Human Enriched Serum Following Hydrolysed Collagen Absorption Modulates Bone Cell Activity: from Bedside to Bench and Vice Versa


 ,
,  , ,
, , 
Abstract

1. Introduction
2. Materials and Methods
2.1. Ethics
2.2. Hydrolysed Collagens
2.3. Cell Cultures
2.4. Dietary Supplementation and Ovariectomy-Related Bone Loss
2.5. Tissue Sampling, Biochemical Parameters and Bone Mineral Density Analysis
2.6. Metabolism Models and Serum Collection
2.7. In Vitro and Ex Vivo Assays
2.8. Statistical Analysis
3. Results
3.1. Bovine HC Promotes Osteoblast Proliferation, Differentiation and Function in Vitro
3.2. B2000 Significantly Reduces Bone Loss in Vivo by Modulating the Level of RANKL
3.3. B2000-Enriched Mouse Serum Stimulates Osteoblast Function While Repressing Osteoclastogenesis
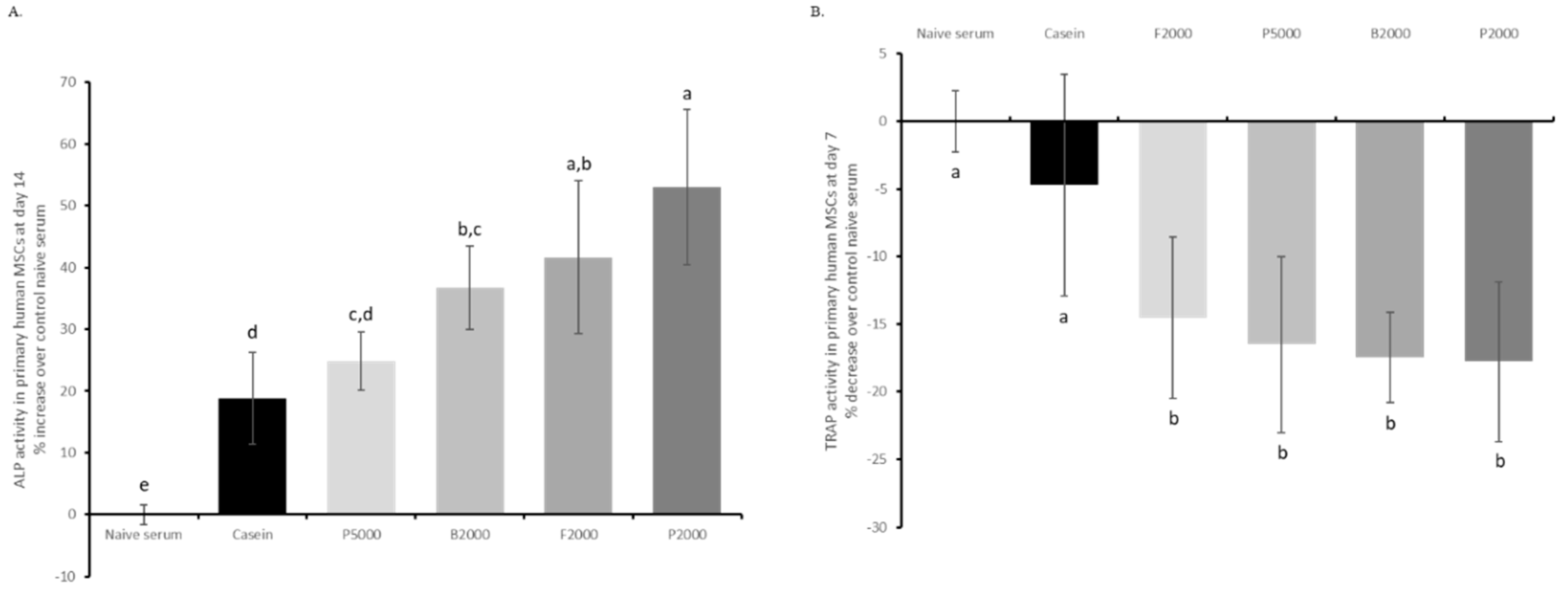
3.4. Assays with Human Enriched Sera Confirm the in Vivo Data and Allow Comparative Activity Screening.
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hanley, D.A.; McClung, M.R.; Davison, K.S.; Dian, L.; Harris, S.T.; Miller, P.D.; Lewiecki, E.M.; Kendler, D.L. Western osteoporosis alliance clinical practice series: Evaluating the balance of benefits and risks of long-term osteoporosis therapies. Am. J. Med. 2017, 130, 862-e1. [Google Scholar] [CrossRef] [PubMed]
- Lotters, F.J.; Lenoir-Wijnkoop, I.; Fardellone, P.; Rizzoli, R.; Rocher, E.; Poley, M.J. Dairy foods and osteoporosis: An example of assessing the health-economic impact of food products. Osteoporos. Int. 2013, 24, 139–150. [Google Scholar] [CrossRef] [PubMed]
- Elam, M.L.; Elam, M.L.; Johnson, S.A.; Hooshmand, S.; Feresin, R.G.; Payton, M.E.; Gu, J.; Arjmandi, B.H. A calcium-collagen chelate dietary supplement attenuates bone loss in postmenopausal women with osteopenia: A randomized controlled trial. J. Med. Food 2015, 18, 324–331. [Google Scholar] [CrossRef] [PubMed]
- Konig, D.; König, D.; Oesser, S.; Scharla, S.; Zdzieblik, D.; Gollhofer, A. Specific collagen peptides improve bone mineral density and bone markers in Postmenopausal women—A randomized controlled study. Nutrients 2018, 10, 97. [Google Scholar] [CrossRef] [PubMed]
- Nieves, J. Skeletal effects of nutrients and nutraceuticals, beyond calcium and vitamin D. Osteoporos. Int. 2013, 24, 771–786. [Google Scholar] [CrossRef] [PubMed]
- Ricard-Blum, S. The collagen family. Cold Spring Harb. Perspect. Biol. 2011, 3, a004978. [Google Scholar] [CrossRef] [PubMed]
- Viguet-Carrin, S.; Garnero, P.; Delmas, P.D. The role of collagen in bone strength. Osteoporos Int. 2006, 17, 319–336. [Google Scholar] [CrossRef] [PubMed]
- Currey, J.D. Role of collagen and other organics in the mechanical properties of bone. Osteoporos Int. 2003, 14, 29–36. [Google Scholar]
- Moskowitz, R.W. Role of collagen hydrolysate in bone and joint disease. Semin. Arthritis Rheum. 2000, 30, 87–99. [Google Scholar] [CrossRef]
- Trc, T.; Bohmova, J. Efficacy and tolerance of enzymatic hydrolysed collagen (EHC) vs. glucosamine sulphate (GS) in the treatment of knee osteoarthritis (KOA). Int. Orthop. 2011, 35, 341–348. [Google Scholar] [CrossRef]
- Fujita, T.; Ohue, M.; Fujii, Y.; Miyauchi, A.; Takagi, Y. The effect of active absorbable algal calcium (AAA Ca) with collagen and other matrix components on back and joint pain and skin impedance. J. Bone Miner. Metab. 2002, 20, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Bruyere, O.; Zegels, B.; Leonori, L.; Rabenda, V.; Janssen, A.; Bourges, C.; Reginster, J.Y. Effect of collagen hydrolysate in articular pain: A 6-month randomized, double-blind, placebo controlled study. Complement. Ther. Med. 2012, 20, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Misner, B.; Bagchi, M.; Kothari, S.C.; Downs, B.W.; Fafard, R.D.; Preuss, H.G. Effects of orally administered undenatured type II collagen against arthritic inflammatory diseases: a mechanistic exploration. Int. J. Clin. Pharmacol. Res. 2002, 22, 101–110. [Google Scholar] [PubMed]
- Henrotin, Y.; Henrotin, Y.; Lambert, C.; Couchourel, D.; Ripoll, C.; Chiotelli, E. Nutraceuticals: do they represent a new era in the management of osteoarthritis?—A narrative review from the lessons taken with five products. Osteoarthr. Cartil. 2011, 19, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Hooshmand, S.; Elam, M.L.; Browne, J.; Campbell, S.C.; Payton, M.E. Evidence for bone reversal properties of a calcium-collagen chelate, a novel dietary supplement. J. Food Nutr. Disord. 2013, 2, 1. [Google Scholar]
- Comblain, F.; Barthélémy, N.; Lefèbvre, M.; Schwartz, C.; Lesponne, I.; Serisier, S.; Feugier, A.; Balligand, M.; Henrotin, Y. A randomized, double-blind, prospective, placebo-controlled study of the efficacy of a diet supplemented with curcuminoids extract, hydrolyzed collagen and green tea extract in owner’s dogs with osteoarthritis. BMC Vet. Res. 2017, 13, 395. [Google Scholar] [CrossRef] [PubMed]
- Bruyere, O.; Rizzoli, R.; Coxam, V.; Avouac, B.; Chevalier, T.; Fabien-Soulé, V.; Kanis, J.A.; Kaufman, J.M.; Tsouderos, Y.; Reginster, J.Y. Assessment of health claims in the field of bone: a view of the group for the respect of ethics and excellence in science (GREES). Osteoporos. Int. 2012, 23, 193–199. [Google Scholar] [CrossRef]
- Wittrant, Y.; Gorin, Y.; Woodruff, K.; Horn, D.; Abboud, H.E.; Mohan, S.; Abboud-Werner, S.L. High d (+) glucose concentration inhibits RANKL-induced osteoclastogenesis. Bone 2008, 42, 1122–1130. [Google Scholar] [CrossRef]
- Adam, M.; Spacek, P.; Hulejova, H.; Galianova, A.; Blahos, J. Postmenopausal osteoporosis. Treatment with calcitonin and a diet rich in collagen proteins. Cas. Lek. Cesk. 1996, 135, 74–78. [Google Scholar]
- Cuneo, F.; Costa-Paiva, L.; Pinto-Neto, A.M.; Morais, S.S.; Amaya-Farfan, J. Effect of dietary supplementation with collagen hydrolysates on bone metabolism of postmenopausal women with low mineral density. Maturitas 2010, 65, 253–257. [Google Scholar] [CrossRef]
- Guillerminet, F.; Beaupied, H.; Fabien-Soulé, V.; Tomé, D.; Benhamou, C.L.; Roux, C.; Blais, A. Hydrolyzed collagen improves bone metabolism and biomechanical parameters in ovariectomized mice: an in vitro and in vivo study. Bone 2010, 46, 827–834. [Google Scholar] [CrossRef] [PubMed]
- Guillerminet, F.; Fabien-Soulé, V.; Even, P.C.; Tomé, D.; Benhamou, C.L.; Roux, C.; Blais, A. Hydrolyzed collagen improves bone status and prevents bone loss in ovariectomized C3H/HeN mice. Osteoporos. Int. 2012, 23, 1909–1919. [Google Scholar] [CrossRef] [PubMed]
- Okiura, T.; Oishi, Y.; Takemura, A.; Ishihara, A. Effects of collagen hydrolysate on the tibialis anterior muscle and femur in senescence-accelerated mouse prone 6. J. Musculoskelet. Neuronal Interact. 2016, 16, 161–167. [Google Scholar] [PubMed]
- Zhang, L.; Zhang, S.; Song, H.; Li, B. Effect of collagen hydrolysates from silver carp skin (hypophthalmichthys molitrix) on osteoporosis in chronologically aged mice: Increasing bone remodeling. Nutrients 2018, 10, 1434. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Machado, G.C.; Eyles, J.P.; Ravi, V.; Hunter, D.J. Dietary supplements for treating osteoarthritis: a systematic review and meta-analysis. Br. J. Sports Med. 2018, 52, 167–175. [Google Scholar] [CrossRef] [PubMed]
- Food and Drug Administration; Center for Drug Evaluation and Research. Estimating the maximum safe starting dose in initial clinical trials for therapeutics in adult healthy volunteers. Pharmacol. Toxicol. 2005. [Google Scholar]
- Ichikawa, S.; Morifuji, M.; Ohara, H.; Matsumoto, H.; Takeuchi, Y.; Sato, K. Hydroxyproline-containing dipeptides and tripeptides quantified at high concentration in human blood after oral administration of gelatin hydrolysate. Int. J. Food Sci. Nutr. 2010, 61, 52–60. [Google Scholar] [CrossRef]
- Shigemura, Y.; Kubomura, D.; Sato, Y.; Sato, K. Dose-dependent changes in the levels of free and peptide forms of hydroxyproline in human plasma after collagen hydrolysate ingestion. Food Chem. 2014, 159, 328–332. [Google Scholar] [CrossRef]
- Taga, Y.; Kusubata, M.; Ogawa-Goto, K.; Hattori, S. Highly accurate quantification of hydroxyproline-containing peptides in blood using a protease digest of stable isotope-labeled collagen. J. Agric. Food Chem. 2014, 62, 12096–12102. [Google Scholar] [CrossRef]
- Taga, Y.; Kusubata, M.; Ogawa-Goto, K.; Hattori, S. Identification of collagen-derived hydroxyproline (hyp)-containing cyclic dipeptides with high oral bioavailability: efficient formation of cyclo(x-hyp) from x-hyp-gly-type tripeptides by heating. J. Agric. Food Chem. 2017, 65, 9514–9521. [Google Scholar] [CrossRef]
- Kim, H.K.; Kim, M.G.; Leem, K.H. Osteogenic activity of collagen peptide via ERK/MAPK pathway mediated boosting of collagen synthesis and its therapeutic efficacy in osteoporotic bone by back-scattered electron imaging and microarchitecture analysis. Molecules 2013, 18, 15474–15489. [Google Scholar] [CrossRef] [PubMed]
- K Kim, H.K.; Kim, M.G.; Leem, K.H. Collagen hydrolysates increased osteogenic gene expressions via a MAPK signaling pathway in MG-63 human osteoblasts. Food Funct. 2014, 5, 573–578. [Google Scholar] [CrossRef]
- Liu, J.; Zhang, B.; Song, S.; Ma, M.; Si, S.; Wang, Y.; Xu, B.; Feng, K.; Wu, J.; Guo, Y. Bovine collagen peptides compounds promote the proliferation and differentiation of MC3T3-E1 pre-osteoblasts. PLoS ONE 2014, 9, e99920. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Nagaoka, H.; Terajima, M.; Tsuda, N.; Hayashi, Y.; Yamauchi, M. Effects of fish collagen peptides on collagen post-translational modifications and mineralization in an osteoblastic cell culture system. Dent. Mater. J. 2013, 32, 88–95. [Google Scholar] [CrossRef] [PubMed]
- Tsuruoka, N.; Yamato, R.; Sakai, Y.; Yoshitake, Y.; Yonekura, H. Promotion by collagen tripeptide of type I collagen gene expression in human osteoblastic cells and fracture healing of rat femur. Biosci. Biotechnol. Biochem. 2007, 71, 2680–2687. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Yoshizawa, Y.; Kawakubo, A.; Ikeda, T.; Yanagiguchi, K.; Hayashi, Y. Early gene and protein expression associated with osteoblast differentiation in response to fish collagen peptides powder. Dent. Mater. J. 2013, 32, 233–240. [Google Scholar] [CrossRef] [PubMed]
- Liu, C.; Sun, J. Hydrolyzed tilapia fish collagen induces osteogenic differentiation of human periodontal ligament cells. Biomed. Mater. 2015, 10, 065020. [Google Scholar] [CrossRef] [PubMed]
- Kawaguchi, T.; Nanbu, P.N.; Kurokawa, M. Distribution of prolylhydroxyproline and its metabolites after oral administration in rats. Biol. Pharm. Bull. 2012, 35, 422–427. [Google Scholar] [CrossRef]
- Dimitri, P.; Rosen, C. Fat and bone: Where are we now? Calcif. Tissue Int. 2017, 100, 431–432. [Google Scholar] [CrossRef]
- Tagliaferri, C.; Salles, J.; Landrier, J.F.; Giraudet, C.; Patrac, V.; Lebecque, P.; Davicco, M.J.; Chanet, A.; Pouyet, C.; Dhaussy, A.; et al. Increased body fat mass and tissue lipotoxicity associated with ovariectomy or high-fat diet differentially affects bone and skeletal muscle metabolism in rats. Eur. J. Nutr. 2015, 54, 1139–1149. [Google Scholar] [CrossRef]
- Ding, K.H.; Cain, M.; Davis, M.; Bergson, C.; McGee-Lawrence, M.; Perkins, C.; Hardigan, T.; Shi, X.; Zhong, Q.; Xu, J.; et al. Amino acids as signaling molecules modulating bone turnover. Bone 2018, 115, 15–24. [Google Scholar] [CrossRef] [PubMed]
- Mine, Y.; Mine, Y.; Makihira, S.; Yamaguchi, Y.; Tanaka, H.; Nikawa, H. Involvement of ERK and p38 MAPK pathways on Interleukin-33-induced RANKL expression in osteoblastic cells. Cell Biol. Int. 2014, 38, 655–662. [Google Scholar] [CrossRef]
- Mizuno, M.; Kuboki, Y. Osteoblast-related gene expression of bone marrow cells during the osteoblastic differentiation induced by type I collagen. J. Biochem. 2001, 129, 133–138. [Google Scholar] [CrossRef] [PubMed]
- Daneault, A.; Prawitt, J.; Fabien Soulé, V.; Coxam, V.; Wittrant, Y. Biological effect of hydrolyzed collagen on bone metabolism. Crit. Rev. Food Sci. Nutr. 2017, 57, 1922–1937. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Sugihara, F.; Suzuki, K.; Inoue, N.; Venkateswarathirukumara, S. A double-blind, placebo-controlled, randomised, clinical study on the effectiveness of collagen peptide on osteoarthritis. J. Sci. Food Agric. 2015, 95, 702–707. [Google Scholar] [CrossRef] [PubMed]
- Watanabe-Kamiyama, M.; Shimizu, M.; Kamiyama, S.; Taguchi, Y.; Sone, H.; Morimatsu, F.; Shirakawa, H.; Furukawa, Y.; Komai, M. Absorption and effectiveness of orally administered low molecular weight collagen hydrolysate in rats. J. Agric. Food Chem. 2010, 58, 835–841. [Google Scholar] [CrossRef] [PubMed]
- Leem, K.H.; Lee, S.; Jang, A.; Kim, H.K. Porcine skin gelatin hydrolysate promotes longitudinal bone growth in adolescent rats. J. Med. Food 2013, 16, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Walsh, M.C.; Choi, Y. Biology of the RANKL-RANK-OPG system in immunity, bone, and beyond. Front. Immunol. 2014, 5, 511. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CTRL (RQ) | CASEIN (RQ) | CASEIN (p-Value) | B2000 (RQ) | B2000 (p-Value) | |
|---|---|---|---|---|---|
| Acp5 | 1.0 | 0.53 | 0.54 | 0.45 | 0.03 |
| Car2 | 1.0 | 0.52 | 0.32 | 0.52 | 0.02 |
| Csf1r | 1.0 | 0.57 | 0.03 | 0.78 | 0.02 |
| Ctsk | 1.0 | 0.69 | 0.72 | 0.75 | 0.06 |
| Itgb3 | 1.0 | 0.47 | 0.39 | 0.53 | 0.06 |
| Mmp9 | 1.0 | 0.55 | 0.56 | 0.63 | 0.08 |
| Nos2 | 1.0 | 0.47 | 0.15 | 0.24 | 0.06 |
| Tgfbr1 | 1.0 | 0.43 | 0.37 | 0.34 | 0.06 |
| Tnfrsf1b | 1.0 | 0.58 | 0.17 | 0.49 | 0.07 |
| Traf2 | 1.0 | 0.56 | 0.12 | 0.69 | 0.06 |
| Traf6 | 1.0 | 0.60 | 0.05 | 0.64 | 0.04 |
| CASEIN (RQ) | B2000 (RQ) | B2000 (p-Value) | |
|---|---|---|---|
| Cd36 | 1.0 | 0.66 | 0.04 |
| Itgam | 1.0 | 0.86 | 0.06 |
| Tlr2 | 1.0 | 0.81 | 0.06 |
| Tnfrsf1b | 1.0 | 0.77 | 0.03 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wauquier, F.; Daneault, A.; Granel, H.; Prawitt, J.; Fabien Soulé, V.; Berger, J.; Pereira, B.; Guicheux, J.; Rochefort, G.Y.; Meunier, N.; et al. Human Enriched Serum Following Hydrolysed Collagen Absorption Modulates Bone Cell Activity: from Bedside to Bench and Vice Versa. Nutrients 2019, 11, 1249. https://doi.org/10.3390/nu11061249
Wauquier F, Daneault A, Granel H, Prawitt J, Fabien Soulé V, Berger J, Pereira B, Guicheux J, Rochefort GY, Meunier N, et al. Human Enriched Serum Following Hydrolysed Collagen Absorption Modulates Bone Cell Activity: from Bedside to Bench and Vice Versa. Nutrients. 2019; 11(6):1249. https://doi.org/10.3390/nu11061249
Chicago/Turabian StyleWauquier, Fabien, Audrey Daneault, Henri Granel, Janne Prawitt, Véronique Fabien Soulé, Juliette Berger, Bruno Pereira, Jérôme Guicheux, Gael Y. Rochefort, Nathalie Meunier, and et al. 2019. "Human Enriched Serum Following Hydrolysed Collagen Absorption Modulates Bone Cell Activity: from Bedside to Bench and Vice Versa" Nutrients 11, no. 6: 1249. https://doi.org/10.3390/nu11061249
APA StyleWauquier, F., Daneault, A., Granel, H., Prawitt, J., Fabien Soulé, V., Berger, J., Pereira, B., Guicheux, J., Rochefort, G. Y., Meunier, N., Blot, A., & Wittrant, Y. (2019). Human Enriched Serum Following Hydrolysed Collagen Absorption Modulates Bone Cell Activity: from Bedside to Bench and Vice Versa. Nutrients, 11(6), 1249. https://doi.org/10.3390/nu11061249