3-(4-Hydroxy-3-methoxyphenyl)propionic Acid Produced from 4-Hydroxy-3-methoxycinnamic Acid by Gut Microbiota Improves Host Metabolic Condition in Diet-Induced Obese Mice

Abstract
1. Introduction
2. Materials and Methods
2.1. Mice, Diet, and Experimental Design
2.2. Plasma Biochemical Analyses
2.3. Quantification of Hepatic Triglyceride Content
2.4. Hepatic Histology
2.5. Quantification of Phytochemicals by HPLC
2.6. Analysis of Gut Microbiota by 16S rRNA Gene Sequencing
2.7. Real Time-PCR (RT-PCR)
2.8. Statistical Analysis
3. Results
3.1. HMCA Intake Suppresses HFD-Induced Obesity
3.2. Gut Microbiota Convert HMCA into HMPA in the Intestine
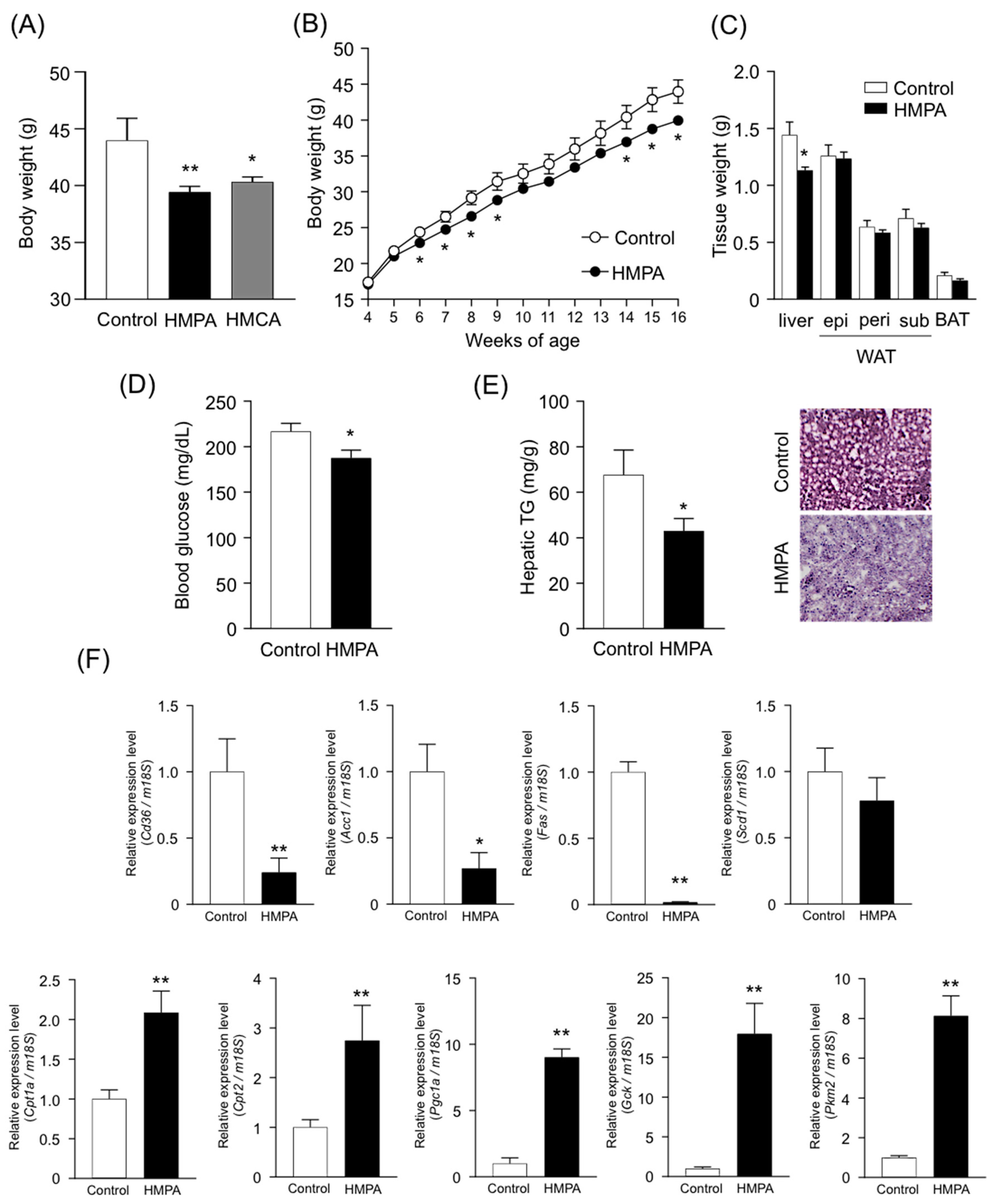
3.3. HMPA is Molecular Entity Underlying Metabolic Improvement Following HMCA Intake
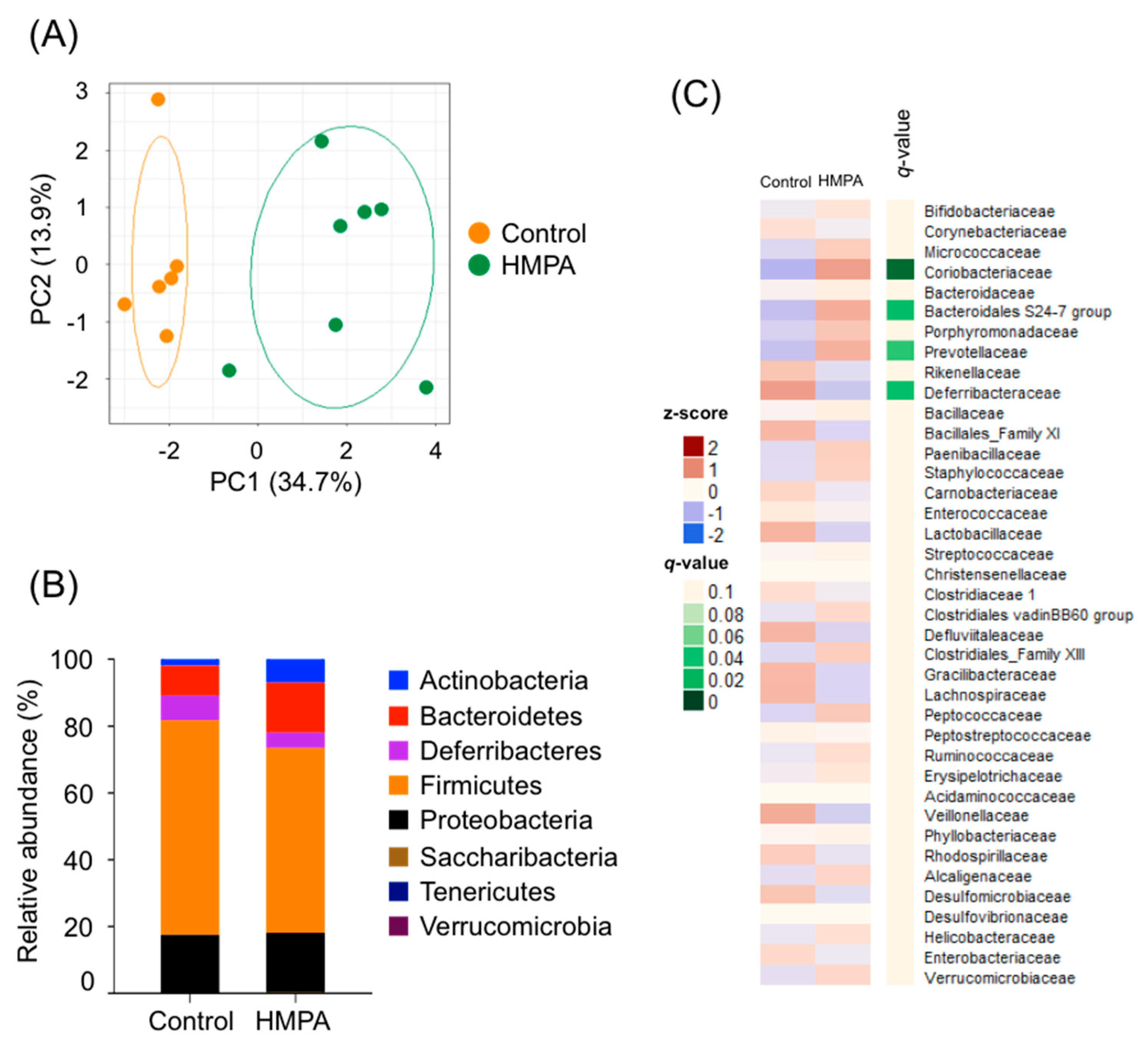
3.4. HMPA Intake Modifies the Changes of HFD-Associated Gut Microbial Composition
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Redeuil, K.; Smarrito-Menozzi, C.; Guy, P.; Rezzi, S.; Dionisi, F.; Williamson, G.; Nagy, K.; Renouf, M. Identification of novel circulating coffee metabolites in human plasma by liquid chromatography–mass spectrometry. J. Chromatogr. A 2011, 1218, 4678–4688. [Google Scholar] [CrossRef] [PubMed]
- Nieber, K. The impact of coffee on health. Planta Med. 2017, 83, 1256–1263. [Google Scholar] [CrossRef] [PubMed]
- Smith, M.M.; Hartley, R.D. Occurrence and nature of ferulic acid substitution of cell wall polysaccharides in gramineous plants. Carbohydr. Res. 1983, 118, 65–80. [Google Scholar] [CrossRef]
- Zubieta, C.; Kota, P.; Ferrer, J.L.; Dixon, R.A.; Noel, J.P. Structural basis for the modulation of lignin monomer methylation by caffeic acid/5-hydroxyferulic acid 3/5-Omethyltransferase. Plant Cell 2002, 14, 1265–1277. [Google Scholar] [CrossRef] [PubMed]
- Gómez-Ruiz, J.A.; Leake, D.S.; Ames, J.M. In vitro antioxidant activity of coffee compounds and their metabolites. J. Agric. Food Chem. 2007, 55, 6962–6969. [Google Scholar] [CrossRef] [PubMed]
- Chowdhury, S.; Ghosh, S.; Das, A.K.; Sil, P.C. Ferulic acid protects hyperglycemia-induced kidney damage by regulating oxidative insult, inflammation and autophagy. Front. Pharmacol. 2019, 10, 27. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.E.; Park, E. Ferulic acid in combination with PARP inhibitor sensitizes breast cancer cells as chemotherapeutic strategy. Biochem. Biophys. Res. Commun. 2015, 458, 520–524. [Google Scholar] [CrossRef]
- Mori, T.; Koyama, N.; Tan, J.; Segawa, T.; Maeda, M.; Town, T. Combination therapy with octyl gallate and ferulic acid improves cognition and neurodegeneration in a transgenic mouse model of Alzheimer’s disease. J. Biol. Chem. 2017, 292, 11310–11325. [Google Scholar] [CrossRef]
- Sassa, S.; Kikuchi, T.; Shinoda, H.; Suzuki, S.; Kudo, H.; Sakamoto, S. Preventive effect of ferulic acid on bone loss in ovariectomized rats. In Vivo 2003, 17, 277–280. [Google Scholar] [PubMed]
- Salazar-López, N.J.; Astiazarán-García, H.; González-Aguilar, G.A.; Loarca-Piña, G.; Ezquerra-Brauer, J.M.; Domínguez Avila, J.A.; Robles-Sánchez, M. Ferulic acid on glucose dysregulation, dyslipidemia, and inflammation in diet-induced obese rats: An integrated study. Nutrients 2017, 9, 675. [Google Scholar] [CrossRef]
- Bumrungpert, A.; Lilitchan, S.; Tuntipopipat, S.; Tirawanchai, N.; Komindr, S. Ferulic acid supplementation improves lipid profiles, oxidative stress, and inflammatory status in hyperlipidemic subjects: A randomized, double-blind, placebo-controlled clinical trial. Nutrients 2018, 10, 713. [Google Scholar] [CrossRef] [PubMed]
- Miyamoto, J.; Watanabe, K.; Taira, S.; Kasubuchi, M.; Li, X.; Irie, J.; Itoh, H.; Kimura, I. Barley β-glucan improves metabolic condition via short-chain fatty acids produced by gut microbial fermentation in high fat diet fed mice. PLoS ONE 2018, 13, e0196579. [Google Scholar] [CrossRef]
- Nakatani, A.; Li, X.; Miyamoto, J.; Igarashi, M.; Watanabe, H.; Sutou, A.; Watanabe, K.; Motoyama, T.; Tachibana, N.; Kohno, M.; et al. Dietary mung bean protein reduces high-fat diet-induced weight gain by modulating host bile acid metabolism in a gut microbiota-dependent manner. Biochem. Biophys. Res. Commun. 2018, 501, 955–961. [Google Scholar] [CrossRef] [PubMed]
- Renouf, M.; Guy, P.; Marmet, C.; Longet, K.; Fraering, A.L.; Moulin, J.; Barron, D.; Dionisi, F.; Cavin, C.; Steiling, H.; et al. Plasma appearance and correlation between coffee and green tea metabolites in human subjects. Br. J. Nutr. 2010, 104, 1635–1640. [Google Scholar] [CrossRef] [PubMed]
- Larrosa, M.; Luceri, C.; Vivoli, E.; Pagliuca, C.; Lodovici, M.; Moneti, G.; Dolara, P. Polyphenol metabolites from colonic microbiota exert anti-inflammatory activity on different inflammation models. Mol. Nutr. Food Res. 2009, 53, 1044–1054. [Google Scholar] [CrossRef]
- Verzelloni, E.; Pellacani, C.; Tagliazucchi, D.; Tagliaferri, S.; Calani, L.; Costa, L.G.; Brighenti, F.; Borges, G.; Crozier, A.; Conte, A.; et al. Antiglycative and neuroprotective activity of colon-derived polyphenol catabolites. Mol. Nutr. Food Res. 2011, 55, S35–S43. [Google Scholar] [CrossRef] [PubMed]
- Kimura, I.; Ozawa, K.; Inoue, D.; Imamura, T.; Kimura, K.; Maeda, T.; Terasawa, K.; Kashihara, D.; Hirano, K.; Tani, T.; et al. The gut microbiota suppresses insulin-mediated fat accumulation via the short-chain fatty acid receptor GPR43. Nat. Commun. 2013, 4, 1829. [Google Scholar] [CrossRef]
- Gogoi, B.; Chatterjee, P.; Mukherjee, S.; Buragohain, A.K.; Bhattacharya, S.; Dasgupta, S. A polyphenol rescues lipid induced insulin resistance in skeletal muscle cells and adipocytes. Biochem. Biophys. Res. Commun. 2014, 452, 382–388. [Google Scholar] [CrossRef]
- Poquet, L.; Clifford, M.N.; Williamson, G. Transport and metabolism of ferulic acid through the colonic epithelium. Drug Metab. Dispos. 2008, 36, 190–197. [Google Scholar] [CrossRef]
- Gonzalez de Mejia, E.; Ramirez-Mares, M.V. Impact of caffeine and coffee on our health. Trends Endocrinol. Metab. 2014, 25, 489–492. [Google Scholar] [CrossRef] [PubMed]
- Flint, H.J.; Scott, K.P.; Louis, P.; Duncan, S.H. The role of the gut microbiota in nutrition and health. Nat. Rev. Gastroenterol. Hepatol. 2012, 9, 577–589. [Google Scholar] [CrossRef] [PubMed]
- Claus, S.P.; Ellero, S.L.; Berger, B.; Krause, L.; Bruttin, A.; Molina, J.; Paris, A.; Want, E.J.; de Waziers, I.; Cloarec, O.; et al. Colonization-induced host-gut microbial metabolic interaction. mBio 2011, 2, e00271-10. [Google Scholar] [CrossRef] [PubMed]
- Sànchez-Maldonado, A.F.; Schieber, A.; Gänzle, M.G. Structure-function relationshipsof the antibacterial activity of phenolic acids and their metabolism by lactic acid bacteria. J. Appl. Microbiol. 2011, 111, 1176–1184. [Google Scholar] [CrossRef]
- Filannino, P.; Gobbetti, M.; De Angelis, M.; Di Cagno, R. Hydroxycinnamic acids used as external acceptors of electrons: an energetic advantage for strictly heterofermentative lactic acid bacteria. Appl. Environ. Microbiol. 2014, 80, 7574–7582. [Google Scholar] [CrossRef] [PubMed]
- Knockaert, D.; Raes, K.; Wille, C.; Struijs, K.; Van Camp, J. Metabolism of ferulic acid during growth of Lactobacillus plantarum and Lactobacillus collinoides. J. Sci. Food Agric. 2012, 92, 2291–2296. [Google Scholar] [CrossRef]
- Santamaría, L.; Reverón, I.; López de Felipe, F.; de Las Rivas, B.; Muñoz, R. Unravelling the reduction pathway as an alternative metabolic route to hydroxycinnamate decarboxylation in Lactobacillus plantarum. Appl. Environ. Microbiol. 2018, 84, e01123-18. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formula | HFD 1 | HMCA 2 |
|---|---|---|
| Product | kcal % | kcal % |
| Protein | 20 | 20 |
| Carbohydrate | 20 | 20 |
| Fat | 60 | 60 |
| Ingredient | gm | gm |
| Casein, 30 mesh | 200 | 198 |
| L-cystine | 3 | 2.97 |
| Corn starch | 0 | 0 |
| Maltodextrin 10 | 125 | 123.75 |
| Sucrose | 68.8 | 68.112 |
| Cellulose, BW200 | 50 | 49.5 |
| HMCA | 0 | 7.7385 |
| Cellulose | 0 | 0 |
| Soybean oil | 25 | 24.75 |
| Lard | 245 | 242.55 |
| Mineral mix S10026 | 10 | 9.9 |
| Dicalcium phosphate | 13 | 12.87 |
| Calcium carbonate | 5.5 | 5.445 |
| Potassium citrate, 1 H2O | 16.5 | 16.335 |
| Vitamin mix V10001 | 10 | 9.9 |
| Choline bitartrate | 2 | 1.98 |
| FD&C blue dye #1 | 0.05 | 0.0495 |
| Formula | Control 1 | HMPA 2 |
|---|---|---|
| Product | kcal % | kcal % |
| Protein | 20 | 20 |
| Carbohydrate | 20 | 20 |
| Fat | 60 | 60 |
| Ingredient | gm | gm |
| Casein, 30 mesh | 198 | 198 |
| L-cystine | 2.97 | 2.97 |
| Corn starch | 0 | 0 |
| Maltodextrin 10 | 123.75 | 123.75 |
| Sucrose | 68.112 | 68.112 |
| Cellulose, BW200 | 49.5 | 49.5 |
| HMPA | 0 | 7.7385 |
| Cellulose | 7.7385 | 0 |
| Soybean oil | 24.75 | 24.75 |
| Lard | 242.55 | 242.55 |
| Mineral mix S10026 | 9.9 | 9.9 |
| Dicalcium phosphate | 12.87 | 12.87 |
| Calcium carbonate | 5.445 | 5.445 |
| Potassium citrate, 1 H2O | 16.335 | 16.335 |
| Vitamin mix V10001 | 9.9 | 9.9 |
| Choline bitartrate | 1.98 | 1.98 |
| FD&C blue dye #1 | 0.0495 | 0.0495 |
| Gene | Primer | Sequence |
|---|---|---|
| 18S | Forward | 5′-CTCAACACGGGAAACCTCAC-3′ |
| Reverse | 5′-AGACAAATCGCTCCACCAAC-3′ | |
| Pgc1a | Forward | 5′-GAGAATGAGGCAAACTTGCTAGCG-3′ |
| Reverse | 5′-TGCATGGTTCTGAGTGCTAAGACC-3′ | |
| Acc1 | Forward | 5′-AAGGCTATGTGAAGGATG-3′ |
| Reverse | 5′-CTGTCTGAAGAGGTTAGG-3′ | |
| Cpt1a | Forward | 5′-GCATAAACGCAGAGCATTCC-3′ |
| Reverse | 5′-GATGTTGGGGTTCTTGTCTCC-3′ | |
| Cpt2 | Forward | 5′-CTCATCCGCTTTGTTCCTTC-3′ |
| Reverse | 5′-AGTTCATCACGACTGGGTTTG-3′ | |
| Cd36 | Forward | 5′-TGGCAAAGAACAGCAGCAAA-3′ |
| Reverse | 5′-GACAGTGAAGGCTCAAAGATGG-3′ | |
| Pkm2 | Forward | 5′-TCTTCTGGACCCATCGGCCCCAGGA-3′ |
| Reverse | 5′-AAAGGGATAGGGGAGGGGAAG-3′ | |
| Fas | Forward | 5′-GCTGCGGAAACTTCAGGAAAT-3′ |
| Reverse | 5′-AGAGACGTGTCACTCCTGGACTT-3′ | |
| Scd1 | Forward | 5′-GTCAGGAGGGCAGGTTTC-3′ |
| Reverse | 5′-GAGCGTGGACTTCGGTTC-3′ | |
| Gck | Forward | 5′-TACCCCTGGGCTTCACCTT-3′ |
| Reverse | 5′-CACCTGCGACACAAACGG-3′ |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ohue-Kitano, R.; Taira, S.; Watanabe, K.; Masujima, Y.; Kuboshima, T.; Miyamoto, J.; Nishitani, Y.; Kawakami, H.; Kuwahara, H.; Kimura, I. 3-(4-Hydroxy-3-methoxyphenyl)propionic Acid Produced from 4-Hydroxy-3-methoxycinnamic Acid by Gut Microbiota Improves Host Metabolic Condition in Diet-Induced Obese Mice. Nutrients 2019, 11, 1036. https://doi.org/10.3390/nu11051036
Ohue-Kitano R, Taira S, Watanabe K, Masujima Y, Kuboshima T, Miyamoto J, Nishitani Y, Kawakami H, Kuwahara H, Kimura I. 3-(4-Hydroxy-3-methoxyphenyl)propionic Acid Produced from 4-Hydroxy-3-methoxycinnamic Acid by Gut Microbiota Improves Host Metabolic Condition in Diet-Induced Obese Mice. Nutrients. 2019; 11(5):1036. https://doi.org/10.3390/nu11051036
Chicago/Turabian StyleOhue-Kitano, Ryuji, Satsuki Taira, Keita Watanabe, Yuki Masujima, Toru Kuboshima, Junki Miyamoto, Yosuke Nishitani, Hideaki Kawakami, Hiroshige Kuwahara, and Ikuo Kimura. 2019. "3-(4-Hydroxy-3-methoxyphenyl)propionic Acid Produced from 4-Hydroxy-3-methoxycinnamic Acid by Gut Microbiota Improves Host Metabolic Condition in Diet-Induced Obese Mice" Nutrients 11, no. 5: 1036. https://doi.org/10.3390/nu11051036
APA StyleOhue-Kitano, R., Taira, S., Watanabe, K., Masujima, Y., Kuboshima, T., Miyamoto, J., Nishitani, Y., Kawakami, H., Kuwahara, H., & Kimura, I. (2019). 3-(4-Hydroxy-3-methoxyphenyl)propionic Acid Produced from 4-Hydroxy-3-methoxycinnamic Acid by Gut Microbiota Improves Host Metabolic Condition in Diet-Induced Obese Mice. Nutrients, 11(5), 1036. https://doi.org/10.3390/nu11051036