insulin-signaling pathway, and improved insulin sensitivity. It showed potent inhibitory activity against the key enzymes involved in diabetes mellitus, including protein tyrosine phosphatase 1B (PTP1B), α- glucosidase, advanced glycation end products (AGEs), and aldose reductase (AR). | [62] |
| Naringenin | Flavanone | 0.01–1 μM; 1 and 24 h | NSC34 (mouse neuroblastoma and embryonic spinal cord motor neurons) cell line | Naringenin suppressed neuronal apoptosis and enhanced antioxidant protective effects in methylglyoxal (MG)-treated NSC34 cells. It prevented MG-induced hyperglycemia-related neurotoxicity via regulating insulin-like growth factor 1 receptor (IGF-1R)-mediated signaling. | [63] |
| Naringenin | Flavanone | 0, 1, 10, and 50 μM; 30 min, 3 and 6 h | 3T3-L1 (adipocytes) and RAW264 (macrophages) cells | Naringenin inhibited monocyte chemotactic protein (MCP)-3 expression in 3T3-L1 adipocytes and a coculture of 3T3-L1 adipocytes and RAW264 macrophages. It did not affect the expression of macrophage inflammatory protein-2 (MIP-2), a key chemokine for neutrophil migration and activation, in macrophages or a coculture of adipocytes and macrophages. | [64] |
| Nobiletin | Flavone | 10 μM; 1 and 4 h | HepG2 (human hepatocarcinoma) cell line | Nobiletin increased pAMPK in HepG2 incubated with high glucose content, in which the phosphorylation of AMPK was suppressed, which was comparable to the action carried out by the reference standards (resveratrol and metformin). | [46] |
2.6. Sudachitin
2.7. Tangeretin
2.8. Didymin
2.9. Hesperidin
2.10. Hesperetin
| Flavonoids | Class | Animal Models | Dose/Route/Duration of the Experiment | Effects and Molecular Mechanisms | Ref. |
|---|---|---|---|---|---|
| Hesperidin | Flavanone | Wistar Rats | Hesperidin-containing animal diet (10 g/kg diet); 28 days | Hesperidin attenuated hyperglycemia and hyperlipidemia by decreasing blood glucose and normalizing hepatic glucose-regulating enzyme activities but did not affect bone tissue and bone metabolic parameters in streptozotocin (STZ)-injected marginal diabetic weanling rats. | [82] |
| Rutin | Flavonol | Wistar Rats | 50 mg/kg (intraperitoneal); 45 days | Rutin significantly reduced the blood glucose level, improved the lipid profiles, and normalized the activities of hepatic enzymes in STZ-induced diabetic rats. It also regulated hyperglycemia and dyslipidemia, and inhibited the progression of liver and heart dysfunction in STZ-induced diabetic rats. | [99] |
| Nobiletin | Flavone | C57BL/6 Ldlr-/- Mice | Nobiletin (0.1 or 0.3% mixed in high-fat Western diet); 56 to 182 days | Nobiletin regulated liver biomarkers by increasing hepatic and peripheral insulin sensitivity, improving glucose tolerance, and protecting against the development of atherosclerosis. | [41] |
| Naringenin | Flavanone | Wistar Rats | 10 mg/kg (intraperitoneal); 35 days | Naringenin ameliorated aortic reactivity dysfunction in diabetic rats by attenuating lipid peroxidation and oxidative injury via a nitric acid-dependent pathway. | [100] |
| Hesperidin and naringin | Flavanone | Wistar Rats | 50 mg/kg (oral administration); 28 and 30 days | Hesperidin and naringin lowered the level of pro-inflammatory cytokine (tumor necrosis factor-alpha (TNF-α) and interleukin (IL)-6) production and enhanced antioxidant defenses in a type 2 diabetes rat model by normalizing the altered blood glucose and antioxidant parameters in the liver. | [83] |
| Diosmin | Flavone | Wistar Rats | 100 mg/kg (intragastric); 45 days | Diosmin attenuated lipid abnormalities in the diabetic rats via reducing the plasma and tissue lipids significantly, along with a profound increase in high-density lipoprotein cholesterol (HDL-C) levels. | [33] |
| Hesperidin | Flavanone | Wistar Rats | 25, 50, or 100 mg/kg (oral administration); 21 days | Hesperidin reduced hyperglycaemia, decreased malondialdehyde (MDA) and IL-6 levels, and enhanced the brain-derived neurotrophic factor (BDNF) and monoamines in the brain, thereby enabling it to be effective in treating and managing neurogenesis in diabetic rats. | [84] |
| Naringenin | Flavanone | Wistar Rats | 20, 50, and 100 mg/kg (oral administration); 56 days | Naringenin restored hyperglycemia, down-regulated superoxide dismutase activity, and reversed chemical and thermal hyperalgesia in the diabetic rats, showing its preventive and therapeutic effectiveness in diabetic neuropathy treatment. | [101] |
| Diosmin | Flavone | Sprague-Dawley Rats | 50 and 100 mg/kg (oral administration); 28 days | Diosmin significantly restored the blood glucose levels, antioxidant parameters, and lipid profiles in the diabetic rats. It also improved their thermal hyperalgesia, cold allodynia, and walking function. | [34] |
| Sudachitin | Flavone | C57BL/6 J and db/db Mice | 5 mg/kg (oral administration); 84 days | Sudachitin significantly improved dyslipidemia, reduced triglyceride and free fatty acid contents, enhanced glucose tolerance, and reduced insulin resistance in the diabetic mice. β-oxidation of fatty acids was also markedly enhanced via increased mitochondrial biogenesis. | [54] |
| Tangeretin | Flavone | Wistar Rats | 25, 50, and 100 mg/kg (intragastric); 30 days | Tangeretin normalized the levels and activities of plasma glucose, insulin, glycosylated hemoglobin, and key enzymes of carbohydrate metabolism in the livers of diabetic rats. | [71] |
| Hesperidin | Flavanone | Sprague-Dawley Rats | 25, 50, and 100 mg/kg (oral administration); 28 days | Hesperidin decreased the levels of STZ-induced hyperglycemia and pro-inflammatory cytokines and increased the nociceptive threshold, motor nerve conduction velocity, sensory nerve conduction velocity, insulin levels, and Na-K-adenosine triphosphate (ATP)ase activity in the diabetic rats. | [85] |
| Naringenin | Flavanone | C57BL/6J Mice | 100 mg/kg (oral administration); 14 days | Naringenin suppressed macrophage infiltration into the adipose tissues of the high-fat diet (HFD)-fed obese mice. It also down-regulated monocyte chemoattractant protein-1 (MCP-1) in the adipose tissues via inhibiting the c-Jun NH2-terminal kinase (JNK) pathway. | [55] |
| Neohesperidin | Flavanone | KK-Ay and C57BL/6 Mices | 50 mg/kg (oral administration); 42 days | Neohesperidin attenuated fasting blood glucose and insulin resistance. The levels of total cholesterol, triglycerides, and leptins were significantly decreased, while the phosphorylation of AMP-activated protein kinase (AMPK) and its target genes was increased in the drug-treated mice. It also significantly decreased the size of epididymal adipocytes in the diabetic mice. | [102] |
| Hesperidin and naringin | Flavanones | Wistar Rats | 50 mg/kg (oral administration); 30 days | Hesperidin and naringin significantly reduced the glucose level, restored the altered parameters of glucose metabolism, and enhanced adipose tissue glucose transporter type 4 (GLUT4) mRNA and protein expression in the diabetic rats. | [56] |
| Tangeretin | Flavone | Wistar Rats | 100 mg/kg (intragastric); 30 days | Tangeretin significantly reduced plasma and cardiac lipid profiles by regulating key lipid metabolic enzymes in the livers of diabetic rats. It also markedly restored the GLUT4 expression, antioxidant enzyme activities, and levels of inflammatory cytokines in the heart tissues of the the tangeretin-treated diabetic rats. | [72] |
| Nobiletin | Flavone | Wistar rats | 10 and 25 mg/kg (oral administration); 28 days | Nobiletin substantially ameliorated hemodynamic parameters, oxidative stress, collagen levels, matrix metalloproteinase (MMP)-2 levels, and MMP-9 levels in the diabetic rats. It also markedly attenuated deterioration in the morphology of cardiomyocytes. | [44] |
| Nobiletin | Flavone | C57BL/6 Mice | 50 mg/kg (oral administration); 77 days | Nobiletin significantly decreased the expression of nicotinamide adenine dinucleotide (NADH) oxidase isoforms p67phox, p22phox, and p91phox, and attenuated oxidative stress in diabetic mice. It also ameliorated the development of cardiac dysfunction and interstitial fibrosis by down-regulating the c-Jun N-terminal kinase (JNK), P38, and nuclear factor kappa B NF-κB signaling pathways. | [45] |
| Diosmin | Flavone | Sprague-Dawley Rats | 160 mg/kg (intraperitoneal); 7 days | Diosmin reduced hyperglycemia by enhancing the secretion of β-endorphin from the adrenal glands via imidazoline 1–2 receptor (I-2R) activation, which triggered the opioid receptors to attenuate gluconeogenesis metabolism in the livers of diabetic rats. It decreased the hepatic glycogen content and plasma lipid profiles in STZ-induced diabetic rats. However, it did not adversely affect the body weight, food intake, and plasma insulin level in the diabetic rats. | [35] |
| Hesperidin and quercetin | Flavanone and flavone | Wistar Rats | 100 mg/kg (oral administration); 15 days | Hesperidin and quercetin exerted positive effects on insulin metabolism. They lowered the levels of triglycerides, MDA, TNFα, and IL-6, and restored the level of glutathione (GSH) in experimental diabetic rats induced by STZ. | [87] |
| Hesperidin | Flavanone | Sprague-Dawley Rats | 25, 50, and 100 mg/kg (oral administration); 21 days | Hesperidin ameliorated the increased levels of blood glucose, serum insulin, food intake, and water intake in STZ- induced diabetes. It also had a protective effect on the wound architecture by accelerating angiogenesis and vasculogenesis via the up-regulation of vascular endothelial growth factor c (VEGF-c), Angiopoietin (Ang)-1/Tie-2, transforming growth factor (TGF-β), and small mothers against decapentaplegic (Smad)-2/3 mRNA expression to enhance wound healing in the chronic diabetic foot ulcer condition in the diabetic rats. | [86] |
| Xanthohumol and 8-prenylnaringenin | Prenylflavonoid | C57Bl/6 Mice | 0.1% of flavonoids dissolved in ethanol; 140 days | Xanthohumol and 8-prenylnaringenin have a potent therapeutic effect on diabetic mice, as evidenced by the decreased levels of diabetes-linked biochemical parameters in the liver and kidney. They also decreased the overexpression of galectin-3 (Gal3), which was correlated with oxidative stress in diabetic mice. | [27] |
| Naringin | Flavanone | Sprague-Dawley Rats | 100 mg/kg (oral administration); 28 days | Naringin reduced blood glucose, total cholesterol, triglycerides, and low-density lipoproteins in fructose-fed rats. Naringin restored acetylcholine-mediated vasorelaxation, suggesting its potential influence on fructose-induced metabolic alterations and endothelial dysfunction. Naringin improved serum nitrate/nitrite (NOx), endothelial nitric oxide synthase (eNOS), and phosphorylated eNOS (p-eNOS) protein expression, and preserved endothelium-dependent relaxation in the aortae of the fructose-fed rats. | [103] |
| Hesperetin | Flavanone | Wistar Rats | 40 mg/kg (intragastric); 45 days | Hesperetin reduced the blood glucose level and enhanced the plasma insulin and the hepatic glycogen levels in the STZ-induced diabetic rats. It also restored the altered hepatic glucose metabolic enzymes, lipid profiles, and serum biomarkers, and protected from STZ-mediated structural alterations and functional changes in the liver, kidneys, and pancreatic β-cells of diabetic animals. | [97] |
| Hesperetin | Flavanone | Wistar Rats | 50 mg/kg (oral administration); 46 days | Hesperetin significantly reduced the serum glucose level and improved the serum testosterone level in the STZ-induced diabetic rats. Additionally, it augmented the testicular antioxidant enzymes and attenuated the testicular inflammatory markers, such as TNFα and IL-17, besides preventing the seminiferous tubules’ damage in diabetic rats. | [98] |
| Naringenin | Flavanone | C57BL/6J Mice | 100 mg/kg (oral administration); 14 days | Naringenin inhibited neutrophil infiltration into the adipose tissues of the high-fat diet (HFD)-fed mice by reducing the expression of several chemokines, including monocyte chemoattractant protein (MCP)-1 and MCP-3, in the adipose tissues. | [64] |
| Nobiletin | Flavone | Ldlr-/- and Ampkβ1-/- mice from a C57BL/6J background | 0.3% of nobiletin mixed in HFD; 84–126 days | Nobiletin attenuated obesity, hepatic steatosis, dyslipidemia, and insulin resistance, and improved energy utilization in HFD-fed mice. It conferred metabolic protection independently of AMPK activation in the liver and adipose tissues. | [46] |
2.11. Naringenin
2.12. Naringin
2.13. Neohesperidin
2.14. Xanthohumol
2.15. Quercetin
2.16. Rutin
3. Composition of Antidiabetic Citrus Flavonoids in Common Citrus Fruit Sources
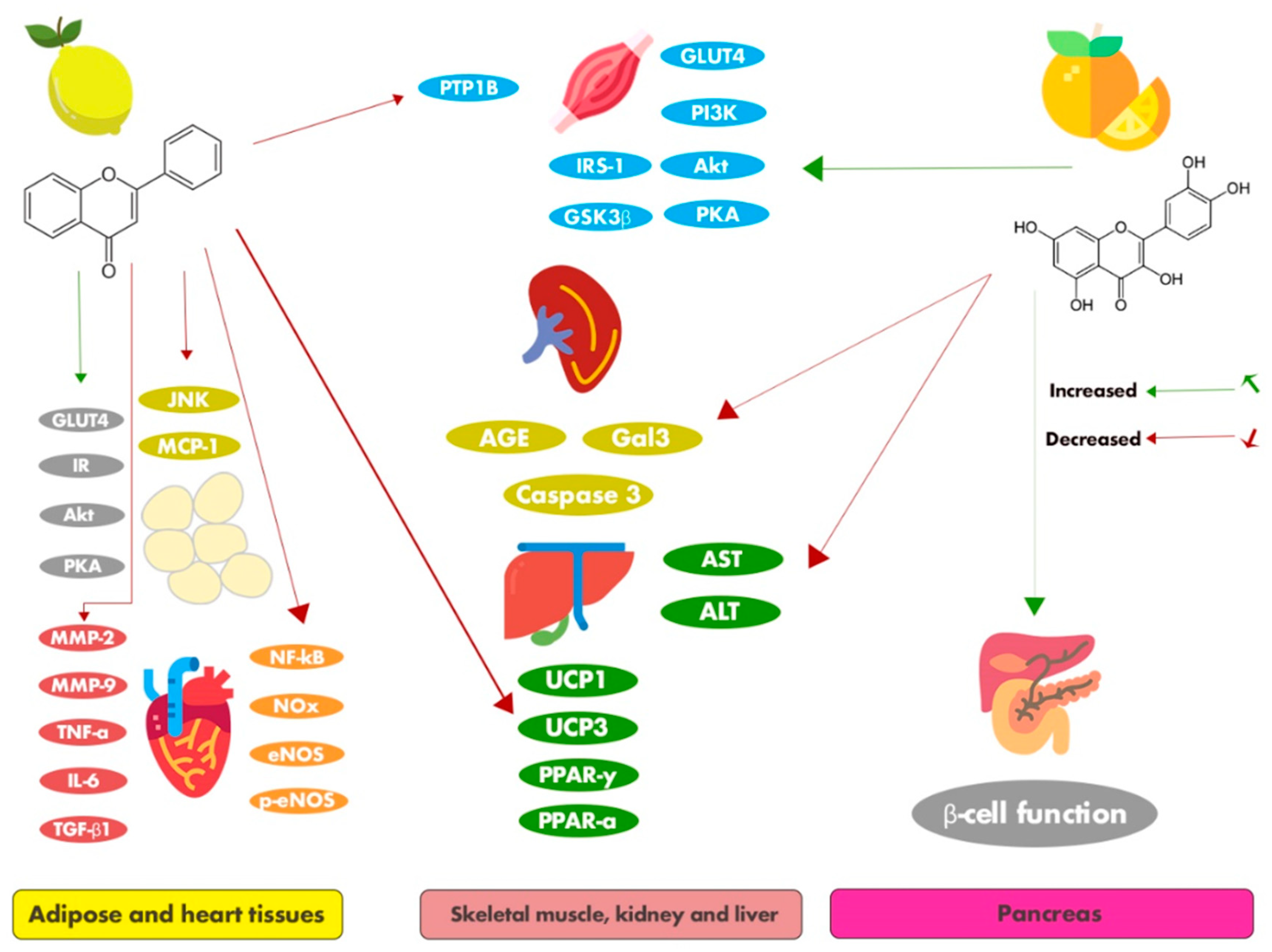
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Zech-Matterne, V.; Fiorentino, G. AGRUMED: Archaeology and History of Citrus Fruit in the Mediterranean: Acclimatization, Diversifications, Uses; Collection du Centre Jean Bérard, Publications du Centre Jean Bérard: Naples, Italy, 2017. [Google Scholar] [CrossRef]
- Luro, F.; Gatto, J.; Costantino, G.; Pailly, O. Analysis of genetic diversity in Citrus. Plant Genet. Resour. 2011, 9, 218–221. [Google Scholar] [CrossRef]
- Goulet, B.E.; Roda, F.; Hopkins, R. Hybridization in plants: Old ideas, new techniques. Plant Physiol. 2017, 173, 65–78. [Google Scholar] [CrossRef] [PubMed]
- Ollitrault, P.; Navarro, L. Citrus. In Fruit Breeding; Badenes, M., Byrne, D., Eds.; Springer: Boston, MA, USA, 2012; Volume 8, pp. 623–662. [Google Scholar] [CrossRef]
- Gill, H.; Garg, H. Citrus Pathology; IntechOpen: London, UK, 2017. [Google Scholar] [CrossRef]
- Man, M.Q.; Yang, B.; Elias, P.M. Benefits of hesperidin for cutaneous functions. Evid. Based Complement. Alternat. Med. 2019, 2019, 2676307. [Google Scholar] [CrossRef] [PubMed]
- Tripoli, E.; Guardia, M.L.; Giammanco, S.; Majo, D.D.; Giammanco, M. Citrus flavonoids: Molecular structure, biological activity and nutritional properties: A review. Food Chem. 2007, 104, 466–479. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Hernández Bautista, R.J.; Sandhu, M.A.; Hussein, O.E. Beneficial effects of citrus flavonoids on cardiovascular and metabolic health. Oxid. Med. Cell. Longev. 2019, 2019, 5484138. [Google Scholar] [CrossRef]
- Diabetes Atlas 9th edition 2019. Available online: https://www.diabetesatlas.org/en/ (accessed on 25 August 2020).
- Reach, G.; Pechtner, V.; Gentilella, R.; Corcos, A.; Ceriello, A. Clinical inertia and its impact on treatment intensification in people with type 2 diabetes mellitus. Diabetes Metab. 2017, 43, 501–511. [Google Scholar] [CrossRef]
- Cho, N.H.; Shaw, J.E.; Karuranga, S.; Huang, Y.; da Rocha Fernandes, J.D.; Ohlrogge, A.W.; Malanda, B. IDF Diabetes Atlas: Global estimates of diabetes prevalence for 2017 and projections for 2045. Diabetes Res. Clin. Pract. 2018, 138, 271–281. [Google Scholar] [CrossRef]
- Gray, S.P.; Jandeleit-Dahm, K. The pathobiology of diabetic vascular complications—Cardiovascular and kidney disease. J. Mol. Med. 2014, 92, 441–452. [Google Scholar] [CrossRef]
- Wong, E.; Backholer, K.; Gearon, E.; Harding, J.; Freak-Poli, R.; Stevenson, C.; Peeters, A. Diabetes and risk of physical disability in adults: A systematic review and meta-analysis. Lancet Diabetes Endocrinol. 2013, 1, 106–114. [Google Scholar] [CrossRef]
- Heinonen, S.E.; Genové, G.; Bengtsson, E.; Hübschle, T.; Åkesson, L.; Hiss, K.; Benardeau, A.; Ylä-Herttuala, S.; Jönsson-Rylander, A.-C.; Gomez, M.F. Animal models of diabetic macrovascular complications: Key players in the development of new therapeutic approaches. J. Diabetes Res. 2015, 2015, 404085. [Google Scholar] [CrossRef]
- Zhang, P.; Li, T.; Wu, X.; Nice, E.C.; Huang, C.; Zhang, Y. Oxidative stress and diabetes: Antioxidative strategies. Front. Med. 2020. [Google Scholar] [CrossRef] [PubMed]
- Yaribeygi, H.; Sathyapalan, T.; Atkin, S.L.; Sahebkar, A. Molecular mechanisms linking oxidative stress and diabetes mellitus. Oxid. Med. Cell. Longev. 2020, 2020, 8609213. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Schluesener, H. Health-promoting effects of the citrus flavanone hesperidin. Crit. Rev. Food Sci. Nutr. 2017, 57, 613–631. [Google Scholar] [CrossRef] [PubMed]
- Millar, C.L.; Duclos, Q.; Blesso, C.N. Effects of dietary flavonoids on reverse cholesterol transport, HDL metabolism, and HDL function. Adv. Nutr. 2017, 8, 226–239. [Google Scholar] [CrossRef] [PubMed]
- Rees, A.; Dodd, G.F.; Spencer, J.P.E. The effects of flavonoids on cardiovascular health: A review of human intervention trials and implications for cerebrovascular function. Nutrients 2018, 10, 1852. [Google Scholar] [CrossRef]
- Zaidun, N.H.; Thent, Z.C.; Latiff, A.A. Combating oxidative stress disorders with citrus flavonoid: Naringenin. Life Sci. 2018, 208, 111–122. [Google Scholar] [CrossRef]
- Zhang, X.; Li, X.; Fang, H.; Guo, F.; Li, F.; Chen, A.; Huang, S. Flavonoids as inducers of white adipose tissue browning and thermogenesis: Signalling pathways and molecular triggers. Nutr. Metab. 2019, 16, 47. [Google Scholar] [CrossRef]
- Kopustinskiene, D.M.; Jakstas, V.; Savickas, A.; Bernatoniene, J. Flavonoids as anticancer agents. Nutrients 2020, 12, 457. [Google Scholar] [CrossRef]
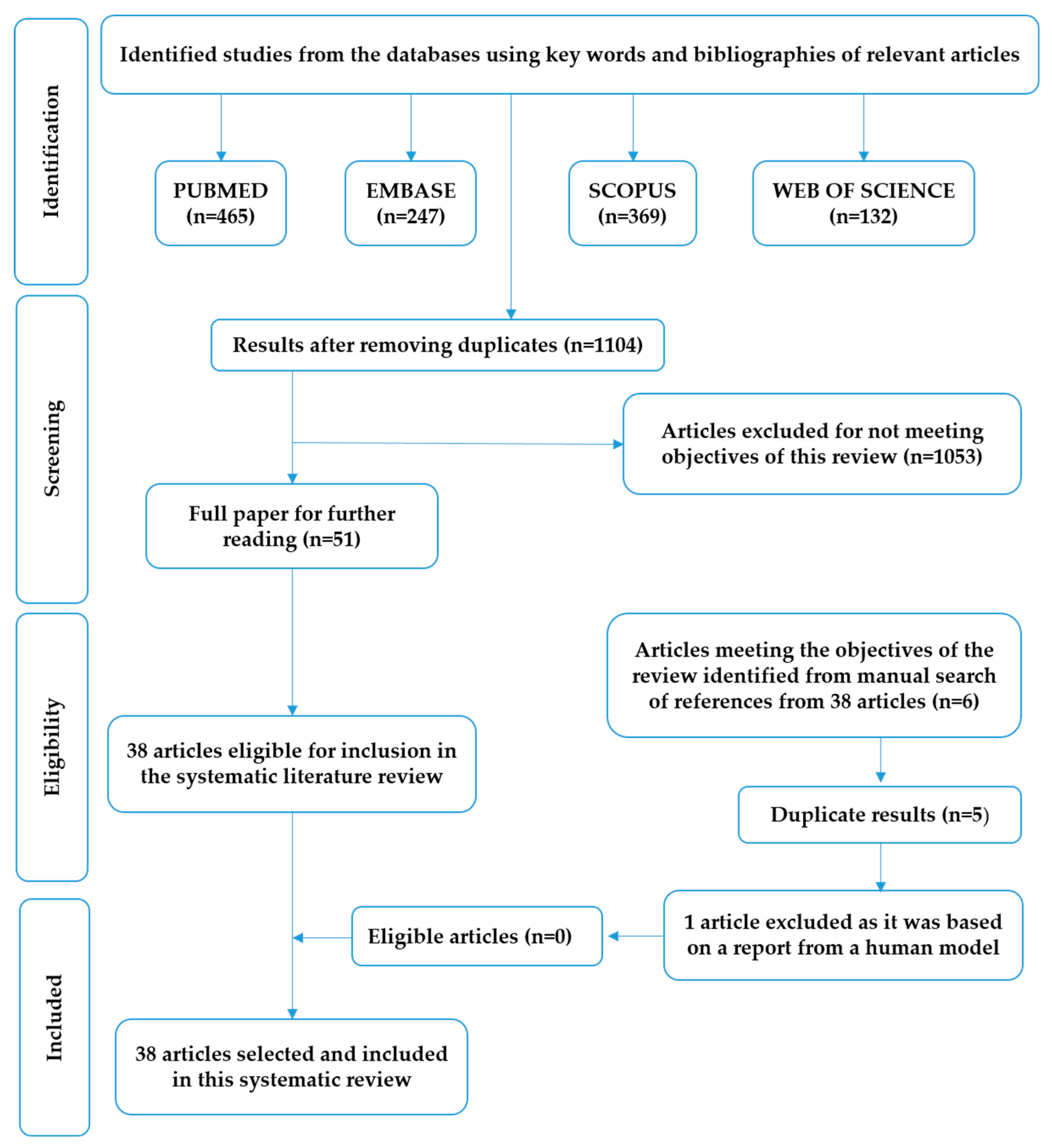
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. PLoS Med. 2009, 6, e1000097. [Google Scholar] [CrossRef]
- Żołnierczyk, A.K.; Mączka, W.K.; Grabarczyk, M.; Wińska, K.; Woźniak, E.; Anioł, M. Isoxanthohumol—Biologically active hop flavonoid. Fitoterapia 2015, 103, 71–82. [Google Scholar] [CrossRef]
- Štulíková, K.; Karabín, M.; Nešpor, J.; Dostálek, P. Therapeutic perspectives of 8-prenylnaringenin, a potent phytoestrogen from hops. Molecules 2018, 23, 660. [Google Scholar] [CrossRef] [PubMed]
- Negrão, R.; Costa, R.; Duarte, D.; Taveira Gomes, T.; Mendanha, M.; Moura, L.; Vasques, L.; Azevedo, I.; Soares, R. Angiogenesis and inflammation signaling are targets of beer polyphenols on vascular cells. J. Cell. Biochem. 2010, 111, 1270–1279. [Google Scholar] [CrossRef] [PubMed]
- Luís, C.; Costa, R.; Rodrigues, I.; Castela, Â.; Coelho, P.; Guerreiro, S.; Gomes, J.; Reis, C.; Soares, R. Xanthohumol and 8-prenylnaringenin reduce type 2 diabetes-associated oxidative stress by downregulating galectin-3. Porto Biomed. J. 2018, 4, e23. [Google Scholar] [CrossRef] [PubMed]
- Petrie, J.R.; Guzik, T.J.; Touyz, R.M. Diabetes, hypertension and cardiovascular disease: Clinical insights and vascular mechanisms. Can. J. Cardiol. 2018, 34, 575–584. [Google Scholar] [CrossRef]
- Munhoz, A.C.M.; Frode, T.S. Isolated compounds from natural products with potential antidiabetic activity—A systematic review. Curr. Diabetes Rev. 2018, 14, 36–106. [Google Scholar] [CrossRef] [PubMed]
- Rao, Y.K.; Lee, M.J.; Chen, K.; Lee, Y.C.; Wu, W.S.; Tzeng, Y.M. Insulin-mimetic action of rhoifolin and cosmosiin isolated from Citrus grandis (L.) Osbeck leaves: Enhanced adiponectin secretion and insulin receptor phosphorylation in 3T3-L1 cells. Evid. Based Complement. Alternat. Med. 2011, 2011, 624375. [Google Scholar] [CrossRef]
- Shalkami, A.S.; Hassan, M.; Bakr, A.G. Anti-inflammatory, antioxidant and anti-apoptotic activity of diosmin in acetic acid-induced ulcerative colitis. Hum. Exp. Toxicol. 2018, 37, 78–86. [Google Scholar] [CrossRef]
- Bogucka-Kocka, A.; Woźniak, M.; Feldo, M.; Kocki, J.; Szewczyk, K. Diosmin—Isolation techniques, determination in plant material and pharmaceutical formulations, and clinical use. Nat. Prod. Commun. 2013, 8, 1934578X1300800435. [Google Scholar] [CrossRef]
- Srinivasan, S.; Pari, L. Antihyperlipidemic effect of diosmin: A citrus flavonoid on lipid metabolism in experimental diabetic rats. J. Funct. Foods 2013, 5, 484–492. [Google Scholar] [CrossRef]
- Jain, D.; Bansal, M.K.; Dalvi, R.; Upganlawar, A.; Somani, R. Protective effect of diosmin against diabetic neuropathy in experimental rats. J. Integr. Med. 2014, 12, 35–41. [Google Scholar] [CrossRef]
- Hsu, C.C.; Lin, M.H.; Cheng, J.T.; Wu, M.C. Diosmin, a citrus nutrient, activates imidazoline receptors to alleviate blood glucose and lipids in type 1-like diabetic rats. Nutrients 2017, 9, 684. [Google Scholar] [CrossRef]
- Huang, H.; Li, L.; Shi, W.; Liu, H.; Yang, J.; Yuan, X.; Wu, L. The multifunctional effects of nobiletin and its metabolites in vivo and in vitro. Evid. Based Complement. Alternat. Med. 2016, 2016, 2918796. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-S.; Cha, B.-Y.; Choi, S.-S.; Choi, B.-K.; Yonezawa, T.; Teruya, T.; Nagai, K.; Woo, J.-T. Nobiletin improves obesity and insulin resistance in high-fat diet-induced obese mice. J. Nutr. Biochem. 2013, 24, 156–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Xie, J.; Ai, Z.; Su, J. Nobiletin-loaded micelles reduce ovariectomy-induced bone loss by suppressing osteoclastogenesis. Int. J. Nanomed. 2019, 14, 7839–7849. [Google Scholar] [CrossRef]
- Goh, J.X.H.; Tan, L.T.; Goh, J.K.; Chan, K.G.; Pusparajah, P.; Lee, L.H.; Goh, B.H. Nobiletin and derivatives: Functional compounds from citrus fruit peel for colon cancer chemoprevention. Cancers 2019, 11, 867. [Google Scholar] [CrossRef]
- Nohara, K.; Nemkov, T.; D‘Alessandro, A.; Yoo, S.H.; Chen, Z. Coordinate regulation of cholesterol and bile acid metabolism by the clock modifier nobiletin in metabolically challenged old mice. Int. J. Mol. Sci. 2019, 20, 4281. [Google Scholar] [CrossRef] [PubMed]
- Mulvihill, E.E.; Assini, J.M.; Lee, J.K.; Allister, E.M.; Sutherland, B.G.; Koppes, J.B.; Sawyez, C.G.; Edwards, J.Y.; Telford, D.E.; Charbonneau, A.; et al. Nobiletin attenuates VLDL overproduction, dyslipidemia, and atherosclerosis in mice with diet-induced insulin resistance. Diabetes 2011, 60, 1446–1457. [Google Scholar] [CrossRef] [PubMed]
- Onda, K.; Horike, N.; Suzuki, T.; Hirano, T. Polymethoxyflavonoids tangeretin and nobiletin increase glucose uptake in murine adipocytes. Phytother. Res. 2013, 27, 312–316. [Google Scholar] [CrossRef]
- Kanda, K.; Nishi, K.; Kadota, A.; Nishimoto, S.; Liu, M.C.; Sugahara, T. Nobiletin suppresses adipocyte differentiation of 3T3-L1 cells by an insulin and IBMX mixture induction. Biochim. Biophys. Acta 2012, 1820, 461–468. [Google Scholar] [CrossRef]
- Parkar, N.A.; Bhatt, L.K.; Addepalli, V. Efficacy of nobiletin, a citrus flavonoid, in the treatment of the cardiovascular dysfunction of diabetes in rats. Food Funct. 2016, 7, 3121–3129. [Google Scholar] [CrossRef]
- Zhang, N.; Yang, Z.; Xiang, S.Z.; Jin, Y.G.; Wei, W.Y.; Bian, Z.Y.; Deng, W.; Tang, Q.Z. Nobiletin attenuates cardiac dysfunction, oxidative stress, and inflammatory in streptozotocin: Induced diabetic cardiomyopathy. Mol. Cell. Biochem. 2016, 417, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Morrow, N.M.; Burke, A.C.; Samsoondar, J.P.; Seigel, K.E.; Wang, A.; Telford, D.E.; Sutherland, B.G.; O’Dwyer, C.; Steinberg, G.R.; Fullerton, M.D.; et al. The citrus flavonoid nobiletin confers protection from metabolic dysregulation in high-fat-fed mice independent of AMPK. J. Lipid Res. 2020, 61, 387–402. [Google Scholar] [CrossRef] [PubMed]
- Refaat, J.; Desoukey, S.Y.; Ramadan, M.A.; Kamel, M.S. Rhoifolin: A review of sources and biological activities. Int. J. Pharmacogn. 2015, 2, 102–109. [Google Scholar] [CrossRef]
- Liao, S.; Song, F.; Feng, W.; Ding, X.; Yao, J.; Song, H.; Liu, Y.; Ma, S.; Wang, Z.; Lin, X.; et al. Rhoifolin ameliorates titanium particle-stimulated osteolysis and attenuates osteoclastogenesis via RANKL-induced NF-κB and MAPK pathways. J. Cell. Physiol. 2019, 234, 17600–17611. [Google Scholar] [CrossRef] [PubMed]
- Phan, V.K.; Nguyen, T.M.; Minh, C.V.; Nguyen, H.K.; Nguyen, H.D.; Nguyen, P.T.; Nguyen, X.C.; Nguyen, H.N.; Nguyen, X.N.; Heyden, Y.V.; et al. Two new C-glucosyl benzoic acids and flavonoids from Mallotus nanus and their antioxidant activity. Arch. Pharm. Res. 2010, 33, 203–208. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Ren, Y.; Lin, D.; Peng, S.; Zhong, B.; Ma, Z. The anti-inflammatory properties of Citrus wilsonii tanaka extract in LPS-induced RAW 264.7 and primary mouse bone marrow-derived dendritic cells. Molecules 2017, 22, 1213. [Google Scholar] [CrossRef]
- Sultana, B.; Yaqoob, S.; Zafar, Z.; Bhatti, H.N. Escalation of liver malfunctioning: A step toward Herbal Awareness. J. Ethnopharmacol. 2018, 216, 104–119. [Google Scholar] [CrossRef]
- Koyuncu, I. Evaluation of anticancer, antioxidant activity and phenolic compounds of Artemisia absinthium L. Extract. Cell. Mol. Biol. 2018, 64, 25–34. [Google Scholar] [CrossRef]
- Kim, G.S.; Park, H.J.; Woo, J.H.; Kim, M.K.; Koh, P.O.; Min, W.; Ko, Y.G.; Kim, C.H.; Won, C.K.; Cho, J.H. Citrus aurantium flavonoids inhibit adipogenesis through the Akt signaling pathway in 3T3-L1 cells. BMC Complement. Altern. Med. 2012, 12, 31. [Google Scholar] [CrossRef]
- Tsutsumi, R.; Yoshida, T.; Nii, Y.; Okahisa, N.; Iwata, S.; Tsukayama, M.; Hashimoto, R.; Taniguchi, Y.; Sakaue, H.; Hosaka, T.; et al. Sudachitin, a polymethoxylated flavone, improves glucose and lipid metabolism by increasing mitochondrial biogenesis in skeletal muscle. Nutr. Metab. 2014, 11, 32. [Google Scholar] [CrossRef]
- Yoshida, H.; Watanabe, H.; Ishida, A.; Watanabe, W.; Narumi, K.; Atsumi, T.; Sugita, C.; Kurokawa, M. Naringenin suppresses macrophage infiltration into adipose tissue in an early phase of high-fat diet-induced obesity. Biochem. Biophys. Res. Commun. 2014, 454, 95–101. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, A.M.; Ahmed, O.M.; Ashour, M.B.; Abdel-Moneim, A. In vivo and in vitro antidiabetic effects of citrus flavonoids; a study on the mechanism of action. Int. J. Diabetes Dev. Ctries. 2015, 35, 250–263. [Google Scholar] [CrossRef]
- Dhanya, R.; Arya, A.D.; Nisha, P.; Jayamurthy, P. Quercetin, a lead compound against type 2 diabetes ameliorates glucose uptake via AMPK pathway in skeletal muscle cell line. Front. Pharmacol. 2017, 8, 336. [Google Scholar] [CrossRef]
- Liu, W.; Liou, S.S.; Hong, T.Y.; Liu, I.M. Protective effects of hesperidin (citrus flavonone) on high glucose induced oxidative stress and apoptosis in a cellular model for diabetic retinopathy. Nutrients 2017, 9, 1323. [Google Scholar] [CrossRef] [PubMed]
- Zareei, S.; Boojar, M.M.A.; Amanlou, M. Inhibition of liver alanine aminotransferase and aspartate aminotransferase by hesperidin and its aglycone hesperetin: An in vitro and in silico study. Life Sci. 2017, 178, 49–55. [Google Scholar] [CrossRef]
- Chen, F.; Ma, Y.; Sun, Z.; Zhu, X. Tangeretin inhibits high glucose-induced extracellular matrix accumulation in human glomerular mesangial cells. Biomed. Pharmacother. 2018, 102, 1077–1083. [Google Scholar] [CrossRef]
- Shukla, K.; Sonowal, H.; Saxena, A.; Ramana, K.V. Didymin prevents hyperglycemia-induced human umbilical endothelial cells dysfunction and death. Biochem. Pharmacol. 2018, 152, 1–10. [Google Scholar] [CrossRef]
- Ali, M.Y.; Zaib, S.; Rahman, M.M.; Jannat, S.; Iqbal, J.; Park, S.K.; Chang, M.S. Didymin, a dietary citrus flavonoid exhibits anti-diabetic complications and promotes glucose uptake through the activation of PI3K/Akt signaling pathway in insulin-resistant HepG2 cells. Chem. Biol. Interact. 2019, 305, 180–194. [Google Scholar] [CrossRef]
- Tseng, Y.-T.; Hsu, H.-T.; Lee, T.-Y.; Chang, W.-H.; Lo, Y.-C. Naringenin, a dietary flavanone, enhances insulin-like growth factor 1 receptor-mediated antioxidant defense and attenuates methylglyoxal-induced neurite damage and apoptotic death. Nutr. Neurosci. 2019, 1–11. [Google Scholar] [CrossRef]
- Tsuhako, R.; Yoshida, H.; Sugita, C.; Kurokawa, M. Naringenin suppresses neutrophil infiltration into adipose tissue in high-fat diet-induced obese mice. J. Nat. Med. 2019, 74, 229–237. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, H.; Takaishi, Y.; Tanaka, N.; Tsuchiya, K.; Shibata, H.; Higuti, T. Chemical constituents from the peels of Citrus sudachi. J. Nat. Prod. 2006, 69, 1177–1179. [Google Scholar] [CrossRef] [PubMed]
- Ohyama, Y.; Ito, J.; Kitano, V.J.; Shimada, J.; Hakeda, Y. The polymethoxy flavonoid sudachitin suppresses inflammatory bone destruction by directly inhibiting osteoclastogenesis due to reduced ROS production and MAPK activation in osteoclast precursors. PLoS ONE 2018, 13, e0191192. [Google Scholar] [CrossRef] [PubMed]
- Abe, S.; Hirose, S.; Nishitani, M.; Yoshida, I.; Tsukayama, M.; Tsuji, A.; Yuasa, K. Citrus peel polymethoxyflavones, sudachitin and nobiletin, induce distinct cellular responses in human keratinocyte HaCaT cells. Biosci. Biotechnol. Biochem. 2018, 82, 2064–2071. [Google Scholar] [CrossRef]
- Mitani, M.; Minatogawa, Y.; Nakamoto, A.; Nakamoto, M.; Shuto, E.; Nii, Y.; Sakai, T. Sudachitin, polymethoxyflavone from Citrus sudachi, enhances antigen-specific cellular and humoral immune responses in BALB/c mice. J. Clin. Biochem. Nutr. 2019, 64, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Hosokawa, Y.; Hosokawa, I.; Ozaki, K.; Matsuo, T. Sudachitin inhibits matrix metalloproteinase-1 and -3 production in tumor necrosis factor-α-stimulated human periodontal ligament cells. Inflammation 2019, 42, 1456–1462. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Ahmadi, Z.; Mohammadinejad, R.; Afshar, E.G. Tangeretin: A mechanistic review of its pharmacological and therapeutic effects. J. Basic Clin. Physiol. Pharmacol. 2020, 31, 20190191. [Google Scholar] [CrossRef]
- Sundaram, R.; Shanthi, P.; Sachdanandam, P. Effect of tangeretin, a polymethoxylated flavone on glucose metabolism in streptozotocin-induced diabetic rats. Phytomedicine 2014, 21, 793–799. [Google Scholar] [CrossRef]
- Sundaram, R.; Shanthi, P.; Sachdanandam, P. Tangeretin, a polymethoxylated flavone, modulates lipid homeostasis and decreases oxidative stress by inhibiting NF-κB activation and proinflammatory cytokines in cardiac tissue of streptozotocin-induced diabetic rats. J. Funct. Foods 2015, 16, 315–333. [Google Scholar] [CrossRef]
- Yao, Q.; Lin, M.T.; Zhu, Y.D.; Xu, H.L.; Zhao, Y.Z. Recent trends in potential therapeutic applications of the dietary flavonoid didymin. Molecules 2018, 23, 2547. [Google Scholar] [CrossRef]
- Hung, J.Y.; Hsu, Y.L.; Ko, Y.C.; Tsai, Y.M.; Yang, C.J.; Huang, M.S.; Kuo, P.L. Didymin, a dietary flavonoid glycoside from citrus fruits, induces Fas-mediated apoptotic pathway in human non-small-cell lung cancer cells in vitro and in vivo. Lung Cancer 2010, 68, 366–374. [Google Scholar] [CrossRef]
- Singhal, J.; Nagaprashantha, L.D.; Vatsyayan, R.; Awasthi, S.; Singhal, S.S. Didymin induces apoptosis by inhibiting N-Myc and upregulating RKIP in neuroblastoma. Cancer Prev. Res. 2012, 5, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Huang, Q.; Bai, F.; Lin, J.; Nie, J.; Lu, S.; Lu, C.; Huang, R.; Lu, Z.; Lin, X. Didymin induces apoptosis through mitochondrial dysfunction and up-regulation of RKIP in human hepatoma cells. Chem. Biol. Interact. 2017, 261, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Peterson, J.J.; Beecher, G.R.; Bhagwat, S.A.; Dwyer, J.T.; Gebhardt, S.E.; Haytowitz, D.B.; Holden, J.M. Flavanones in grapefruit, lemons, and limes: A compilation and review of the data from the analytical literature. J. Food Compos. Anal. 2006, 19, S74–S80. [Google Scholar] [CrossRef]
- Rangel-Huerta, O.D.; Aguilera, C.M.; Martin, M.V.; Soto, M.J.; Rico, M.C.; Vallejo, F.; Tomas-Barberan, F.; Perez-de-la-Cruz, A.J.; Gil, A.; Mesa, M.D. Normal or high polyphenol concentration in orange juice affects antioxidant activity, blood pressure, and body weight in obese or overweight adults. J. Nutr. 2015, 145, 1808–1816. [Google Scholar] [CrossRef]
- Nandakumar, N.; Balasubramanian, M.P. Hesperidin a citrus bioflavonoid modulates hepatic biotransformation enzymes and enhances intrinsic antioxidants in experimental breast cancer rats challenged with 7, 12-dimethylbenz (a) anthracene. J. Exp. Ther. Oncol. 2012, 9, 321–335. [Google Scholar]
- Mahmoud, A.M.; Mohammed, H.M.; Khadrawy, S.M.; Galaly, S.R. Hesperidin protects against chemically induced hepatocarcinogenesis via modulation of Nrf2/ARE/HO-1, PPARγ and TGF-β1/Smad3 signaling, and amelioration of oxidative stress and inflammation. Chem. Biol. Interact. 2017, 277, 146–158. [Google Scholar] [CrossRef]
- Carballo-Villalobos, A.I.; González-Trujano, M.E.; Alvarado-Vázquez, N.; López-Muñoz, F.J. Pro-inflammatory cytokines involvement in the hesperidin antihyperalgesic effects at peripheral and central levels in a neuropathic pain model. Inflammopharmacology 2017, 25, 265–269. [Google Scholar] [CrossRef]
- Akiyama, S.; Katsumata, S.; Suzuki, K.; Ishimi, Y.; Wu, J.; Uehara, M. Dietary hesperidin exerts hypoglycemic and hypolipidemic effects in streptozotocin-induced marginal type 1 diabetic rats. J. Clin. Biochem. Nutr. 2010, 46, 87–92. [Google Scholar] [CrossRef]
- Mahmoud, A.M.; Ashour, M.B.; Abdel-Moneim, A.; Ahmed, O.M. Hesperidin and naringin attenuate hyperglycemia-mediated oxidative stress and proinflammatory cytokine production in high fat fed/streptozotocin-induced type 2 diabetic rats. J. Diabetes Complicat. 2012, 26, 483–490. [Google Scholar] [CrossRef]
- El-Marasy, S.A.; Abdallah, H.M.; El-Shenawy, S.M.; El-Khatib, A.S.; El-Shabrawy, O.A.; Kenawy, S.A. Anti-depressant effect of hesperidin in diabetic rats. Can. J. Physiol. Pharmacol. 2014, 92, 945–952. [Google Scholar] [CrossRef]
- Visnagri, A.; Kandhare, A.D.; Chakravarty, S.; Ghosh, P.; Bodhankar, S.L. Hesperidin, a flavanoglycone attenuates experimental diabetic neuropathy via modulation of cellular and biochemical marker to improve nerve functions. Pharm. Biol. 2014, 52, 814–828. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Kandhare, A.D.; Mukherjee, A.A.; Bodhankar, S.L. Hesperidin, a plant flavonoid accelerated the cutaneous wound healing in streptozotocin-induced diabetic rats: Role of TGF-ß/Smads and Ang-1/Tie-2 signaling pathways. EXCLI J. 2018, 17, 399–419. [Google Scholar] [CrossRef] [PubMed]
- Dokumacioglu, E.; Iskender, H.; Sen, T.M.; Ince, I.; Dokumacioglu, A.; Kanbay, Y.; Erbas, E.; Saral, S. The effects of hesperidin and quercetin on serum tumor necrosis factor-alpha and interleukin-6 levels in streptozotocin-induced diabetes model. Pharmacogn. Mag. 2018, 14, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Tilg, H.; Hotamisligil, G.S. Nonalcoholic fatty liver disease: Cytokine-adipokine interplay and regulation of insulin resistance. Gastroenterology 2006, 131, 934–945. [Google Scholar] [CrossRef] [PubMed]
- Mehta, S.; Farmer, J.A. Obesity and inflammation: A new look at an old problem. Curr. Atheroscler. Rep. 2007, 9, 134–138. [Google Scholar] [CrossRef]
- Somerset, S.M.; Johannot, L. Dietary flavonoid sources in Australian adults. Nutr. Cancer 2008, 60, 442–449. [Google Scholar] [CrossRef]
- Furtado, A.F.; Nunes, M.A.; Ribeiro, M.H. Hesperidinase encapsulation towards hesperitin production targeting improved bioavailability. J. Mol. Recognit. 2012, 25, 595–603. [Google Scholar] [CrossRef]
- Paramita, P.; Sethu, S.N.; Subhapradha, N.; Ragavan, V.; Ilangovan, R.; Balakrishnan, A.; Srinivasan, N.; Murugesan, R.; Moorthi, A. Neuro-protective effects of nano-formulated hesperetin in a traumatic brain injury model of Danio rerio. Drug Chem. Toxicol. 2020, 1–8. [Google Scholar] [CrossRef]
- Kottaiswamy, A.; Kizhakeyil, A.; Padmanaban, A.; Bushra, F.; Vijay, V.R.; Lee, P.S.; Verma, N.K.; Kalaiselvan, P.; Samuel, S. The citrus flavanone hesperetin induces apoptosis in CTCL cells via STAT3/Notch1/NFκB-mediated signaling axis. Anticancer Agents Med. Chem. 2020. [Google Scholar] [CrossRef]
- Jo, S.H.; Kim, M.E.; Cho, J.H.; Lee, Y.; Lee, J.; Park, Y.D.; Lee, J.S. Hesperetin inhibits neuroinflammation on microglia by suppressing inflammatory cytokines and MAPK pathways. Arch. Pharm. Res. 2019, 42, 695–703. [Google Scholar] [CrossRef]
- Kim, W.J.; Lee, S.E.; Park, Y.G.; Jeong, S.G.; Kim, E.Y.; Park, S.P. Antioxidant hesperetin improves the quality of porcine oocytes during aging in vitro. Mol. Reprod. Dev. 2019, 86, 32–41. [Google Scholar] [CrossRef]
- Chen, X.; Wei, W.; Li, Y.; Huang, J.; Ci, X. Hesperetin relieves cisplatin-induced acute kidney injury by mitigating oxidative stress, inflammation and apoptosis. Chem. Biol. Interact. 2019, 308, 269–278. [Google Scholar] [CrossRef] [PubMed]
- Jayaraman, R.; Subramani, S.; Sheik Abdullah, S.H.; Udaiyar, M. Antihyperglycemic effect of hesperetin, a citrus flavonoid, extenuates hyperglycemia and exploring the potential role in antioxidant and antihyperlipidemic in streptozotocin-induced diabetic rats. Biomed. Pharmacother. 2018, 97, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Samie, A.; Sedaghat, R.; Baluchnejadmojarad, T.; Roghani, M. Hesperetin, a citrus flavonoid, attenuates testicular damage in diabetic rats via inhibition of oxidative stress, inflammation, and apoptosis. Life Sci. 2018, 210, 132–139. [Google Scholar] [CrossRef]
- Fernandes, A.A.; Novelli, E.L.; Okoshi, K.; Okoshi, M.P.; Di Muzio, B.P.; Guimarães, J.F.; Fernandes Junior, A. Influence of rutin treatment on biochemical alterations in experimental diabetes. Biomed. Pharmacother. 2010, 64, 214–219. [Google Scholar] [CrossRef] [PubMed]
- Fallahi, F.; Roghani, M.; Moghadami, S. Citrus flavonoid naringenin improves aortic reactivity in streptozotocin-diabetic rats. Indian J. Pharmacol. 2012, 44, 382–386. [Google Scholar] [CrossRef]
- Hasanein, P.; Fazeli, F. Role of naringenin in protection against diabetic hyperalgesia and tactile allodynia in male Wistar rats. J. Physiol. Biochem. 2014, 70, 997–1006. [Google Scholar] [CrossRef]
- Jia, S.; Hu, Y.; Zhang, W.; Zhao, X.; Chen, Y.; Sun, C.; Li, X.; Chen, K. Hypoglycemic and hypolipidemic effects of neohesperidin derived from Citrus aurantium L. in diabetic KK-A(y) mice. Food Funct. 2015, 6, 878–886. [Google Scholar] [CrossRef]
- Malakul, W.; Pengnet, S.; Kumchoom, C.; Tunsophon, S. Naringin ameliorates endothelial dysfunction in fructose-fed rats. Exp. Ther. Med. 2018, 15, 3140–3146. [Google Scholar] [CrossRef]
- Alam, M.A.; Subhan, N.; Rahman, M.M.; Uddin, S.J.; Reza, H.M.; Sarker, S.D. Effect of citrus flavonoids, naringin and naringenin, on metabolic syndrome and their mechanisms of action. Adv. Nutr. 2014, 5, 404–417. [Google Scholar] [CrossRef]
- Chen, R.; Qi, Q.L.; Wang, M.T.; Li, Q.Y. Therapeutic potential of naringin: An overview. Pharm. Biol. 2016, 54, 3203–3210. [Google Scholar] [CrossRef] [PubMed]
- Kandhare, A.D.; Raygude, K.S.; Ghosh, P.; Ghule, A.E.; Bodhankar, S.L. Neuroprotective effect of naringin by modulation of endogenous biomarkers in streptozotocin induced painful diabetic neuropathy. Fitoterapia 2012, 83, 650–659. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, S.F.; Attia, S.M.; Bakheet, S.A.; Zoheir, K.M.A.; Ansari, M.A.; Korashy, H.M.; Abdel-Hamied, H.E.; Ashour, A.E.; Abd-Allah, A.R.A. Naringin attenuates the development of carrageenan-induced acute lung inflammation through inhibition of NF-κb, STAT3 and pro-inflammatory mediators and enhancement of IκBα and anti-inflammatory cytokines. Inflammation 2015, 38, 846–857. [Google Scholar] [CrossRef] [PubMed]
- Jiao, H.Y.; Su, W.W.; Li, P.B.; Liao, Y.; Zhou, Q.; Zhu, N.; He, L. Therapeutic effects of naringin in a guinea pig model of ovalbumin-induced cough-variant asthma. Pulm. Pharmacol. Ther. 2015, 33, 59–65. [Google Scholar] [CrossRef]
- Xiong, G.; Liu, S.; Gao, J.; Wang, S. Naringin protects ovalbumin-induced airway inflammation in a mouse model of asthma. Inflammation 2016, 39, 891–899. [Google Scholar] [CrossRef]
- Zhang, Y.S.; Li, Y.; Wang, Y.; Sun, S.Y.; Jiang, T.; Li, C.; Cui, S.X.; Qu, X.J. Naringin, a natural dietary compound, prevents intestinal tumorigenesis in Apc (Min/+) mouse model. J. Cancer Res. Clin. Oncol. 2016, 142, 913–925. [Google Scholar] [CrossRef]
- Kwatra, M.; Jangra, A.; Mishra, M.; Sharma, Y.; Ahmed, S.; Ghosh, P.; Kumar, V.; Vohora, D.; Khanam, R. Naringin and sertraline ameliorate doxorubicin-induced behavioral deficits through modulation of serotonin level and mitochondrial complexes protection pathway in rat hippocampus. Neurochem. Res. 2016, 41, 2352–2366. [Google Scholar] [CrossRef]
- Sachdeva, A.K.; Kuhad, A.; Chopra, K. Naringin ameliorates memory deficits in experimental paradigm of Alzheimer’s disease by attenuating mitochondrial dysfunction. Pharmacol. Biochem. Behav. 2014, 127, 101–110. [Google Scholar] [CrossRef] [PubMed]
- Vij, G.; Gupta, A.; Chopra, K. Modulation of antigen-induced chronic fatigue in mouse model of water immersion stress by naringin, a polyphenolic antioxidant. Fundam. Clin. Pharmacol. 2009, 23, 331–337. [Google Scholar] [CrossRef]
- Chtourou, Y.; Aouey, B.; Kebieche, M.; Fetoui, H. Protective role of naringin against cisplatin induced oxidative stress, inflammatory response and apoptosis in rat striatum via suppressing ROS-mediated NF-κB and P53 signaling pathways. Chem. Biol. Interact. 2015, 239, 76–86. [Google Scholar] [CrossRef]
- Zhang, J.; Sun, C.; Yan, Y.; Chen, Q.; Luo, F.; Zhu, X.; Li, X.; Chen, K. Purification of naringin and neohesperidin from Huyou (Citrus changshanensis) fruit and their effects on glucose consumption in human HepG2 cells. Food Chem. 2012, 135, 1471–1478. [Google Scholar] [CrossRef] [PubMed]
- Gong, N.; Zhang, B.; Yang, D.; Gao, Z.; Du, G.; Lu, Y. Development of new reference material neohesperidin for quality control of dietary supplements. J. Sci. Food Agric. 2015, 95, 1885–1891. [Google Scholar] [CrossRef] [PubMed]
- Hwang, S.-L.; Yen, G.-C. Neuroprotective effects of the citrus flavanones against H2 O2 -induced cytotoxicity in PC12 cells. J. Agric. Food Chem. 2008, 56, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Bellocco, E.; Barreca, D.; Laganà, G.; Leuzzi, U.; Tellone, E.; Ficarra, S.; Kotyk, A.; Galtieri, A. Influence of l-rhamnosyl-d-glucosyl derivatives on properties and biological interaction of flavonoids. Mol. Cell. Biochem. 2009, 321, 165–171. [Google Scholar] [CrossRef]
- Zhao, T.; Hu, S.; Ma, P.; Che, D.; Liu, R.; Zhang, Y.; Wang, J.; Li, C.; Ding, Y.; Fu, J.; et al. Neohesperidin suppresses IgE-mediated anaphylactic reactions and mast cell activation via Lyn-PLC-Ca(2+) pathway. Phytother. Res. 2019, 33, 2034–2043. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Fang, Y.; Jiang, F.; Li, L.; Zhou, H.; Xu, X.; Ning, W. Neohesperidin inhibits TGF-β1/Smad3 signaling and alleviates bleomycin-induced pulmonary fibrosis in mice. Eur. J. Pharmacol. 2019, 864, 172712. [Google Scholar] [CrossRef]
- Tan, Z.; Cheng, J.; Liu, Q.; Zhou, L.; Kenny, J.; Wang, T.; Lin, X.; Yuan, J.; Quinn, J.M.W.; Tickner, J.; et al. Neohesperidin suppresses osteoclast differentiation, bone resorption and ovariectomised-induced osteoporosis in mice. Mol. Cell. Endocrinol. 2017, 439, 369–378. [Google Scholar] [CrossRef]
- Jiang, C.H.; Sun, T.L.; Xiang, D.X.; Wei, S.S.; Li, W.Q. Anticancer activity and mechanism of xanthohumol: A prenylated flavonoid from hops (Humulus lupulus L.). Front. Pharmacol. 2018, 9, 530. [Google Scholar] [CrossRef]
- Li, F.; Yao, Y.; Huang, H.; Hao, H.; Ying, M. Xanthohumol attenuates cisplatin-induced nephrotoxicity through inhibiting NF-κB and activating Nrf2 signaling pathways. Int. Immunopharmacol. 2018, 61, 277–282. [Google Scholar] [CrossRef]
- Wang, S.; Yang, C.; Tu, H.; Zhou, J.; Liu, X.; Cheng, Y.; Luo, J.; Deng, X.; Zhang, H.; Xu, J. Characterization and metabolic diversity of flavonoids in Citrus species. Sci. Rep. 2017, 7, 10549. [Google Scholar] [CrossRef]
- Sharma, A.; Sharma, P.; Singh Tuli, H.; Sharma, A.K. Phytochemical and Pharmacological Properties of Flavonols. In eLS; John Wiley & Sons: Chichester, UK, 2018; pp. 1–12. [Google Scholar] [CrossRef]
- Gautam, R.; Singh, M.; Gautam, S.; Rawat, J.K.; Saraf, S.A.; Kaithwas, G. Rutin attenuates intestinal toxicity induced by Methotrexate linked with anti-oxidative and anti-inflammatory effects. BMC Complement. Altern. Med. 2016, 16, 99. [Google Scholar] [CrossRef] [PubMed]
- Klimczak, I.; Małecka, M.; Szlachta, M.; Gliszczyńska-Świgło, A. Effect of storage on the content of polyphenols, vitamin C and the antioxidant activity of orange juices. J. Food Compos. Anal. 2007, 20, 313–322. [Google Scholar] [CrossRef]
- Stuetz, W.; Prapamontol, T.; Hongsibsong, S.; Biesalski, H. Polymethoxylated flavones, flavanone glycosides, carotenoids, and antioxidants in different cultivation types of tangerines (Citrus reticulata Blanco cv. Sainampueng) from Northern Thailand. J. Agric. Food Chem. 2010, 58, 6069–6074. [Google Scholar] [CrossRef] [PubMed]
- Ramful, D.; Bahorun, T.; Bourdon, E.; Tarnus, E.; Aruoma, O.I. Bioactive phenolics and antioxidant propensity of flavedo extracts of Mauritian citrus fruits: Potential prophylactic ingredients for functional foods application. Toxicology 2010, 278, 75–87. [Google Scholar] [CrossRef] [PubMed]
- Gattuso, G.; Barreca, D.; Gargiulli, C.; Leuzzi, U.; Caristi, C. Flavonoid composition of citrus juices. Molecules 2007, 12, 1641–1673. [Google Scholar] [CrossRef]




© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gandhi, G.R.; Vasconcelos, A.B.S.; Wu, D.-T.; Li, H.-B.; Antony, P.J.; Li, H.; Geng, F.; Gurgel, R.Q.; Narain, N.; Gan, R.-Y. Citrus Flavonoids as Promising Phytochemicals Targeting Diabetes and Related Complications: A Systematic Review of In Vitro and In Vivo Studies. Nutrients 2020, 12, 2907. https://doi.org/10.3390/nu12102907
Gandhi GR, Vasconcelos ABS, Wu D-T, Li H-B, Antony PJ, Li H, Geng F, Gurgel RQ, Narain N, Gan R-Y. Citrus Flavonoids as Promising Phytochemicals Targeting Diabetes and Related Complications: A Systematic Review of In Vitro and In Vivo Studies. Nutrients. 2020; 12(10):2907. https://doi.org/10.3390/nu12102907
Chicago/Turabian StyleGandhi, Gopalsamy Rajiv, Alan Bruno Silva Vasconcelos, Ding-Tao Wu, Hua-Bin Li, Poovathumkal James Antony, Hang Li, Fang Geng, Ricardo Queiroz Gurgel, Narendra Narain, and Ren-You Gan. 2020. "Citrus Flavonoids as Promising Phytochemicals Targeting Diabetes and Related Complications: A Systematic Review of In Vitro and In Vivo Studies" Nutrients 12, no. 10: 2907. https://doi.org/10.3390/nu12102907
APA StyleGandhi, G. R., Vasconcelos, A. B. S., Wu, D.-T., Li, H.-B., Antony, P. J., Li, H., Geng, F., Gurgel, R. Q., Narain, N., & Gan, R.-Y. (2020). Citrus Flavonoids as Promising Phytochemicals Targeting Diabetes and Related Complications: A Systematic Review of In Vitro and In Vivo Studies. Nutrients, 12(10), 2907. https://doi.org/10.3390/nu12102907