Cow’s Milk Allergy: Immunomodulation by Dietary Intervention



 ,
,  and
and 
Abstract
1. Introduction
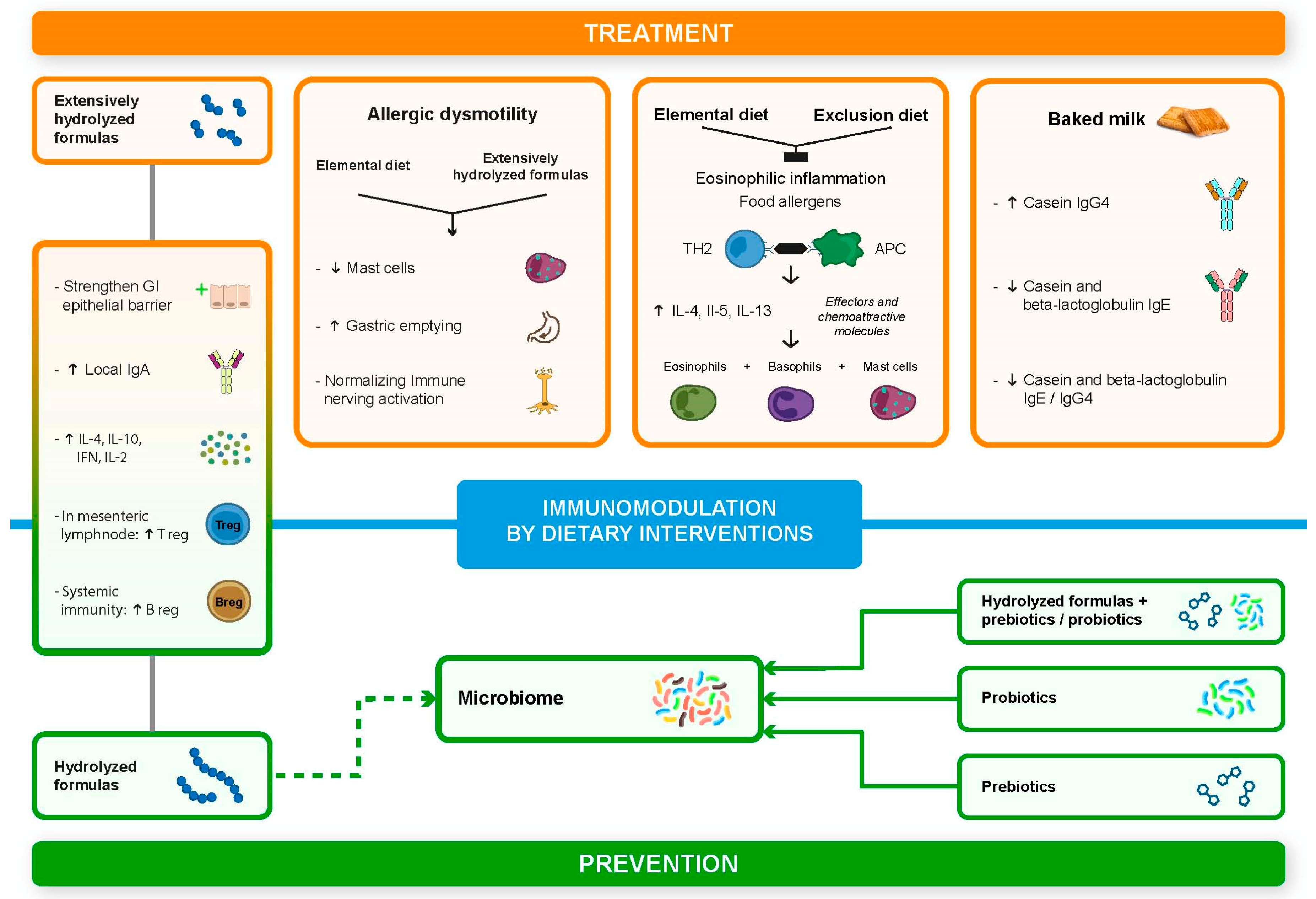
2. Cow’s Milk Allergy and Allergic Dysmotility: A Pleiomorphic Disorder
3. Eosinophilic Esophagitis: Insights on Pathogenetic Mechanisms and Dietary Immunomodulation
4. Immune Modulation by Hydrolysate Proteins
5. Baked Milk: A Possible Form of Oral Immunotherapy?
6. Gut Microbiota in Perinatal Period and Its Relationship with Immune Function and Allergy Development
7. Prebiotics and CMA
8. Probiotics and CM
9. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Sicherer, S.H. Epidemiology of food allergy. J. Allergy Clin. Immunol. 2011, 127, 594–602. [Google Scholar] [CrossRef] [PubMed]
- Schappi, M.G.; Borrelli, O. Mast cell-nerve interactions in children with functional dyspepsia. J. Pediatr. Gastroenterol. Nutr. 2008, 47, 472–480. [Google Scholar] [CrossRef] [PubMed]
- Molina-Infante, J.; Arias, A. Four-food group elimination diet for adult eosinophilic esophagitis: A prospective multicenter study. J. Allergy Clin. Immunol. 2014, 134, 1093–1099.e1. [Google Scholar] [CrossRef] [PubMed]
- Bougle, D.; Bouhallab, S. Dietary bioactive peptides: Human studies. Crit. Rev. Food Sci. Nutr. 2017, 57, 335–343. [Google Scholar] [CrossRef] [PubMed]
- Esmaeilzadeh, H.; Alyasin, S. The effect of baked milk on accelerating unheated cow’s milk tolerance: A control randomized clinical trial. Pediatr. Allergy Immunol. 2018, 29, 747–753. [Google Scholar] [CrossRef]
- Huang, F.; Nowak-Wegrzyn, A. Extensively heated milk and egg as oral immunotherapy. Curr. Opin. Allergy Clin. Immunol. 2012, 12, 283–292. [Google Scholar] [CrossRef]
- Berni Canani, R.; Gilbert, J.A. The role of the commensal microbiota in the regulation of tolerance to dietary allergens. Curr. Opin. Allergy Clin. Immunol. 2015, 15, 243–249. [Google Scholar] [CrossRef]
- Plunkett, C.H.; Nagler, C.R. The Influence of the Microbiome on Allergic Sensitization to Food. J. Immunol. 2017, 198, 581–589. [Google Scholar] [CrossRef]
- Dupont, C. Diagnosis of cow’s milk allergy in children: Determining the gold standard? Expert Rev. Clin. Immunol. 2014, 10, 257–267. [Google Scholar] [CrossRef]
- Hill, D.J.; Hosking, C.S. The cow milk allergy complex: Overlapping disease profiles in infancy. Eur. J. Clin. Nutr. 1995, 49 (Suppl. 1), S1–S12. [Google Scholar]
- du Toit, G.; Meyer, R. Identifying and managing cow’s milk protein allergy. Arch. Dis. Child. Educ. Pract. Ed. 2010, 95, 134–144. [Google Scholar] [CrossRef] [PubMed]
- Heine, R.G. Gastroesophageal reflux disease, colic and constipation in infants with food allergy. Curr. Opin. Allergy Clin. Immunol. 2006, 6, 220–225. [Google Scholar] [CrossRef] [PubMed]
- Rosen, R.; Vandenplas, Y. Pediatric Gastroesophageal Reflux Clinical Practice Guidelines: Joint Recommendations of the North American Society for Pediatric Gastroenterology, Hepatology, and Nutrition and the European Society for Pediatric Gastroenterology, Hepatology, and Nutrition. J. Pediatr. Gastroenterol. Nutr. 2018, 66, 516–554. [Google Scholar] [CrossRef] [PubMed]
- Salvatore, S.; Vandenplas, Y. Gastroesophageal reflux and cow milk allergy: Is there a link? Pediatrics 2002, 110, 972–984. [Google Scholar] [CrossRef] [PubMed]
- Murch, S. Allergy and intestinal dysmotility—Evidence of genuine causal linkage? Curr. Opin. Gastroenterol. 2006, 22, 664–668. [Google Scholar] [CrossRef] [PubMed]
- Nakajima-Adachi, H.; Ebihara, A. Food antigen causes TH2-dependent enteropathy followed by tissue repair in T-cell receptor transgenic mice. J. Allergy Clin. Immunol. 2006, 117, 1125–1132. [Google Scholar] [CrossRef] [PubMed]
- Barbara, G.; Stanghellini, V. Functional gastrointestinal disorders and mast cells: Implications for therapy. Neurogastroenterol. Motil. 2006, 18, 6–17. [Google Scholar] [CrossRef]
- Jakate, S.; Demeo, M. Mastocytic enterocolitis: Increased mucosal mast cells in chronic intractable diarrhea. Arch. Pathol. Lab. Med. 2006, 130, 362–367. [Google Scholar]
- Ramsay, D.B.; Stephen, S. Mast cells in gastrointestinal disease. Gastroenterol. Hepatol. 2010, 6, 772–777. [Google Scholar]
- Feldman, M.J.; Morris, G.P. Mast cells mediate acid-induced augmentation of opossum esophageal blood flow via histamine and nitric oxide. Gastroenterology 1996, 110, 121–128. [Google Scholar] [CrossRef]
- Wu, S.V.; Yuan, P.Q. Identification and characterization of multiple corticotropin-releasing factor type 2 receptor isoforms in the rat esophagus. Endocrinology 2007, 148, 1675–1687. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.J.; Wang, K. Mast cell activation is involved in stress-induced epithelial barrier dysfunction in the esophagus. J. Dig. Dis. 2015, 16, 186–196. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.; Ding, X. Alterations of Mast Cells in the Esophageal Mucosa of the Patients with Non-Erosive Reflux Disease. Gastroenterol. Res. 2011, 4, 70–75. [Google Scholar] [CrossRef] [PubMed]
- Zhong, C.; Liu, K. Developing a diagnostic understanding of GERD phenotypes through the analysis of levels of mucosal injury, immune activation, and psychological comorbidity. Dis. Esophagus 2018, 31, doy039. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Gao, G. TRPA1 in mast cell activation-induced long-lasting mechanical hypersensitivity of vagal afferent C-fibers in guinea pig esophagus. Am. J. Physiol. Gastrointest. Liver Physiol. 2009, 297, G34–G42. [Google Scholar] [CrossRef] [PubMed]
- Yu, S.; Kollarik, M. Mast cell-mediated long-lasting increases in excitability of vagal C fibers in guinea pig esophagus. Am. J. Physiol. Gastrointest. Liver Physiol. 2007, 293, G850–G856. [Google Scholar] [CrossRef]
- Cenac, N.; Chin, A.C. PAR2 activation alters colonic paracellular permeability in mice via IFN-gamma-dependent and -independent pathways. J. Physiol. 2004, 558, 913–925. [Google Scholar] [CrossRef]
- Itoh, Y.; Sendo, T. Physiology and pathophysiology of proteinase-activated receptors (PARs): Role of tryptase/PAR-2 in vascular endothelial barrier function. J. Pharmacol. Sci. 2005, 97, 14–19. [Google Scholar] [CrossRef]
- Liu, H.; Miller, D.V. Proteinase-activated receptor-2 activation evokes oesophageal longitudinal smooth muscle contraction via a capsaicin-sensitive and neurokinin-2 receptor-dependent pathway. Neurogastroenterol. Motil. 2010, 22, 210–216, e67. [Google Scholar] [CrossRef]
- Kandulski, A.; Wex, T. Proteinase-activated receptor-2 in the pathogenesis of gastroesophageal reflux disease. Am. J. Gastroenterol. 2010, 105, 1934–1943. [Google Scholar] [CrossRef]
- De Boissieu, D.; Waguet, J.C. The atopy patch tests for detection of cow’s milk allergy with digestive symptoms. J. Pediatr. 2003, 142, 203–205. [Google Scholar] [CrossRef] [PubMed]
- Majamaa, H.; Moisio, P. Cow’s milk allergy: Diagnostic accuracy of skin prick and patch tests and specific IgE. Allergy 1999, 54, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Koletzko, S.; Niggemann, B. Diagnostic approach and management of cow’s-milk protein allergy in infants and children: ESPGHAN GI Committee practical guidelines. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 221–229. [Google Scholar] [CrossRef] [PubMed]
- Sampson, H.A.; Gerth van Wijk, R. Standardizing double-blind, placebo-controlled oral food challenges: American Academy of Allergy, Asthma & Immunology-European Academy of Allergy and Clinical Immunology PRACTALL consensus report. J. Allergy Clin. Immunol. 2012, 130, 1260–1274. [Google Scholar] [PubMed]
- Giovannini, M.; D’Auria, E. Nutritional management and follow up of infants and children with food allergy: Italian Society of Pediatric Nutrition/Italian Society of Pediatric Allergy and Immunology Task Force Position Statement. Ital. J. Pediatr. 2014, 40, 1. [Google Scholar] [CrossRef]
- Borrelli, O.; Barbara, G. Neuroimmune interaction and anorectal motility in children with food allergy-related chronic constipation. Am. J. Gastroenterol. 2009, 104, 454–463. [Google Scholar] [CrossRef]
- Lucendo, A.J.; Molina-Infante, J. Guidelines on eosinophilic esophagitis: Evidence-based statements and recommendations for diagnosis and management in children and adults. United Eur. Gastroenterol. J. 2017, 5, 335–358. [Google Scholar] [CrossRef]
- Papadopoulou, A.; Koletzko, S. Management guidelines of eosinophilic esophagitis in childhood. J. Pediatr. Gastroenterol. Nutr. 2014, 58, 107–118. [Google Scholar] [CrossRef]
- Miehlke, S. Clinical features of eosinophilic esophagitis. Dig. Dis. 2014, 32, 61–67. [Google Scholar] [CrossRef]
- Papadopoulou, A.; Dias, J.A. Eosinophilic esophagitis: An emerging disease in childhood—Review of diagnostic and management strategies. Front. Pediatr. 2014, 2, 129. [Google Scholar] [CrossRef]
- Dellon, E.S.; Gonsalves, N. ACG clinical guideline: Evidenced based approach to the diagnosis and management of esophageal eosinophilia and eosinophilic esophagitis (EoE). Am. J. Gastroenterol. 2013, 108, 679–692. [Google Scholar] [CrossRef] [PubMed]
- Hoofien, A.; Dias, J.A. Pediatric Eosinophilic Esophagitis: Results of the European Retrospective Pediatric Eosinophilic Esophagitis Registry (RetroPEER). J. Pediatr. Gastroenterol. Nutr. 2019, 68, 552–558. [Google Scholar] [CrossRef] [PubMed]
- Arias, A.; Gonzalez-Cervera, J. Efficacy of dietary interventions for inducing histologic remission in patients with eosinophilic esophagitis: A systematic review and meta-analysis. Gastroenterology 2014, 146, 1639–1648. [Google Scholar] [CrossRef] [PubMed]
- Loizou, D.; Enav, B. A pilot study of omalizumab in eosinophilic esophagitis. PLoS ONE 2015, 10, e0113483. [Google Scholar] [CrossRef] [PubMed]
- Aceves, S.S.; Chen, D. Mast cells infiltrate the esophageal smooth muscle in patients with eosinophilic esophagitis, express TGF-beta1, and increase esophageal smooth muscle contraction. J. Allergy Clin. Immunol. 2010, 126, 1198–1204.e4. [Google Scholar] [CrossRef] [PubMed]
- Leung, J.; Beukema, K.R. Allergic mechanisms of Eosinophilic oesophagitis. Best Pract. Res. Clin. Gastroenterol. 2015, 29, 709–720. [Google Scholar] [CrossRef] [PubMed]
- Kagalwalla, A.F.; Sentongo, T.A. Effect of six-food elimination diet on clinical and histologic outcomes in eosinophilic esophagitis. Clin. Gastroenterol. Hepatol. 2006, 4, 1097–1102. [Google Scholar] [CrossRef] [PubMed]
- Liacouras, C.A.; Spergel, J.M. Eosinophilic esophagitis: A 10-year experience in 381 children. Clin. Gastroenterol. Hepatol. 2005, 3, 1198–1206. [Google Scholar] [CrossRef]
- Spergel, J.M.; Andrews, T. Treatment of eosinophilic esophagitis with specific food elimination diet directed by a combination of skin prick and patch tests. Ann. Allergy Asthma Immunol. 2005, 95, 336–343. [Google Scholar] [CrossRef]
- Gomez Torrijos, E.; Gonzalez-Mendiola, R. Eosinophilic Esophagitis: Review and Update. Front. Med. (Lausanne) 2018, 5, 247. [Google Scholar] [CrossRef]
- Gonsalves, N.; Yang, G.Y. Elimination diet effectively treats eosinophilic esophagitis in adults; food reintroduction identifies causative factors. Gastroenterology 2012, 142, 1451–1459.e1. [Google Scholar] [CrossRef] [PubMed]
- Molina-Infante, J.; Arias, A. Step-up empiric elimination diet for pediatric and adult eosinophilic esophagitis: The 2-4-6 study. J. Allergy Clin. Immunol. 2018, 141, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Kagalwalla, A.F.; Wechsler, J.B. Efficacy of a 4-Food Elimination Diet for Children with Eosinophilic Esophagitis. Clin. Gastroenterol. Hepatol. 2017, 15, 1698–1707.e7. [Google Scholar] [CrossRef] [PubMed]
- Kagalwalla, A.F.; Amsden, K. Cow’s milk elimination: A novel dietary approach to treat eosinophilic esophagitis. J. Pediatr. Gastroenterol. Nutr. 2012, 55, 711–716. [Google Scholar] [CrossRef] [PubMed]
- Kruszewski, P.G.; Russo, J.M. Prospective, comparative effectiveness trial of cow’s milk elimination and swallowed fluticasone for pediatric eosinophilic esophagitis. Dis. Esophagus 2016, 29, 377–384. [Google Scholar] [CrossRef] [PubMed]
- Erwin, E.A.; Tripathi, A. IgE Antibody Detection and Component Analysis in Patients with Eosinophilic Esophagitis. J. Allergy Clin. Immunol. Pract. 2015, 3, 896–904.e3. [Google Scholar] [CrossRef]
- Clayton, F.; Fang, J.C. Eosinophilic esophagitis in adults is associated with IgG4 and not mediated by IgE. Gastroenterology 2014, 147, 602–609. [Google Scholar] [CrossRef] [PubMed]
- Rosenberg, C.E.; Mingler, M.K. Esophageal IgG4 levels correlate with histopathologic and transcriptomic features in eosinophilic esophagitis. Allergy 2018, 73, 1892–1901. [Google Scholar] [CrossRef]
- Schuyler, A.J.; Wilson, J.M. Specific IgG4 antibodies to cow’s milk proteins in pediatric patients with eosinophilic esophagitis. J. Allergy Clin. Immunol. 2018, 142, 139–148.e12. [Google Scholar] [CrossRef]
- Jyonouchi, S.; Abraham, V. Invariant natural killer T cells from children with versus without food allergy exhibit differential responsiveness to milk-derived sphingomyelin. J. Allergy Clin. Immunol. 2011, 128, 102–109.e13. [Google Scholar] [CrossRef]
- Jyonouchi, S.; Smith, C.L. Invariant natural killer T cells in children with eosinophilic esophagitis. Clin. Exp. Allergy 2014, 44, 58–68. [Google Scholar] [CrossRef] [PubMed]
- Tanabe, S. Analysis of food allergen structures and development of foods for allergic patients. Biosci. Biotechnol. Biochem. 2008, 72, 649–659. [Google Scholar] [CrossRef] [PubMed]
- Kneepkens, C.M.; Meijer, Y. Clinical practice. Diagnosis and treatment of cow’s milk allergy. Eur. J. Pediatr. 2009, 168, 891–896. [Google Scholar] [CrossRef] [PubMed]
- Sicherer, S.H.; Sampson, H.A. Food allergy: A review and update on epidemiology, pathogenesis, diagnosis, prevention, and management. J. Allergy Clin. Immunol. 2018, 141, 41–58. [Google Scholar] [CrossRef] [PubMed]
- Visser, J.T.; Lammers, K. Restoration of impaired intestinal barrier function by the hydrolysed casein diet contributes to the prevention of type 1 diabetes in the diabetes-prone BioBreeding rat. Diabetologia 2010, 53, 2621–2628. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, H.; Pihlanto, A. Food-derived bioactive peptides--opportunities for designing future foods. Curr. Pharm. Des. 2003, 9, 1297–1308. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.B.G.; Gros, M. Immunomodulating properties of protein hydrolysates for application in cow’s milk allergy. Pediatr. Allergy Immunol. 2015, 26, 206–217. [Google Scholar] [CrossRef]
- Wichers, H. Immunomodulation by food: Promising concept for mitigating allergic disease? Anal. Bioanal. Chem. 2009, 395, 37–45. [Google Scholar] [CrossRef]
- Sakaguchi, S.; Powrie, F. Emerging challenges in regulatory T cell function and biology. Science 2007, 317, 627–629. [Google Scholar] [CrossRef]
- Meulenbroek, L.A.; van Esch, B.C. Oral treatment with beta-lactoglobulin peptides prevents clinical symptoms in a mouse model for cow’s milk allergy. Pediatr. Allergy Immunol. 2013, 24, 656–664. [Google Scholar] [CrossRef]
- Ortega-Gonzalez, M.; Capitan-Canadas, F. Validation of bovine glycomacropeptide as an intestinal anti-inflammatory nutraceutical in the lymphocyte-transfer model of colitis. Br. J. Nutr. 2014, 111, 1202–1212. [Google Scholar] [CrossRef] [PubMed]
- van Esch, B.C.; Schouten, B. Oral tolerance induction by partially hydrolyzed whey protein in mice is associated with enhanced numbers of Foxp3+ regulatory T-cells in the mesenteric lymph nodes. Pediatr. Allergy Immunol. 2011, 22, 820–826. [Google Scholar] [CrossRef] [PubMed]
- Espeche Turbay, M.B.; de Moreno de LeBlanc, A. Beta-Casein hydrolysate generated by the cell envelope-associated proteinase of Lactobacillus delbrueckii ssp. lactis CRL 581 protects against trinitrobenzene sulfonic acid-induced colitis in mice. J. Dairy Sci. 2012, 95, 1108–1118. [Google Scholar] [CrossRef] [PubMed]
- Requena, P.; Daddaoua, A. Bovine glycomacropeptide induces cytokine production in human monocytes through the stimulation of the MAPK and the NF-kappaB signal transduction pathways. Br. J. Pharmacol. 2009, 157, 1232–1240. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Wegrzyn, A.; Chehade, M. International consensus guidelines for the diagnosis and management of food protein-induced enterocolitis syndrome: Executive summary-Workgroup Report of the Adverse Reactions to Foods Committee, American Academy of Allergy, Asthma & Immunology. J. Allergy Clin. Immunol. 2017, 139, 1111–1126.e4. [Google Scholar]
- Kiewiet, M.B.G.; van Esch, B. Partially hydrolyzed whey proteins prevent clinical symptoms in a cow’s milk allergy mouse model and enhance regulatory T and B cell frequencies. Mol. Nutr. Food Res. 2017, 61, 1700340. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.R.; Kim, H.S. Mesenteric IL-10-producing CD5+ regulatory B cells suppress cow’s milk casein-induced allergic responses in mice. Sci. Rep. 2016, 6, 19685. [Google Scholar] [CrossRef] [PubMed]
- Kiewiet, M.B.G.; Dekkers, R. Toll-like receptor mediated activation is possibly involved in immunoregulating properties of cow’s milk hydrolysates. PLoS ONE 2017, 12, e0178191. [Google Scholar] [CrossRef]
- Berni Canani, R.; Nocerino, R. Effect of Lactobacillus GG on tolerance acquisition in infants with cow’s milk allergy: A randomized trial. J. Allergy Clin. Immunol. 2012, 129, 580–582. [Google Scholar] [CrossRef]
- Berni Canani, R.; Nocerino, R. Formula selection for management of children with cow’s milk allergy influences the rate of acquisition of tolerance: A prospective multicenter study. J. Pediatr. 2013, 163, 771–777.e1. [Google Scholar] [CrossRef]
- Kiewiet, M.B.G.; Dekkers, R. Immunomodulating protein aggregates in soy and whey hydrolysates and their resistance to digestion in an in vitro infant gastrointestinal model: New insights in the mechanism of immunomodulatory hydrolysates. Food Funct. 2018, 9, 604–613. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Nowak-Wegrzyn, A. Dietary baked milk accelerates the resolution of cow’s milk allergy in children. J. Allergy Clin. Immunol. 2011, 128, 125–131 e2. [Google Scholar] [CrossRef]
- Leonard, S.A.; Nowak-Wegrzyn, A.H. Baked Milk and Egg Diets for Milk and Egg Allergy Management. Immunol. Allergy Clin. North Am. 2016, 36, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Nowak-Wegrzyn, A.; Bloom, K.A. Tolerance to extensively heated milk in children with cow’s milk allergy. J. Allergy Clin. Immunol. 2008, 122, 342–347. [Google Scholar] [CrossRef] [PubMed]
- Shreffler, W.G.; Wanich, N. Association of allergen-specific regulatory T cells with the onset of clinical tolerance to milk protein. J. Allergy Clin. Immunol. 2009, 123, 43–52.e7. [Google Scholar] [CrossRef]
- Caubet, J.C.; Nowak-Wegrzyn, A. Utility of casein-specific IgE levels in predicting reactivity to baked milk. J. Allergy Clin. Immunol. 2013, 131, 222–224. [Google Scholar] [CrossRef] [PubMed]
- Dang, T.D.; Peters, R.L. Debates in allergy medicine: Baked egg and milk do not accelerate tolerance to egg and milk. World Allergy Organ. J. 2016, 9, 2. [Google Scholar] [CrossRef]
- Lambert, R.; Grimshaw, K.E.C. Evidence that eating baked egg or milk influences egg or milk allergy resolution: A systematic review. Clin. Exp. Allergy 2017, 47, 829–837. [Google Scholar] [CrossRef]
- Tordesillas, L.; Berin, M.C. Immunology of Food Allergy. Immunity 2017, 47, 32–50. [Google Scholar] [CrossRef]
- Upton, J.; Nowak-Wegrzyn, A. The Impact of Baked Egg and Baked Milk Diets on IgE- and Non-IgE-Mediated Allergy. Clin. Rev. Allergy Immunol. 2018, 55, 118–138. [Google Scholar] [CrossRef]
- Goldberg, M.R.; Nachshon, L. Efficacy of baked milk oral immunotherapy in baked milk-reactive allergic patients. J. Allergy Clin. Immunol. 2015, 136, 1601–1606. [Google Scholar] [CrossRef] [PubMed]
- D’Auria, E.; Abrahams, M. Personalized Nutrition Approach in Food Allergy: Is It Prime Time Yet? Nutrients 2019, 11, 359. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Mehr, S. Adherence to extensively heated egg and cow’s milk after successful oral food challenge. J. Allergy Clin. Immunol. Pract. 2015, 3, 125–127.e4. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Rautava, S. Human gut colonisation may be initiated in utero by distinct microbial communities in the placenta and amniotic fluid. Sci. Rep. 2016, 6, 23129. [Google Scholar] [CrossRef] [PubMed]
- Pickard, J.M.; Zeng, M.Y. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunol. Rev. 2017, 279, 70–89. [Google Scholar] [CrossRef]
- Van Zwol, A.; Van Den Berg, A. Intestinal microbiota in allergic and nonallergic 1-year-old very low birth weight infants after neonatal glutamine supplementation. Acta Paediatr. 2010, 99, 1868–1874. [Google Scholar] [CrossRef] [PubMed]
- Vandenplas, Y. Prevention and Management of Cow’s Milk Allergy in Non-Exclusively Breastfed Infants. Nutrients 2017, 9, 731. [Google Scholar] [CrossRef]
- Wopereis, H.; Oozeer, R. The first thousand days—Intestinal microbiology of early life: Establishing a symbiosis. Pediatr. Allergy Immunol. 2014, 25, 428–438. [Google Scholar] [CrossRef]
- Pratico, G.; Capuani, G. Exploring human breast milk composition by NMR-based metabolomics. Nat. Prod. Res. 2014, 28, 95–101. [Google Scholar] [CrossRef]
- Mastromarino, P.; Capobianco, D. Correlation between lactoferrin and beneficial microbiota in breast milk and infant’s feces. Biometals 2014, 27, 1077–1086. [Google Scholar] [CrossRef]
- Buccigrossi, V.; de Marco, G. Lactoferrin induces concentration-dependent functional modulation of intestinal proliferation and differentiation. Pediatr. Res. 2007, 61, 410–414. [Google Scholar] [CrossRef] [PubMed]
- Donnet-Hughes, A.; Perez, P.F. Potential role of the intestinal microbiota of the mother in neonatal immune education. Proc. Nutr. Soc. 2010, 69, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Jost, T.; Lacroix, C. New insights in gut microbiota establishment in healthy breast fed neonates. PLoS ONE 2012, 7, e44595. [Google Scholar] [CrossRef] [PubMed]
- Schwartz, S.; Friedberg, I. A metagenomic study of diet-dependent interaction between gut microbiota and host in infants reveals differences in immune response. Genome Biol. 2012, 13, r32. [Google Scholar] [CrossRef] [PubMed]
- Campeotto, F.; Baldassarre, M. Fecal expression of human beta-defensin-2 following birth. Neonatology 2010, 98, 365–369. [Google Scholar] [CrossRef] [PubMed]
- Furuta, S.; Toyama, S. Disposition of polaprezinc (zinc L-carnosine complex) in rat gastrointestinal tract and effect of cimetidine on its adhesion to gastric tissues. J. Pharm. Pharmacol. 1995, 47, 632–636. [Google Scholar] [CrossRef]
- Berin, M.C.; Shreffler, W.G. Mechanisms Underlying Induction of Tolerance to Foods. Immunol. Allergy Clin. N. Am. 2016, 36, 87–102. [Google Scholar] [CrossRef]
- Weissler, K.A.; Caton, A.J. The role of T-cell receptor recognition of peptide:MHC complexes in the formation and activity of Foxp3(+) regulatory T cells. Immunol. Rev. 2014, 259, 11–22. [Google Scholar] [CrossRef]
- West, C.E.; Jenmalm, M.C. The gut microbiota and its role in the development of allergic disease: A wider perspective. Clin. Exp. Allergy 2015, 45, 43–53. [Google Scholar] [CrossRef]
- West, C.E.; Jenmalm, M.C. Probiotics for treatment and primary prevention of allergic diseases and asthma: Looking back and moving forward. Expert Rev. Clin. Immunol. 2016, 12, 625–639. [Google Scholar] [CrossRef]
- Wopereis, H.; Sim, K. Intestinal microbiota in infants at high risk for allergy: Effects of prebiotics and role in eczema development. J. Allergy Clin. Immunol. 2018, 141, 1334–1342.e5. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, T.R.; Jakobsson, H.E. Low diversity of the gut microbiota in infants with atopic eczema. J. Allergy Clin. Immunol. 2012, 129, 434–440. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Konya, T. Infant gut microbiota and food sensitization: Associations in the first year of life. Clin. Exp. Allergy 2015, 45, 632–643. [Google Scholar] [CrossRef] [PubMed]
- Ismail, I.H.; Oppedisano, F. Reduced gut microbial diversity in early life is associated with later development of eczema but not atopy in high-risk infants. Pediatr. Allergy Immunol. 2012, 23, 674–681. [Google Scholar] [CrossRef] [PubMed]
- Boehm, G.; Lidestri, M. Supplementation of a bovine milk formula with an oligosaccharide mixture increases counts of faecal bifidobacteria in preterm infants. Arch. Dis. Child. Fetal Neonatal Ed. 2002, 86, F178–F181. [Google Scholar] [CrossRef]
- Moro, G.; Arslanoglu, S. A mixture of prebiotic oligosaccharides reduces the incidence of atopic dermatitis during the first six months of age. Arch. Dis. Child. 2006, 91, 814–819. [Google Scholar] [CrossRef] [PubMed]
- Alderete, T.L.; Autran, C. Associations between human milk oligosaccharides and infant body composition in the first 6 mo of life. Am. J. Clin. Nutr. 2015, 102, 1381–1388. [Google Scholar] [CrossRef]
- Bode, L.; Contractor, N. Overcoming the limited availability of human milk oligosaccharides: Challenges and opportunities for research and application. Nutr. Rev. 2016, 74, 635–644. [Google Scholar] [CrossRef]
- Seppo, A.E.; Autran, C.A. Human milk oligosaccharides and development of cow’s milk allergy in infants. J. Allergy Clin. Immunol. 2017, 139, 708–711.e5. [Google Scholar] [CrossRef]
- Doherty, A.M.; Lodge, C.J. Human Milk Oligosaccharides and Associations with Immune-Mediated Disease and Infection in Childhood: A Systematic Review. Front. Pediatr. 2018, 6, 91. [Google Scholar] [CrossRef]
- Osborn, D.A.; Sinn, J.K. Prebiotics in infants for prevention of allergy. Cochrane Database Syst. Rev. 2013, CD006474. [Google Scholar] [CrossRef] [PubMed]
- Cuello-Garcia, C.A.; Fiocchi, A. World Allergy Organization-McMaster University Guidelines for Allergic Disease Prevention (GLAD-P): Prebiotics. World Allergy Organ. J. 2016, 9, 10. [Google Scholar] [CrossRef] [PubMed]
- Gruber, C.; van Stuijvenberg, M. Reduced occurrence of early atopic dermatitis because of immunoactive prebiotics among low-atopy-risk infants. J. Allergy Clin. Immunol. 2010, 126, 791–797. [Google Scholar] [CrossRef]
- Francavilla, R.; Calasso, M. Effect of lactose on gut microbiota and metabolome of infants with cow’s milk allergy. Pediatr. Allergy Immunol. 2012, 23, 420–427. [Google Scholar] [CrossRef] [PubMed]
- Candy, D.C.A.; Van Ampting, M.T.J. A synbiotic-containing amino-acid-based formula improves gut microbiota in non-IgE-mediated allergic infants. Pediatr. Res. 2018, 83, 677–686. [Google Scholar] [CrossRef]
- Fox, A.T.; Wopereis, H. A specific synbiotic-containing amino acid-based formula in dietary management of cow’s milk allergy: A randomized controlled trial. Clin. Transl. Allergy 2019, 9, 5. [Google Scholar] [CrossRef] [PubMed]
- Kerperien, J.; Jeurink, P.V. Non-digestible oligosaccharides modulate intestinal immune activation and suppress cow’s milk allergic symptoms. Pediatr. Allergy Immunol. 2014, 25, 747–754. [Google Scholar] [CrossRef]
- Kleinjans, L.; Veening-Griffioen, D.H. Mice co-administrated with partially hydrolysed whey proteins and prebiotic fibre mixtures show allergen-specific tolerance and a modulated gut microbiota. Benef. Microbes 2019, 10, 165–178. [Google Scholar] [CrossRef]
- Braegger, C.; Chmielewska, A. Supplementation of infant formula with probiotics and/or prebiotics: A systematic review and comment by the ESPGHAN committee on nutrition. J. Pediatr. Gastroenterol. Nutr. 2011, 52, 238–250. [Google Scholar] [CrossRef]
- Baldassarre, M.E.; Palladino, V. Rationale of Probiotic Supplementation during Pregnancy and Neonatal Period. Nutrients 2018, 10, 1693. [Google Scholar] [CrossRef]
- Mastromarino, P.; Capobianco, D. Administration of a multistrain probiotic product (VSL#3) to women in the perinatal period differentially affects breast milk beneficial microbiota in relation to mode of delivery. Pharmacol. Res. 2015, 95–96, 63–70. [Google Scholar]
- Baldassarre, M.E.; Di Mauro, A. Administration of a Multi-Strain Probiotic Product to Women in the Perinatal Period Differentially Affects the Breast Milk Cytokine Profile and May Have Beneficial Effects on Neonatal Gastrointestinal Functional Symptoms. A Randomized Clinical Trial. Nutrients 2016, 8, 677. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S.; Walker, W.A. Academy of Breastfeeding Medicine founder’s lecture 2008: Breastfeeding--an extrauterine link between mother and child. Breastfeed. Med. 2009, 4, 3–10. [Google Scholar] [CrossRef] [PubMed]
- Dotterud, C.K.; Storro, O. Probiotics in pregnant women to prevent allergic disease: A randomized, double-blind trial. Br. J. Dermatol. 2010, 163, 616–623. [Google Scholar] [CrossRef] [PubMed]
- Enomoto, T.; Sowa, M. Effects of bifidobacterial supplementation to pregnant women and infants in the prevention of allergy development in infants and on fecal microbiota. Allergol. Int. 2014, 63, 575–585. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kwon, J.H. Effect of probiotic mix (Bifidobacterium bifidum, Bifidobacterium lactis, Lactobacillus acidophilus) in the primary prevention of eczema: A double-blind, randomized, placebo-controlled trial. Pediatr. Allergy Immunol. 2010, 21, e386–e393. [Google Scholar] [CrossRef]
- Niers, L.; Martin, R. The effects of selected probiotic strains on the development of eczema (the PandA study). Allergy 2009, 64, 1349–1358. [Google Scholar] [CrossRef]
- Ou, C.Y.; Kuo, H.C. Prenatal and postnatal probiotics reduces maternal but not childhood allergic diseases: A randomized, double-blind, placebo-controlled trial. Clin. Exp. Allergy 2012, 42, 1386–1396. [Google Scholar] [CrossRef]
- Rautava, S.; Kainonen, E. Maternal probiotic supplementation during pregnancy and breast-feeding reduces the risk of eczema in the infant. J. Allergy Clin. Immunol. 2012, 130, 1355–1360. [Google Scholar] [CrossRef]
- Simpson, M.R.; Dotterud, C.K. Perinatal probiotic supplementation in the prevention of allergy related disease: 6 year follow up of a randomised controlled trial. BMC Dermatol. 2015, 15, 13. [Google Scholar] [CrossRef]
- Wickens, K.; Barthow, C. Maternal supplementation alone with Lactobacillus rhamnosus HN001 during pregnancy and breastfeeding does not reduce infant eczema. Pediatr. Allergy Immunol. 2018, 29, 296–302. [Google Scholar] [CrossRef] [PubMed]
- Fiocchi, A.; Pawankar, R. World Allergy Organization-McMaster University Guidelines for Allergic Disease Prevention (GLAD-P): Probiotics. World Allergy Organ. J. 2015, 8, 4. [Google Scholar] [CrossRef] [PubMed]
- Muraro, A.; Agache, I. EAACI food allergy and anaphylaxis guidelines: Managing patients with food allergy in the community. Allergy 2014, 69, 1046–1057. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, M.E.; Laforgia, N. Lactobacillus GG improves recovery in infants with blood in the stools and presumptive allergic colitis compared with extensively hydrolyzed formula alone. J. Pediatr. 2010, 156, 397–401. [Google Scholar] [CrossRef] [PubMed]
- Kapel, N.; Campeotto, F. Faecal calprotectin in term and preterm neonates. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 542–547. [Google Scholar] [CrossRef]
- Tan-Lim, C.S.C.; Esteban-Ipac, N.A.R. Probiotics as treatment for food allergies among pediatric patients: A meta-analysis. World Allergy Organ. J. 2018, 11, 25. [Google Scholar] [CrossRef] [PubMed]
- Berni Canani, R.; Di Costanzo, M. Extensively hydrolyzed casein formula containing Lactobacillus rhamnosus GG reduces the occurrence of other allergic manifestations in children with cow’s milk allergy: 3-year randomized controlled trial. J. Allergy Clin. Immunol. 2017, 139, 1906–1913.e4. [Google Scholar] [CrossRef]
- Dupont, C.; Hol, J.; Nieuwenhuis, E.E. An extensively hydrolysed casein-based formula for infants with cows’ milk protein allergy: tolerance/hypo-allergenicity and growth catch-up. Br. J. Nutr. 2015, 113, 1102–1112. [Google Scholar] [CrossRef]
- Hol, J.; Van leer, E.H.; Elink schuurman, B.E.; de Ruiter, L.F.; Samsom, J.N.; Hop, W.; Neijens, H.J.; de Jongste, J.C.; Nieuwenhuis, E.E.; Cow’s Milk Allergy Modified by Elimination and Lactobacilli study group. The acquisition of tolerance toward cow’s milk through probiotic supplementation: a randomized, controlled trial. J. Allergy Clin. Immunol. 2008, 121, 1448–1454. [Google Scholar] [CrossRef]
- Kirjavainen, P.V.; Salminen, S.J.; Isolauri, E. Probiotic bacteria in the management of atopic disease: underscoring the importance of viability. J. Pediatr. Gastroenterol. Nutr. 2003, 36, 223–227. [Google Scholar] [CrossRef]
- Majamaa, H.; Isolauri, E. Probiotics: a novel approach in the management of food allergy. J. Allergy Clin. Immunol. 1997, 99, 179–185. [Google Scholar] [CrossRef]
- Viljanen, M.; Savilahti, E.; Haahtela, T.; Juntunen-Backman, K.; Korpela, R.; Poussa, T.; Tuure, T.; Kuitunen, M. Probiotics in the treatment of atopic eczema/dermatitis syndrome in infants: a double-blind placebo-controlled trial. Allergy 2005, 60, 494–500. [Google Scholar] [CrossRef] [PubMed]
- Kuitunen, M.; Kukkonen, K. Pro- and prebiotic supplementation induces a transient reduction in hemoglobin concentration in infants. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 626–630. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, M.E. Probiotic Genera/Species Identification Is Insufficient for Evidence-Based Medicine. Am. J. Gastroenterol. 2018, 113, 1561. [Google Scholar] [CrossRef] [PubMed]
- Baldassarre, M.E. Harms Reporting in Randomized Controlled Trials of Interventions Aimed at Modifying Microbiota. Ann. Intern. Med. 2019, 170, 143. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Author, Year | Study | Subjects | Strain, Dose, Beginning of the Treatment (S), End of the Treatment (E) | Placebo | Outcomes | Follow-Up (Years) | Side Effects |
|---|---|---|---|---|---|---|---|
| Dotterud et al. [134] | RCT | 415 pregnant women | LGG 5 × 1010 CFU, Bb-12 5 × 1010 CFU and La-5. 5 × 109 (CFU) daily S: 4 weeks before expected delivery date E: 3 weeks after delivery (breastfeeding) | yes | Probiotic supplementation reduces incidence of atopic dermatitis (AD) in children | 2 | No |
| Enomoto et al. [135] | Open-trial | 166 pregnant women and newborns | BB536 5 × 109 CFU and BB M-16V 5 × 109 CFU daily S: 4 weeks before expected delivery date E: 6 months after delivery (to infants) | no | Probiotic supplementation reduces incidence of AD in children | 3 | no |
| Wickens et al. [141] | RCT | 423 pregnant women | LR HN001 6 × 109 CFU daily S: from 14–16 weeks gestation E: 6 months after delivery (breastfeeding) | yes | Probiotic supplementation does not prevent AD in infants | 1 | no |
| Ou et al. [138] | RCT | 191 pregnant women and related newborns | LGG ATCC 53103, 1 × 1010 CFU daily S: From the second trimester of pregnancy; E: 6 months after delivery (to mothers and infants) during breastfeeding | yes | Probiotic supplementation doesn’t prevent infant allergic disease (AD, allergic rhinitis, asthma) | 3 | no |
| Rautava et al. [139] | RCT | 241 pregnant women | LPR 1 × 109 CFU BL999 1 × 109 CFU ST11 1 × 109 CFU daily S: 2 months before expected delivery E: 2 months after delivery (breast-feeding) | yes | Probiotic supplementation prevents infant eczema | 2 | Not observed |
| Kim et al. [136] | Randomized placebo-controlled trial | 112 pregnant women and newborns | BGN4 1.6 × 109 CFU, AD011 1.6 × 109 CFU, and AD031 1.6 × 109 CFU daily S: 4–8 weeks before expected delivery E: 6 months after delivery (to mothers during breastfeeding and to infants) | yes | Probiotics supplementation reduces incidence of AD in children | 1 | yes |
| Niers et al. [137] | Double-blind, randomized, placebo-controlled trial | 136 pregnant women and newborns | BB: 1 × 109 CFU; BL 1 × 109 CFU; LL 1 × 109 CFU S: last 6 weeks of pregnancy E: 12 months after delivery (to infants) | yes | Probiotics supplementation reduces the incidence of AD in children at 3 months of life | 24 months after delivery | no |
| Simpson et al. [140] | Randomized placebo-controlled trial | 415 pregnant women | Probiotic milk: LGG, 5 × 1010 CFU; La-5 5 × 109 CFU and Bb-12 5 × 1010 CFU S: from 36 weeks gestation E: 3 months after delivery (breastfeeding) | yes | Probiotics supplementation reduces incidence of AD | 6 years after delivery | no |
| Author, Year | Study Design | Subjects | Strain, Dose (D) | Placebo | Outcomes | Treatment Period (Months) | Side Effects |
|---|---|---|---|---|---|---|---|
| Baldassarre et al. [144] | RCT | 30 infants | LGG 1 × 106 CFU/g | yes | Probiotic supplementation improves gastrointestinal symptoms (hematochezia and fecal calprotectin) | 1 | No |
| Berni Canani et al. [79] | RCT | 80 infants | LGG, 1.4 × 107 CFU/100 mL | yes | Probiotic supplementation accelerates tolerance acquisition to cow’s milk proteins | 12 | No |
| Berni Canani et al. [80] | RCT | 260 infants | LGG (dose not specified) | yes | Probiotic supplementation accelerates tolerance acquisition to cow’s milk proteins | 12 | No |
| Berni Canani et al. [147] | RCT | 220 children | LGG (dose not specified) | yes | Probiotic supplementation reduces the incidence of other allergic manifestations and hastens the development of oral tolerance to cow’s milk proteins | 36 | No |
| Dupont et al. [148] | RCT | 119 infants | LC CRL431 and Bb-12 (dose not specified) | yes | Probiotic supplementation significantly improves the SCORAD index and growth indices | 6 | No |
| Hol et al. [149] | RCT | 119 infants | LC CRL431 and Bb-12 1 × 107 CFU/g formula | yes | Probiotic supplementation does not accelerate tolerance acquisition to cow’s milk proteins | 6 | No |
| Kirjavainen et al. [150] | RCT | 35 infants | LGG ATCC 53103 1 × 109 CFU/g | yes | Supplementation with viable probiotics improves the SCORAD index | 2 | Diarrhea (with heat-inactivated LGG) |
| Majamaa et al. [151] | RCT | 31 infants | LGG ATCC 53103- 5 × 108 CFU/g formula twice a day | yes | Probiotic supplementation improves the SCORAD index and reduces markers of intestinal inflammation | 1 | No |
| Viljanen et al. [152] | RCT | 230 infants | LGG (ATCC 53103) 5 × 109 CFU vs. LGG 5 × 109 CFU, LR LC705- 5 × 109 CFU, Bbi99- 2 × 108 CFU, and PJS- 2 × 109 CFU twice a day | yes | Probiotic supplementation improves the SCORAD index in IgE-sensitized infants but not in non-IgE-sensitized infants | 1 | No |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
D’Auria, E.; Salvatore, S.; Pozzi, E.; Mantegazza, C.; Sartorio, M.U.A.; Pensabene, L.; Baldassarre, M.E.; Agosti, M.; Vandenplas, Y.; Zuccotti, G. Cow’s Milk Allergy: Immunomodulation by Dietary Intervention. Nutrients 2019, 11, 1399. https://doi.org/10.3390/nu11061399
D’Auria E, Salvatore S, Pozzi E, Mantegazza C, Sartorio MUA, Pensabene L, Baldassarre ME, Agosti M, Vandenplas Y, Zuccotti G. Cow’s Milk Allergy: Immunomodulation by Dietary Intervention. Nutrients. 2019; 11(6):1399. https://doi.org/10.3390/nu11061399
Chicago/Turabian StyleD’Auria, Enza, Silvia Salvatore, Elena Pozzi, Cecilia Mantegazza, Marco Ugo Andrea Sartorio, Licia Pensabene, Maria Elisabetta Baldassarre, Massimo Agosti, Yvan Vandenplas, and GianVincenzo Zuccotti. 2019. "Cow’s Milk Allergy: Immunomodulation by Dietary Intervention" Nutrients 11, no. 6: 1399. https://doi.org/10.3390/nu11061399
APA StyleD’Auria, E., Salvatore, S., Pozzi, E., Mantegazza, C., Sartorio, M. U. A., Pensabene, L., Baldassarre, M. E., Agosti, M., Vandenplas, Y., & Zuccotti, G. (2019). Cow’s Milk Allergy: Immunomodulation by Dietary Intervention. Nutrients, 11(6), 1399. https://doi.org/10.3390/nu11061399