Brain and Cognitive Development in Adolescents with Anorexia Nervosa: A Systematic Review of fMRI Studies


Abstract
1. Introduction
1.1. Impact of Genetic and Environmental Factors on the Adolescent Brain
1.2. Aim of Our Review
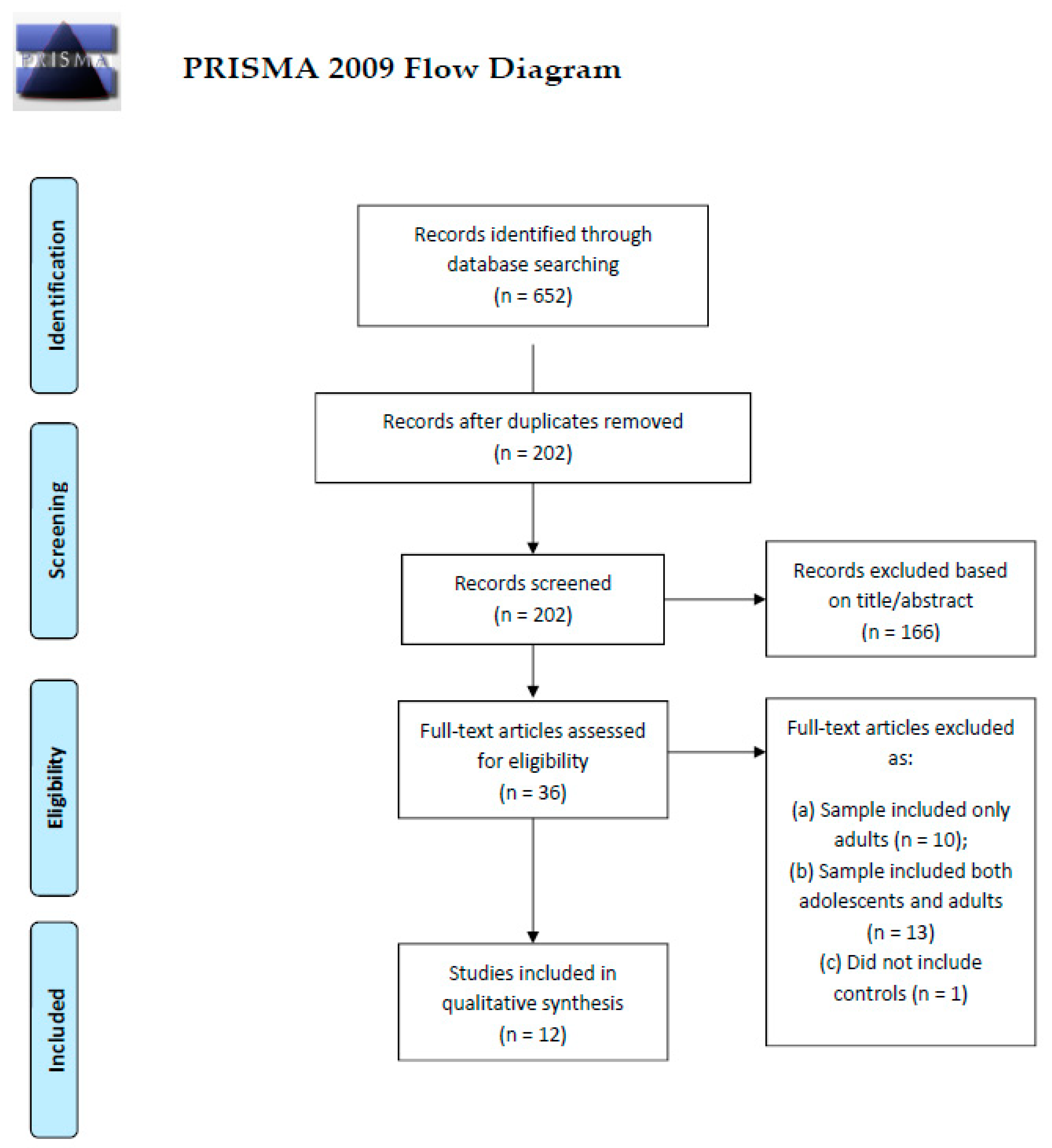
2. Methods
2.1. Search Strategy and Eligibility Criteria
2.2. Quality Assessment
3. Results
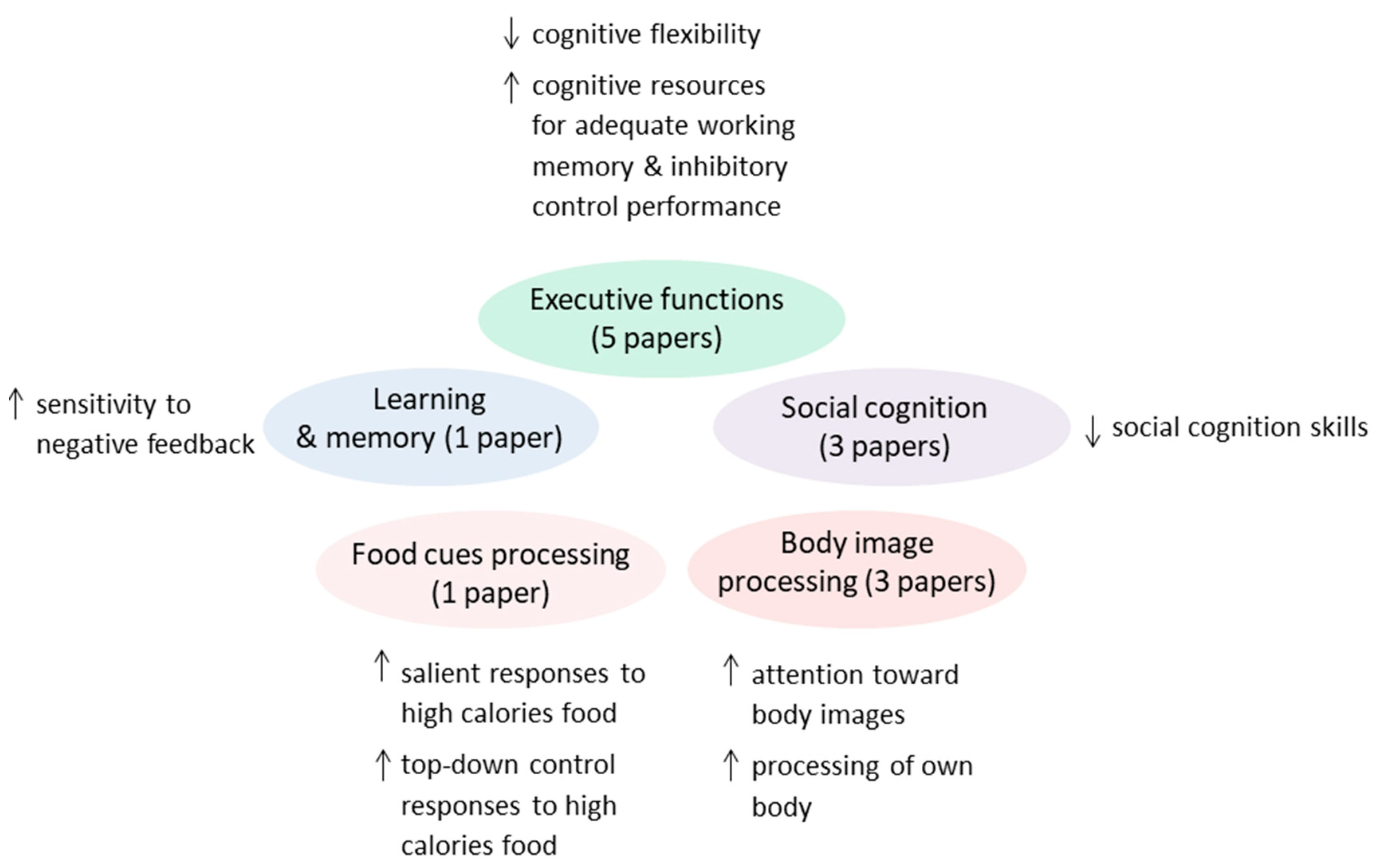
3.1. Executive Functions
3.2. Learning and Memory
3.3. Social Cognition
3.4. ED-Related Stimuli
3.5. Quality Assessment of the Studies
4. Discussion
4.1. Executive Functions in Healthy and Anorectic Adolescents
4.2. Learning and Reward Processing
4.3. Social Cognition
4.4. ED-Related Stimuli
4.5. Pathogenic Model
4.6. Implications for Treatment
4.7. Limitation of Current Research
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Campbell, K.; Peebles, R. Eating disorders in children and adolescents: State of the art review. Pediatrics 2014, 134, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Jagielska, G.; Kacperska, I. Outcome, comorbidity and prognosis in anorexia nervosa. Psychiatr. Polska 2017, 51, 205–218. [Google Scholar] [CrossRef] [PubMed]
- Berends, T.; van Meijel, B.; Nugteren, W.; Deen, M.; Danner, U.N.; Hoek, H.W.; van Elburg, A.A. Rate, timing and predictors of relapse in patients with anorexia nervosa following a relapse prevention program: A cohort study. BMC Psychiatry 2016, 16, 316. [Google Scholar] [CrossRef] [PubMed]
- Khalsa, S.S.; Portnoff, L.C.; McCurdy-McKinnon, D.; Feusner, J.D. What happens after treatment? A systematic review of relapse, remission, and recovery in anorexia nervosa. J. Eat. Disord. 2017, 5, 20. [Google Scholar] [CrossRef] [PubMed]
- Legenbauer, T.M.; Meule, A. Challenges in the Treatment of Adolescent Anorexia Nervosa—Is Enhanced Cognitive Behavior Therapy The Answer? Front. Psychiatry 2015, 6, 148. [Google Scholar] [CrossRef] [PubMed]
- Marucci, S.; Ragione, L.D.; De Iaco, G.; Mococci, T.; Vicini, M.; Guastamacchia, E.; Triggiani, V. Anorexia Nervosa and Comorbid Psychopathology. Endocr. Metab. Immune Disord. Drug Targets 2018, 18, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Meczekalski, B.; Podfigurna-Stopa, A.; Katulski, K. Long-term consequences of anorexia nervosa. Maturitas 2013, 75, 215–220. [Google Scholar] [CrossRef]
- Gorrell, S.; Loeb, K.; Le Grange, D. Family-based Treatment of Eating Disorders: A Narrative Review. Psychiatr. Clin. N. Am. 2019, 42, 193–204. [Google Scholar] [CrossRef]
- Le Grange, D.; Hughes, E.K.; Court, A.; Yeo, M.; Crosby, R.D.; Sawyer, S.M. Randomized Clinical Trial of Parent-Focused Treatment and Family-Based Treatment for Adolescent Anorexia Nervosa. J. Am. Acad. Child. Adolesc. Psychiatry 2016, 55, 683–692. [Google Scholar] [CrossRef]
- Murray, S.B.; Loeb, K.L.; Le Grange, D. Treatment outcome reporting in anorexia nervosa: Time for a paradigm shift? J. Eat. Disord. 2018, 6, 10. [Google Scholar] [CrossRef]
- Le Grange, D.; Lock, J.; Accurso, E.C.; Agras, W.S.; Darcy, A.; Forsberg, S.; Bryson, S.W. Relapse from remission at two-to four-year follow-up in two treatments for adolescent anorexia nervosa. J. Am. Acad Child. Adolesc. Psychiatry 2014, 53, 1162–1167. [Google Scholar] [CrossRef] [PubMed]
- Agras, W.S. Cognitive Behavior Therapy for the Eating Disorders. Psychiatr. Clin. N. Am. 2019, 42, 169–179. [Google Scholar] [CrossRef] [PubMed]
- Vijayakumar, N.; Op de Macks, Z.; Shirtcliff, E.A.; Pfeifer, J.H. Puberty and the human brain: Insights into adolescent development. Neurosci. Biobehav. Rev. 2018, 92, 417–436. [Google Scholar] [CrossRef] [PubMed]
- Teeuw, J.; Brouwer, R.M.; Koenis, M.M.G.; Swagerman, S.C.; Boomsma, D.I.; Hulshoff Pol, H.E. Genetic Influences on the Development of Cerebral Cortical Thickness During Childhood and Adolescence in a Dutch Longitudinal Twin Sample: The Brainscale Study. Cereb. Cortex 2019, 29, 978–993. [Google Scholar] [CrossRef] [PubMed]
- Morgan, S.E.; White, S.R.; Bullmore, E.T.; Vertes, P.E. A Network Neuroscience Approach to Typical and Atypical Brain Development. Biol. Psychiatry Cogn. Neurosci. Neuroimaging 2018, 3, 754–766. [Google Scholar] [CrossRef] [PubMed]
- Herting, M.M.; Johnson, C.; Mills, K.L.; Vijayakumar, N.; Dennison, M.; Liu, C.; Goddings, A.L.; Dahl, R.E.; Sowell, E.R.; Whittle, S.; et al. Development of subcortical volumes across adolescence in males and females: A multisample study of longitudinal changes. Neuroimage 2018, 172, 194–205. [Google Scholar] [CrossRef]
- Wierenga, L.; Langen, M.; Ambrosino, S.; van Dijk, S.; Oranje, B.; Durston, S. Typical development of basal ganglia, hippocampus, amygdala and cerebellum from age 7 to 24. Neuroimage 2014, 96, 67–72. [Google Scholar] [CrossRef]
- Narvacan, K.; Treit, S.; Camicioli, R.; Martin, W.; Beaulieu, C. Evolution of deep gray matter volume across the human lifespan. Hum. Brain Mapp. 2017, 38, 3771–3790. [Google Scholar] [CrossRef]
- Darcey, V.L.; McQuaid, G.A.; Fishbein, D.H.; VanMeter, J.W. Dietary Long-Chain Omega-3 Fatty Acids Are Related to Impulse Control and Anterior Cingulate Function in Adolescents. Front. Neurosci 2018, 12, 1012. [Google Scholar] [CrossRef]
- Herd, T.; Li, M.; Maciejewski, D.; Lee, J.; Deater-Deckard, K.; King-Casas, B.; Kim-Spoon, J. Inhibitory Control Mediates the Association between Perceived Stress and Secure Relationship Quality. Front. Psychol. 2018, 9, 217. [Google Scholar] [CrossRef]
- Qu, Y.; Pomerantz, E.M.; McCormick, E.; Telzer, E.H. Youth’s Conceptions of Adolescence Predict Longitudinal Changes in Prefrontal Cortex Activation and Risk Taking During Adolescence. Child. Dev. 2018, 89, 773–783. [Google Scholar] [CrossRef] [PubMed]
- Moher, D.; Liberati, A.; Tetzlaff, J.; Altman, D.G.; Group, P. Preferred reporting items for systematic reviews and meta-analyses: The PRISMA statement. BMJ 2009, 339, b2535. [Google Scholar] [CrossRef] [PubMed]
- American Psychiatric Association. Diagnostic and Statistical Manual of Mental Disorders, 5th ed.; American Psychiatric Association Publishing: Washington, DC, USA, 2013. [Google Scholar]
- Daniel, R.; Pollmann, S. A universal role of the ventral striatum in reward-based learning: Evidence from human studies. Neurobiol. Learn. Mem. 2014, 114, 90–100. [Google Scholar] [CrossRef] [PubMed]
- Soltani, A.; Izquierdo, A. Adaptive learning under expected and unexpected uncertainty. Nat. Rev. Neurosci. 2019. [Google Scholar] [CrossRef] [PubMed]
- Frank, G.K.W.; Favaro, A.; Marsh, R.; Ehrlich, S.; Lawson, E.A. Toward valid and reliable brain imaging results in eating disorders. Int. J. Eat. Disord. 2018, 51, 250–261. [Google Scholar] [CrossRef] [PubMed]
- Thirion, B.; Pinel, P.; Meriaux, S.; Roche, A.; Dehaene, S.; Poline, J.B. Analysis of a large fMRI cohort: Statistical and methodological issues for group analyses. Neuroimage 2007, 35, 105–120. [Google Scholar] [CrossRef] [PubMed]
- Firk, C.; Mainz, V.; Schulte-Ruether, M.; Fink, G.; Herpertz-Dahlmann, B.; Konrad, K. Implicit sequence learning in juvenile anorexia nervosa: Neural mechanisms and the impact of starvation. J. Child. Psychol. Psychiatry 2015, 56, 1168–1176. [Google Scholar] [CrossRef]
- Hildebrandt, T.; Schulz, K.; Schiller, D.; Heywood, A.; Goodman, W.; Sysko, R. Evidence of prefrontal hyperactivation to food-cue reversal learning in adolescents with anorexia nervosa. Behav. Res. 2018, 111, 36–43. [Google Scholar] [CrossRef]
- Castro-Fornieles, J.; Caldu, X.; Andres-Perpina, S.; Lazaro, L.; Bargallo, N.; Falcon, C.; Plana, M.T.; Junque, C. A cross-sectional and follow-up functional MRI study with a working memory task in adolescent anorexia nervosa. Neuropsychologia 2010, 48, 4111–4116. [Google Scholar] [CrossRef]
- Lock, J.; Garrett, A.; Beenhakker, J.; Reiss, A.L. Aberrant brain activation during a response inhibition task in adolescent eating disorder subtypes. Am. J. Psychiatry 2011, 168, 55–64. [Google Scholar] [CrossRef]
- Wierenga, C.; Bischoff-Grethe, A.; Melrose, A.J.; Grenesko-Stevens, E.; Irvine, Z.; Wagner, A.; Simmons, A.; Matthews, S.; Yau, W.Y.; Fennema-Notestine, C.; et al. Altered BOLD response during inhibitory and error processing in adolescents with anorexia nervosa. PLoS ONE 2014, 9, e92017. [Google Scholar] [CrossRef] [PubMed]
- Bischoff-Grethe, A.; McCurdy, D.; Grenesko-Stevens, E.; Irvine, L.E.; Wagner, A.; Yau, W.Y.; Fennema-Notestine, C.; Wierenga, C.E.; Fudge, J.L.; Delgado, M.R.; et al. Altered brain response to reward and punishment in adolescents with Anorexia nervosa. Psychiatry Res. 2013, 214, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Horndasch, S.; Roesch, J.; Forster, C.; Dorfler, A.; Lindsiepe, S.; Heinrich, H.; Graap, H.; Moll, G.H.; Kratz, O. Neural processing of food and emotional stimuli in adolescent and adult anorexia nervosa patients. PLoS ONE 2018, 13, e0191059. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Ruther, M.; Mainz, V.; Fink, G.R.; Herpertz-Dahlmann, B.; Konrad, K. Theory of mind and the brain in anorexia nervosa: Relation to treatment outcome. J. Am. Acad. Child. Adolesc. Psychiatry 2012, 51, 832–841.e811. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Harper, J.A.; Van Enkevort, E.A.; Latimer, K.; Kelley, U.; McAdams, C.J. Neural activations are related to body-shape, anxiety, and outcomes in adolescent anorexia nervosa. J. Psychiatr. Res. 2017, 87, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Fladung, A.K.; Schulze, U.M.; Scholl, F.; Bauer, K.; Gron, G. Role of the ventral striatum in developing anorexia nervosa. Transl. Psychiatry 2013, 3, e315. [Google Scholar] [CrossRef] [PubMed]
- Seeger, G.; Braus, D.F.; Ruf, M.; Goldberger, U.; Schmidt, M.H. Body image distortion reveals amygdala activation in patients with anorexia nervosa—A functional magnetic resonance imaging study. Neurosci. Lett. 2002, 326, 25–28. [Google Scholar] [CrossRef]
- Wagner, A.; Ruf, M.; Braus, D.F.; Schmidt, M.H. Neuronal activity changes and body image distortion in anorexia nervosa. Neuroreport 2003, 14, 2193–2197. [Google Scholar] [CrossRef]
- Raznahan, A.; Shaw, P.W.; Lerch, J.P.; Clasen, L.S.; Greenstein, D.; Berman, R.; Pipitone, J.; Chakravarty, M.M.; Giedd, J.N. Longitudinal four-dimensional mapping of subcortical anatomy in human development. Proc. Natl. Acad. Sci. USA 2014, 111, 1592–1597. [Google Scholar] [CrossRef]
- Yang, J.; Li, P. Brain networks of explicit and implicit learning. PLoS ONE 2012, 7, e42993. [Google Scholar] [CrossRef]
- Darki, F.; Klingberg, T. The role of fronto-parietal and fronto-striatal networks in the development of working memory: A longitudinal study. Cereb. Cortex 2015, 25, 1587–1595. [Google Scholar] [CrossRef] [PubMed]
- Simmonds, D.J.; Hallquist, M.N.; Luna, B. Protracted development of executive and mnemonic brain systems underlying working memory in adolescence: A longitudinal fMRI study. Neuroimage 2017, 157, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Baker, S.T.; Lubman, D.I.; Yucel, M.; Allen, N.B.; Whittle, S.; Fulcher, B.D.; Zalesky, A.; Fornito, A. Developmental Changes in Brain Network Hub Connectivity in Late Adolescence. J. Neurosci. 2015, 35, 9078–9087. [Google Scholar] [CrossRef] [PubMed]
- Anandakumar, J.; Mills, K.L.; Earl, E.A.; Irwin, L.; Miranda-Dominguez, O.; Demeter, D.V.; Walton-Weston, A.; Karalunas, S.; Nigg, J.; Fair, D.A. Individual differences in functional brain connectivity predict temporal discounting preference in the transition to adolescence. Dev. Cogn. Neurosci. 2018, 34, 101–113. [Google Scholar] [CrossRef] [PubMed]
- Telzer, E.H.; Fuligni, A.J.; Lieberman, M.D.; Miernicki, M.E.; Galvan, A. The quality of adolescents’ peer relationships modulates neural sensitivity to risk taking. Soc. Cogn. Affect. Neurosci. 2015, 10, 389–398. [Google Scholar] [CrossRef] [PubMed]
- Achterberg, M.; Peper, J.S.; van Duijvenvoorde, A.C.; Mandl, R.C.; Crone, E.A. Frontostriatal White Matter Integrity Predicts Development of Delay of Gratification: A Longitudinal Study. J. Neurosci. 2016, 36, 1954–1961. [Google Scholar] [CrossRef]
- Peters, S.; Crone, E.A. Increased striatal activity in adolescence benefits learning. Nat. Commun. 2017, 8, 1983. [Google Scholar] [CrossRef] [PubMed]
- Schreuders, E.; Braams, B.R.; Blankenstein, N.E.; Peper, J.S.; Guroglu, B.; Crone, E.A. Contributions of Reward Sensitivity to Ventral Striatum Activity Across Adolescence and Early Adulthood. Child. Dev. 2018, 89, 797–810. [Google Scholar] [CrossRef] [PubMed]
- Braams, B.R.; van Duijvenvoorde, A.C.; Peper, J.S.; Crone, E.A. Longitudinal changes in adolescent risk-taking: A comprehensive study of neural responses to rewards, pubertal development, and risk-taking behavior. J. Neurosci. 2015, 35, 7226–7238. [Google Scholar] [CrossRef]
- Misra, M.; Klibanski, A. Endocrine consequences of anorexia nervosa. Lancet Diabetes Endocrinol. 2014, 2, 581–592. [Google Scholar] [CrossRef]
- Misra, M.; Klibanski, A. Anorexia Nervosa and Its Associated Endocrinopathy in Young People. Horm. Res. Paediatr. 2016, 85, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Casement, M.D.; Guyer, A.E.; Hipwell, A.E.; McAloon, R.L.; Hoffmann, A.M.; Keenan, K.E.; Forbes, E.E. Girls’ challenging social experiences in early adolescence predict neural response to rewards and depressive symptoms. Dev. Cogn. Neurosci. 2014, 8, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Tachi, T. Family environment in eating disorders: A study of the familiar factors influencing the onset and course of eating disorders. Seishin Shinkeigaku Zasshi 1999, 101, 427–445. [Google Scholar] [PubMed]
- Campos, L.K.; Sampaio, A.B.; Garcia, C., Jr.; Magdaleno, R., Jr.; Battistoni, M.M.; Turato, E.R. Psychological characteristics of mothers of patients with anorexia nervosa: Implications for treatment and prognosis. Trends Psychiatry Psychother. 2012, 34, 13–18. [Google Scholar] [CrossRef] [PubMed]
- Copeland, W.E.; Bulik, C.M.; Zucker, N.; Wolke, D.; Lereya, S.T.; Costello, E.J. Does childhood bullying predict eating disorder symptoms? A prospective, longitudinal analysis. Int. J. Eat. Disord. 2015, 48, 1141–1149. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, J.H.; Kahn, L.E.; Merchant, J.S.; Peake, S.J.; Veroude, K.; Masten, C.L.; Lieberman, M.D.; Mazziotta, J.C.; Dapretto, M. Longitudinal change in the neural bases of adolescent social self-evaluations: Effects of age and pubertal development. J. Neurosci. 2013, 33, 7415–7419. [Google Scholar] [CrossRef] [PubMed]
- Beadle, J.N.; Paradiso, S.; Salerno, A.; McCormick, L.M. Alexithymia, emotional empathy, and self-regulation in anorexia nervosa. Ann. Clin. Psychiatry 2013, 25, 107–120. [Google Scholar]
- Guyer, A.E.; Silk, J.S.; Nelson, E.E. The neurobiology of the emotional adolescent: From the inside out. Neurosci. Biobehav. Rev. 2016, 70, 74–85. [Google Scholar] [CrossRef]
- McRae, K.; Gross, J.J.; Weber, J.; Robertson, E.R.; Sokol-Hessner, P.; Ray, R.D.; Gabrieli, J.D.; Ochsner, K.N. The development of emotion regulation: An fMRI study of cognitive reappraisal in children, adolescents and young adults. Soc. Cogn. Affect. Neurosci. 2012, 7, 11–22. [Google Scholar] [CrossRef]
- Bolling, D.Z.; Pelphrey, K.A.; Vander Wyk, B.C. Unlike adults, children and adolescents show predominantly increased neural activation to social exclusion by members of the opposite gender. Soc. Neurosci. 2016, 11, 475–486. [Google Scholar] [CrossRef]
- Vijayakumar, N.; Cheng, T.W.; Pfeifer, J.H. Neural correlates of social exclusion across ages: A coordinate-based meta-analysis of functional MRI studies. Neuroimage 2017, 153, 359–368. [Google Scholar] [CrossRef] [PubMed]
- Sylvester, C.M.; Whalen, D.J.; Belden, A.C.; Sanchez, S.L.; Luby, J.L.; Barch, D.M. Shyness and Trajectories of Functional Network Connectivity Over Early Adolescence. Child. Dev. 2018, 89, 734–745. [Google Scholar] [CrossRef] [PubMed]
- Weissman, D.G.; Guyer, A.E.; Ferrer, E.; Robins, R.W.; Hastings, P.D. Adolescents’ brain-autonomic coupling during emotion processing. Neuroimage 2018, 183, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Haller, S.P.; Kircanski, K.; Stoddard, J.; White, L.K.; Chen, G.; Sharif-Askary, B.; Zhang, S.; Towbin, K.E.; Pine, D.S.; Leibenluft, E.; et al. Reliability of neural activation and connectivity during implicit face emotion processing in youth. Dev. Cogn. Neurosci. 2018, 31, 67–73. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, J.H.; Blakemore, S.J. Adolescent social cognitive and affective neuroscience: Past, present, and future. Soc. Cogn. Affect. Neurosci. 2012, 7, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Turpyn, C.C.; Poon, J.A.; Ross, C.E.; Thompson, J.C.; Chaplin, T.M. Associations Between Parent Emotional Arousal and Regulation and Adolescents’ Affective Brain Response. Soc. Dev. 2018, 27, 3–18. [Google Scholar] [CrossRef] [PubMed]
- Frank, S.; Kullmann, S.; Veit, R. Food related processes in the insular cortex. Front. Hum. Neurosci. 2013, 7, 499. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, N.R.; Pfeifer, J.H. Age-related changes in reappraisal of appetitive cravings during adolescence. Neuroimage 2015, 108, 173–181. [Google Scholar] [CrossRef]
- Ernst, M.; Luciana, M. Neuroimaging of the dopamine/reward system in adolescent drug use. CNS Spectr. 2015, 20, 427–441. [Google Scholar] [CrossRef]
- Ernst, M.; Mueller, S.C. The adolescent brain: Insights from functional neuroimaging research. Dev. Neurobiol. 2008, 68, 729–743. [Google Scholar] [CrossRef]
- Morita, T.; Saito, D.N.; Ban, M.; Shimada, K.; Okamoto, Y.; Kosaka, H.; Okazawa, H.; Asada, M.; Naito, E. Self-Face Recognition Begins to Share Active Region in Right Inferior Parietal Lobule with Proprioceptive Illusion During Adolescence. Cereb. Cortex 2018, 28, 1532–1548. [Google Scholar] [CrossRef] [PubMed]
- Grzelak, T.; Dutkiewicz, A.; Paszynska, E.; Dmitrzak-Weglarz, M.; Slopien, A.; Tyszkiewicz-Nwafor, M. Neurobiochemical and psychological factors influencing the eating behaviors and attitudes in anorexia nervosa. J. Physiol. Biochem. 2017, 73, 297–305. [Google Scholar] [CrossRef] [PubMed]
- Munro, C.; Randell, L.; Lawrie, S.M. An Integrative Bio-Psycho-Social Theory of Anorexia Nervosa. Clin. Psychol. Psychother. 2017, 24, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Coniglio, K.A.; Becker, K.R.; Franko, D.L.; Zayas, L.V.; Plessow, F.; Eddy, K.T.; Thomas, J.J. Won’t stop or can’t stop? Food restriction as a habitual behavior among individuals with anorexia nervosa or atypical anorexia nervosa. Eat. Behav. 2017, 26, 144–147. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study | Participants | fMRI Paradigm | Main Findings | Main Conclusions |
|---|---|---|---|---|
| EXECUTIVE FUNCTIONS | ||||
| Cognitive flexibility | ||||
| Firk, 2015 [28] | AN n = 19 (females), subtype nd. Mean age 15.9 yrs (±1.5). Controls n = 20 (females). Mean age 15.9 yrs (±1.9). | Serial reaction time (SRT) Implicit learning | SRT performance was impaired in AN patients. AN patients also showed lower activity in the ventral anterior and ventral lateral thalamic nuclei compared with controls. | The impairment in cognitive flexibility in AN patients might contribute to the persistence of habitual behaviors (such as restricting the caloric intake) in these individuals. |
| Hildebrandt, 2018 [29] | AN n = 15 (females), subtype nd. Mean age nd. Controls n = 14 (females). Mean age nd. | Reversal learning | AN and controls did not differ on expectancy ratings (performance) on the task. During the association (learning) phase, AN had higher activity in the DLPFC, IFG, and IOG compared with controls. During the cue-reversal learning phase, AN showed greater activation in the VLPFC, DLPFC, IFG, and MOG compared with controls. | AN patients are as proficient as controls in reversing a stimulus-outcome learned association, however they require a greater engagement of top-down, inhibitory control regions. Thus, a higher cognitive control is required for AN patients compared with controls to achieve the same cognitive flexibility. |
| Working memory | ||||
| * Castro-Fornieles, 2010 [30] | AN n = 14 (12 females), subtype nd. Mean age 15.0 yrs (±1.7). Controls n = 14 (7 females). Mean age 15.4 yrs (±0.1). | 1-back task | AN and controls did not differ in terms of performance (number of errors), however AN showed hyperactivity in the superior temporal gyrus during the task. They also had a trend for higher activity in several temporal and parietal areas, correlating positively with depressive symptomatology and negatively with BMI. Brain activity normalized weight restoration. | AN patients need more cognitive resources to achieve the same level of performance as controls. The cognitive load required is higher for individuals with higher depressive symptoms and lower BMI. Treatment and weight restoration can rescue cognitive abilities. |
| Inhibitory control | ||||
| Lock, 2011 [31] | AN n = 14 (females), restrictive subtype. Mean age 15.0 yrs (±1.7). Controls n = 13 (females). Mean age 15.9 yrs (±1.3). | Go-NoGo task | AN patients and controls did not differ on task accuracy. AN patients showed a positive correlation with percent correctly inhibited trials in the inferior parietal cortex, precuneus, and PCC. | In AN patients, successful response inhibition is associated with greater recruitment of brain regions underlying visual attention and visual working memory. |
| Wierenga, 2014 [32] | AN n = 11 (females), restrictive type. Mean age 16.0 yrs (±2.0). Controls n = 12 (females). Mean age 14.9 yrs (±1.8). | Stop signal task (SST) | AN patients had lower post-error slowing. AN patients had lower activity in the dorsal anterior ACC, MFG, and PCC during hard trials compared with controls. Patients also had lower activity in MFG and PCC during error (failed inhibit) processing trials, compared with controls. | AN patients have impaired representation of task difficulty, reflecting impaired cognitive flexibility. Nonetheless, they seem to require less resources for error monitoring. |
| LEARNING AND MEMORY | ||||
| Reward processing | ||||
| Bischoff-Grethe, 2013 [33] | AN n = 10 (females), restrictive subtype. Mean age 16.2 yrs (±1.8). Controls n = 12 (females). Mean age 15.4 yrs (±1.6) | Monetary guessing task | AN showed normal responses to reward in the anterior limbic system, but greater response to punishment compared with controls in the posterior caudate and in the cognitive cingulate cortex. Controls were more responsive to reward in the anterior putamen and motor cingulate cortex, compared with AN. | During action-outcome learning, AN patients have normal reward expectancies; however they are highly sensitive to punishment (negative feedback). This impairs their ability to appropriately proportion reward and punishment in order to learn from experience. |
| SOCIAL COGNITION | ||||
| Emotional processing | ||||
| Horndasch, 2018 [34] | AN n = 15 (females), subtype nd. Mean age 16.4 yrs (±1.4). Controls n = 18 (females). Mean age 15.9 yrs (±2.1). | Viewing and rating pictures of high-calorie food, low-calorie food, negative, neural, positive stimuli | No differences were found in the ratings of emotional stimuli. Controls showed greater activity compared with AN patients in the cerebellum, ACC, striatum, and inferior frontal gyrus for negative stimuli; in the cerebellum for neutral stimuli; in the cerebellum, striatum, precuneus, ACC, inferior frontal gyrus and hippocampus for positive stimuli. AN patients had higher activity than controls when viewing neutral and positive stimuli in the medial PFC. | AN patients showed lower processing of all emotional stimuli with some specific regions involved in positive picture processing, possibly reflecting impaired ability to experience pleasure by daily natural reinforcers. |
| Theory of the Mind | ||||
| * Schulte-Ruther, 2012 [35] | AN n = 19 (females), 13 restrictive subtype. Mean age 15.7 yrs (±1.5). Controls n = 21 (females). Mean age 15.8 yrs (±1.9). | Social attribution task (SAT) | AN patients and controls did not differ in the attribution of social relations. At baseline, AN patients had lower activity in the STG, MTG, and TP compared with controls when viewing social vs. non-social videos. After weight restoration, patients still had lower activity compared with controls in the MTG and TP. Lower baseline activity correlated with worse treatment outcome. | AN patients show impaired social functioning and social mentalization abilities, partially persisting after treatment and weight restoration. Poorer social cognition correlates with worse treatment outcome. |
| Social evaluation | ||||
| † Xu, 2017 [36] | AN n = 24 (females), 19 restrictive subtype. Mean age 16.4 yrs (±2.0). Controls n = 18 (females). Mean age 16.1 yrs (±2.3). | Social Identity-V2 task. Reading and responding (agree or disagree) to statements related to thinking about oneself, one’s friend, or what one’s friend thinks of them. | AN patients and controls did not differ on neural activity. Within patients, PCC activity was higher in response to friend-relative-to-self evaluations in recovered patients compared with those who remained ill. MPFC-dACC activity correlated with concerns about body shape, and MPFC-Cing activity correlated with anxiety levels. | Differences in social evaluations may contribute to both anxiety and body shape concerns in AN, and might have clinical predictive value. |
| ED-RELATED STIMULI | ||||
| Body image perception | ||||
| Fladung, 2013 [37] | AN n = 13 (females), 10 restrictive subtypes. Mean age 16.0 yrs (±1.1). Controls n = 14 (females). Mean age 16.6 yrs (±1.1). | Viewing and rating images of underweight, normal weight, and overweight female bodies. | AN patients rated underweight stimuli as more satisfying compared with controls. Controls had striatal higher activity compared to patients when processing normal-weight stimuli, while patients had higher striatal activity when processing underweight stimuli. | AN might engage the same circuitry involved in addiction disorders, and might thus be considered as a starvation dependence. |
| Seeger, 2002 [38] | AN n = 3 (females), subtype nd. Mean age 17.0 yrs (±0.5). Controls n = 3 (females). Mean age 17.5 yrs (±0.5). | Viewing: (1) distorted images of own body; (2) distorted imagesof another woman’s body;(3) scrambled images with mixed colors composed of own body images. | AN patients showed greater activity in the brainstem, right amygdala, and right gyrus fusiformis when viewing distorted own body image versus an average of non-target and neutral pictures. | AN patients show aversive responses when confronted with distorted images of own body shape. |
| Wagner, 2003 [39] | AN n = 13 (females), 10 restrictive subtype. Mean age 15.3 yrs (±1.4). Controls n = 10 (females). Mean age 15.1 yrs (±1.9). | Viewing: (1) distorted images of own body; (2) distorted images of another woman’s body; (3) scrambled images with mixed colors composed of own body images. | The PFC activity was higher for own body than for other women’s body or neutral pictures in controls, while it was higher in patients for both own body or other women’s body. AN patients had higher activity in the IPL when viewing own body compared with other women’s body or abstract shapes, while no differences were observed in controls. | AN patients might have an unspecific greater attention toward body stimuli, plus a specific visuo-spatial processing of own body shape. |
| Food stimuli | ||||
| Horndasch, 2018 [34] | AN n = 15 (females), subtype nd. Mean age 16.4 yrs (±1.4). Controls n = 18 (females). Mean age 15.9 yrs (±2.1). | Viewing and rating pictures of high-calorie food, low-calorie food, negative, neural, positive stimuli | AN patients gave lower ratings to high calorie foods compared with controls, while no differences were found in the ratings of low calorie foods. High-calorie foods elicited stronger IFG, medial prefrontal gyrus, and anterior insula activation in AN patients, but lower activity in the cerebellum compared with controls. For low-calorie stimuli, controls showed higher activity in the right cerebellum, and lower activity in the left cerebellar, medial PFC and parietal lobe compared with AN patients. | AN patients showed hyperactivity of the bottom-up and top-down areas in response to food. |
| Paper | Development, Demographic Data, and Illness State (0–13.5) | Effects of Exercise, Hydration Status, Binge Eating and Purging, and Malnutrition (0–16.25) | Stage of Treatment (0–6.0) | Hormonal Effects (0–9.25) | Comorbidity and Medication (0–10.0) | Technical and Statistical Considerations, and Study Design (0–15.25) | TOT (0–70.0) |
|---|---|---|---|---|---|---|---|
| Bischoff-Grethe, 2013 | 7.75 | 7 | 1.5 | 3 | 6 | 10.5 | 35.75 |
| Castro-Fornieles, 2010 | 7.5 | 11.5 | 4 | 3 | 6.5 | 3 | 35.50 |
| Firk, 2015 | 3 | 4 | 4 | 3 | 10 | 7.5 | 31.50 |
| Fladung, 2013 | 6 | 4.5 | 0 | 3 | 0 | 4.5 | 18.00 |
| Hildebrandt, 2018 | 0 | 0 | 0 | 3 | 5 | 4.5 | 12.50 |
| Horndasch, 2018 | 6 | 7 | 2.5 | 3 | 6 | 4.5 | 29.00 |
| Lock, 2011 | 9.25 | 6 | 1.5 | 6 | 3 | 7.5 | 33.25 |
| Schulte-Ruther, 2012 | 3 | 6.5 | 4 | 3 | 6 | 9 | 31.50 |
| Seeger, 2012 | 3 | 4 | 4 | 3 | 10 | 6 | 30.00 |
| Wagner, 2003 | 3 | 5.5 | 4 | 3 | 8.5 | 6 | 30.00 |
| Wierenga, 2014 | 6.25 | 10 | 4 | 3 | 3 | 10.5 | 36.75 |
| Xu, 2017 | 6.25 | 3.5 | 4 | 6 | 3 | 7.5 | 30.25 |
| Mean score | 5.1 | 5.8 | 2.8 | 3.5 | 5.6 | 6.75 | 29.50 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Olivo, G.; Gaudio, S.; Schiöth, H.B. Brain and Cognitive Development in Adolescents with Anorexia Nervosa: A Systematic Review of fMRI Studies. Nutrients 2019, 11, 1907. https://doi.org/10.3390/nu11081907
Olivo G, Gaudio S, Schiöth HB. Brain and Cognitive Development in Adolescents with Anorexia Nervosa: A Systematic Review of fMRI Studies. Nutrients. 2019; 11(8):1907. https://doi.org/10.3390/nu11081907
Chicago/Turabian StyleOlivo, Gaia, Santino Gaudio, and Helgi B. Schiöth. 2019. "Brain and Cognitive Development in Adolescents with Anorexia Nervosa: A Systematic Review of fMRI Studies" Nutrients 11, no. 8: 1907. https://doi.org/10.3390/nu11081907
APA StyleOlivo, G., Gaudio, S., & Schiöth, H. B. (2019). Brain and Cognitive Development in Adolescents with Anorexia Nervosa: A Systematic Review of fMRI Studies. Nutrients, 11(8), 1907. https://doi.org/10.3390/nu11081907