Metabolic Cytokines at Fasting and During Macronutrient Challenges: Influence of Obesity, Female Androgen Excess and Sex

 ,
,  , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Subjects
2.2. Study Design
2.3. Laboratory Measurements
2.4. Multi-Analyte Profiling of Metabolic Cytokines
2.5. Sample Size Analysis
2.6. Statistical Analysis
3. Results
3.1. Effect of Group and Obesity on Clinical and Biochemical Variables in the Fasting State
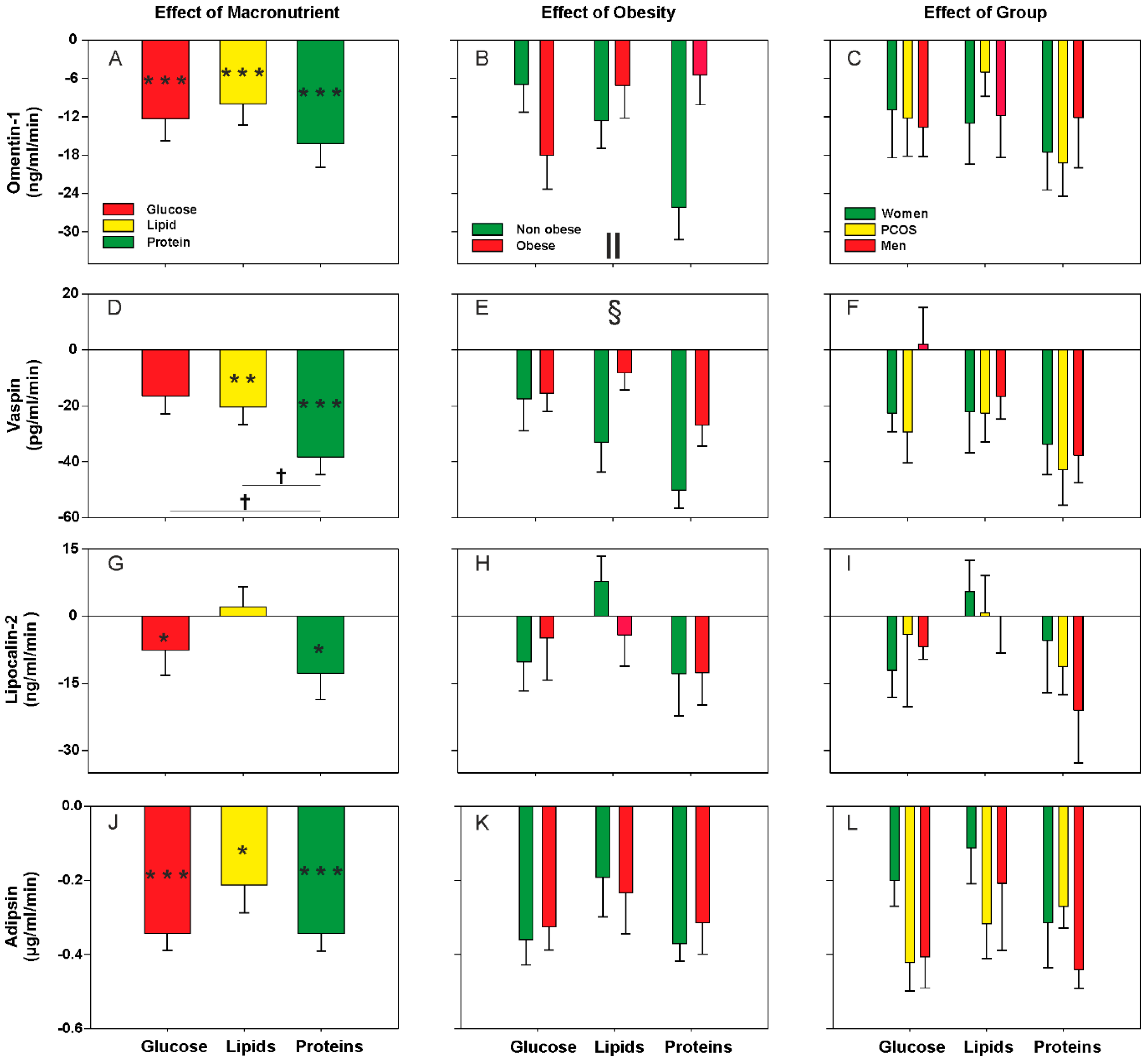
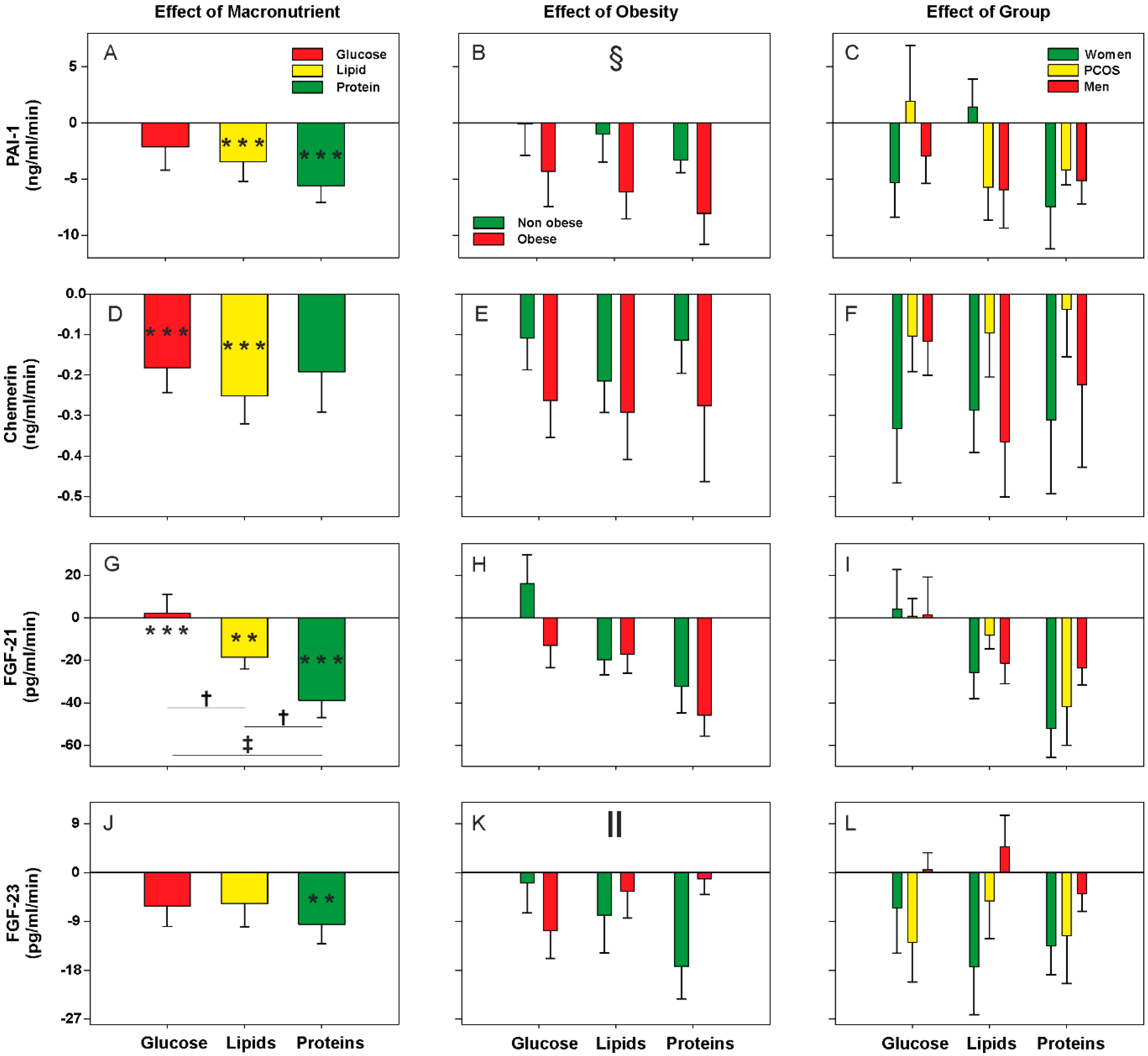
3.2. Effect of Oral Macronutrient Challenges on Metabolic Cytokine Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Petersen, P.S.; Lei, X.; Seldin, M.M.; Rodriguez, S.; Byerly, M.S.; Wolfe, A.; Whitlock, S.; Wong, G.W. Dynamic and extensive metabolic state-dependent regulation of cytokine expression and circulating levels. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2014, 307, R1458–R1470. [Google Scholar] [CrossRef] [PubMed]
- Haugen, F.; Drevon, C.A. The interplay between nutrients and the adipose tissue. Proc. Nutr. Soc. 2007, 66, 171–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calder, P.C.; Ahluwalia, N.; Brouns, F.; Buetler, T.; Clement, K.; Cunningham, K.; Esposito, K.; Jonsson, L.S.; Kolb, H.; Lansink, M.; et al. Dietary factors and low-grade inflammation in relation to overweight and obesity. Br. J. Nutr. 2011, 106, S5–S78. [Google Scholar] [CrossRef] [PubMed]
- Luque-Ramirez, M.; Martinez-Garcia, M.A.; Montes-Nieto, R.; Fernandez-Duran, E.; Insenser, M.; Alpanes, M.; Escobar-Morreale, H.F. Sexual dimorphism in adipose tissue function as evidenced by circulating adipokine concentrations in the fasting state and after an oral glucose challenge. Hum. Reprod. 2013, 28, 1908–1918. [Google Scholar] [CrossRef]
- Newell-Fugate, A.E. The role of sex steroids in white adipose tissue adipocyte function. Reproduction 2017, 153, R133–R149. [Google Scholar] [CrossRef] [Green Version]
- Escobar-Morreale, H.F. Polycystic ovary syndrome: Definition, aetiology, diagnosis and treatment. Nat. Rev. Endocrinol. 2018, 14, 270–284. [Google Scholar] [CrossRef]
- Spritzer, P.M.; Lecke, S.B.; Satler, F.; Morsch, D.M. Adipose tissue dysfunction, adipokines, and low-grade chronic inflammation in polycystic ovary syndrome. Reproduction 2015, 149, R219–R227. [Google Scholar] [CrossRef] [Green Version]
- Escobar-Morreale, H.F.; San Millan, J.L. Abdominal adiposity and the polycystic ovary syndrome. Trends Endocrinol. Metab. 2007, 18, 266–272. [Google Scholar] [CrossRef]
- Bahceci, M.; Aydemir, M.; Tuzcu, A. Effects of oral fat and glucose tolerance test on serum lipid profile, apolipoprotein, and CRP concentration, and insulin resistance in patients with polycystic ovary syndrome. Fertil. Steril. 2007, 87, 1363–1368. [Google Scholar] [CrossRef]
- Sottero, B.; Gargiulo, S.; Russo, I.; Barale, C.; Poli, G.; Cavalot, F. Postprandial Dysmetabolism and Oxidative Stress in Type 2 Diabetes: Pathogenetic Mechanisms and Therapeutic Strategies. Med. Res. Rev. 2015, 35, 968–1031. [Google Scholar] [CrossRef]
- Galland, L. Diet and inflammation. Nutr. Clin. Pr. 2010, 25, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Aljada, A.; Mohanty, P.; Ghanim, H.; Abdo, T.; Tripathy, D.; Chaudhuri, A.; Dandona, P. Increase in intranuclear nuclear factor kappaB and decrease in inhibitor kappaB in mononuclear cells after a mixed meal: Evidence for a proinflammatory effect. Am. J. Clin. Nutr. 2004, 79, 682–690. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, H.; Batra, M.; Abuaysheh, S.; Green, K.; Makdissi, A.; Kuhadiya, N.D.; Chaudhuri, A.; Dandona, P. Antiinflammatory and ROS Suppressive Effects of the Addition of Fiber to a High-Fat High-Calorie Meal. J. Clin. Endocrinol. Metab. 2017, 102, 858–869. [Google Scholar]
- Ghanim, H.; Sia, C.L.; Korzeniewski, K.; Lohano, T.; Abuaysheh, S.; Marumganti, A.; Chaudhuri, A.; Dandona, P. A resveratrol and polyphenol preparation suppresses oxidative and inflammatory stress response to a high-fat, high-carbohydrate meal. J. Clin. Endocrinol. Metab. 2011, 96, 1409–1414. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, H.; Sia, C.L.; Upadhyay, M.; Korzeniewski, K.; Viswanathan, P.; Abuaysheh, S.; Mohanty, P.; Dandona, P. Orange juice neutralizes the proinflammatory effect of a high-fat, high-carbohydrate meal and prevents endotoxin increase and Toll-like receptor expression. Am. J. Clin. Nutr. 2010, 91, 940–949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Comitato, R.; Saba, A.; Turrini, A.; Arganini, C.; Virgili, F. Sex hormones and macronutrient metabolism. Crit. Rev. Food Sci. Nutr. 2014, 55, 227–241. [Google Scholar] [CrossRef]
- Kasim-Karakas, S.E.; Cunningham, W.M.; Tsodikov, A. Relation of nutrients and hormones in polycystic ovary syndrome. Am. J. Clin. Nutr. 2007, 85, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Aljada, A.; Ghanim, H.; Mohanty, P.; Syed, T.; Bandyopadhyay, A.; Dandona, P. Glucose intake induces an increase in activator protein 1 and early growth response 1 binding activities, in the expression of tissue factor and matrix metalloproteinase in mononuclear cells, and in plasma tissue factor and matrix metalloproteinase concentrations. Am. J. Clin. Nutr. 2004, 80, 51–57. [Google Scholar] [Green Version]
- Deopurkar, R.; Ghanim, H.; Friedman, J.; Abuaysheh, S.; Sia, C.L.; Mohanty, P.; Viswanathan, P.; Chaudhuri, A.; Dandona, P. Differential effects of cream, glucose, and orange juice on inflammation, endotoxin, and the expression of Toll-like receptor-4 and suppressor of cytokine signaling-3. Diabetes Care 2010, 33, 991–997. [Google Scholar] [CrossRef]
- Mohanty, P.; Ghanim, H.; Hamouda, W.; Aljada, A.; Garg, R.; Dandona, P. Both lipid and protein intakes stimulate increased generation of reactive oxygen species by polymorphonuclear leukocytes and mononuclear cells. Am. J. Clin. Nutr. 2002, 75, 767–772. [Google Scholar] [CrossRef] [Green Version]
- Mohanty, P.; Hamouda, W.; Garg, R.; Aljada, A.; Ghanim, H.; Dandona, P. Glucose challenge stimulates reactive oxygen species (ROS) generation by leucocytes. J. Clin. Endocrinol. Metab. 2000, 85, 2970–2973. [Google Scholar] [CrossRef] [PubMed]
- Tripathy, D.; Mohanty, P.; Dhindsa, S.; Syed, T.; Ghanim, H.; Aljada, A.; Dandona, P. Elevation of free fatty acids induces inflammation and impairs vascular reactivity in healthy subjects. Diabetes 2003, 52, 2882–2887. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, F.; Considine, R.V.; Abdelhadi, O.A.; Acton, A.J. Saturated fat ingestion promotes lipopolysaccharide-mediated inflammation and insulin resistance in PCOS. J. Clin. Endocrinol. Metab. 2019, 104, 934–936. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, F.; Sia, C.L.; Shepard, M.K.; Rote, N.S.; Minium, J. Hyperglycemia-induced oxidative stress is independent of excess abdominal adiposity in normal-weight women with polycystic ovary syndrome. Hum. Reprod. 2012, 27, 3560–3568. [Google Scholar] [CrossRef] [Green Version]
- Gonzalez, F.; Sia, C.L.; Shepard, M.K.; Rote, N.S.; Minium, J. Inflammation in response to glucose ingestion is independent of excess abdominal adiposity in normal-weight women with polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2012, 97, 4071–4079. [Google Scholar] [CrossRef]
- Gonzalez, F.; Sia, C.L.; Shepard, M.K.; Rote, N.S.; Minium, J. The altered mononuclear cell-derived cytokine response to glucose ingestion is not regulated by excess adiposity in polycystic ovary syndrome. J. Clin. Endocrinol. Metab. 2014, 99, E2244–E2251. [Google Scholar] [CrossRef]
- Martinez-Garcia, M.A.; Moncayo, S.; Insenser, M.; Montes-Nieto, R.; Fernandez-Duran, E.; Alvarez-Blasco, F.; Luque-Ramirez, M.; Escobar-Morreale, H.F. Postprandial inflammatory responses after oral glucose, lipid and protein challenges: Influence of obesity, sex and polycystic ovary syndrome. Clin. Nutr. 2019. [Google Scholar] [CrossRef]
- Montes-Nieto, R.; Insenser, M.; Murri, M.; Fernandez-Duran, E.; Ojeda-Ojeda, M.; Martinez-Garcia, M.A.; Luque-Ramirez, M.; Escobar-Morreale, H.F. Plasma thiobarbituric acid reactive substances (TBARS) in young adults: Obesity increases fasting levels only in men whereas glucose ingestion, and not protein or lipid intake, increases postprandial concentrations regardless of sex and obesity. Mol. Nutr. Food Res. 2017, 61. [Google Scholar] [CrossRef]
- Zawadzki JK, D.A. Diagnostic criteria for polycystic ovary syndrome: Towards a rational approach. In Polycystic Ovary Syndrome; Dunaif, A., Givens, J.R., Haseltine, F.P., Merriam, G.R., Eds.; Blackwell Science Publishing: Boston, MA, USA, 1992; pp. 377–384. [Google Scholar]
- Ankarberg-Lindgren, C.; Norjavaara, E. Sensitive RIA measures testosterone concentrations in prepubertal and pubertal children comparable to tandem mass spectrometry. Scand. J. Clin. Lab. Invest. 2015, 75, 341–344. [Google Scholar] [CrossRef]
- Escobar-Morreale, H.F.; Sanchon, R.; San Millan, J.L. A prospective study of the prevalence of nonclassical congenital adrenal hyperplasia among women presenting with hyperandrogenic symptoms and signs. J. Clin. Endocrinol. Metab. 2008, 93, 527–533. [Google Scholar] [CrossRef]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Matsuda, M.; DeFronzo, R.A. Insulin sensitivity indices obtained from oral glucose tolerance testing: Comparison with the euglycemic insulin clamp. Diabetes Care 1999, 22, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, T.; Watanabe-Kominato, K.; Takahashi, Y.; Kojima, M.; Watanabe, R. Adipose Tissue-Derived Omentin-1 Function and Regulation. Compr. Physiol. 2017, 7, 765–781. [Google Scholar] [PubMed]
- Dimova, R.; Tankova, T. The role of vaspin in the development of metabolic and glucose tolerance disorders and atherosclerosis. Biomed. Res. Int. 2015, 2015, 823481. [Google Scholar] [CrossRef] [PubMed]
- Helfer, G.; Wu, Q.F. Chemerin: A multifaceted adipokine involved in metabolic disorders. J. Endocrinol. 2018, 238, R79–R94. [Google Scholar] [CrossRef]
- Moschen, A.R.; Adolph, T.E.; Gerner, R.R.; Wieser, V.; Tilg, H. Lipocalin-2: A Master Mediator of Intestinal and Metabolic Inflammation. Trends Endocrinol. Metab. 2017, 28, 388–397. [Google Scholar] [CrossRef]
- Lo, J.C.; Ljubicic, S.; Leibiger, B.; Kern, M.; Leibiger, I.B.; Moede, T.; Kelly, M.E.; Chatterjee Bhowmick, D.; Murano, I.; Cohen, P.; et al. Adipsin is an adipokine that improves beta cell function in diabetes. Cell 2014, 158, 41–53. [Google Scholar] [CrossRef]
- Kaji, H. Adipose Tissue-Derived Plasminogen Activator Inhibitor-1 Function and Regulation. Compr. Physiol. 2016, 6, 1873–1896. [Google Scholar]
- Kharitonenkov, A.; DiMarchi, R. Fibroblast growth factor 21 night watch: Advances and uncertainties in the field. J. Intern. Med. 2017, 281, 233–246. [Google Scholar] [CrossRef]
- Vervloet, M. Renal and extrarenal effects of fibroblast growth factor 23. Nat. Rev. Nephrol. 2019, 15, 109–120. [Google Scholar] [CrossRef]
- Romano, M.; Guagnano, M.T.; Pacini, G.; Vigneri, S.; Falco, A.; Marinopiccoli, M.; Manigrasso, M.R.; Basili, S.; Davi, G. Association of inflammation markers with impaired insulin sensitivity and coagulative activation in obese healthy women. J. Clin. Endocrinol. Metab. 2003, 88, 5321–5326. [Google Scholar] [CrossRef] [PubMed]
- Carmassi, F.; De Negri, F.; Fioriti, R.; De Giorgi, A.; Giannarelli, C.; Fruzzetti, F.; Pedrinelli, R.; Dell’Omo, G.; Bersi, C. Insulin resistance causes impaired vasodilation and hypofibrinolysis in young women with polycystic ovary syndrome. Thromb. Res. 2005, 116, 207–214. [Google Scholar] [CrossRef] [PubMed]
- Dandona, P.; Mohanty, P.; Ghanim, H.; Aljada, A.; Browne, R.; Hamouda, W.; Prabhala, A.; Afzal, A.; Garg, R. The suppressive effect of dietary restriction and weight loss in the obese on the generation of reactive oxygen species by leukocytes, lipid peroxidation, and protein carbonylation. J. Clin. Endocrinol. Metab. 2001, 86, 355–362. [Google Scholar] [PubMed]
- Dandona, P.; Weinstock, R.; Thusu, K.; Abdel-Rahman, E.; Aljada, A.; Wadden, T. Tumor necrosis factor-alpha in sera of obese patients: Fall with weight loss. J. Clin. Endocrinol. Metab. 1998, 83, 2907–2910. [Google Scholar] [PubMed]
- Ghanim, H.; Aljada, A.; Daoud, N.; Deopurkar, R.; Chaudhuri, A.; Dandona, P. Role of inflammatory mediators in the suppression of insulin receptor phosphorylation in circulating mononuclear cells of obese subjects. Diabetologia 2007, 50, 278–285. [Google Scholar] [CrossRef] [PubMed]
- Ghanim, H.; Aljada, A.; Hofmeyer, D.; Syed, T.; Mohanty, P.; Dandona, P. Circulating mononuclear cells in the obese are in a proinflammatory state. Circulation 2004, 110, 1564–1571. [Google Scholar] [CrossRef]
- Monte, S.V.; Caruana, J.A.; Ghanim, H.; Sia, C.L.; Korzeniewski, K.; Schentag, J.J.; Dandona, P. Reduction in endotoxemia, oxidative and inflammatory stress, and insulin resistance after Roux-en-Y gastric bypass surgery in patients with morbid obesity and type 2 diabetes mellitus. Surgery 2012, 151, 587–593. [Google Scholar] [CrossRef]
- Deng, Y.; Scherer, P.E. Adipokines as novel biomarkers and regulators of the metabolic syndrome. Ann. N. Y. Acad. Sci. 2011, 1212, E1–E19. [Google Scholar] [CrossRef]
- Korner, A.; Neef, M.; Friebe, D.; Erbs, S.; Kratzsch, J.; Dittrich, K.; Bluher, S.; Kapellen, T.M.; Kovacs, P.; Stumvoll, M.; et al. Vaspin is related to gender, puberty and deteriorating insulin sensitivity in children. Int. J. Obes. (Lond.) 2011, 35, 578–586. [Google Scholar] [CrossRef]
- Ali, F.N.; Falkner, B.; Gidding, S.S.; Price, H.E.; Keith, S.W.; Langman, C.B. Fibroblast growth factor-23 in obese, normotensive adolescents is associated with adverse cardiac structure. J. Pediatr. 2014, 165, 738–743. [Google Scholar] [CrossRef]
- Hu, X.; Ma, X.; Luo, Y.; Xu, Y.; Xiong, Q.; Pan, X.; Xiao, Y.; Bao, Y.; Jia, W. Associations of serum fibroblast growth factor 23 levels with obesity and visceral fat accumulation. Clin. Nutr. 2018, 37, 223–228. [Google Scholar] [CrossRef]
- Holecki, M.; Chudek, J.; Owczarek, A.; Olszanecka-Glinianowicz, M.; Bozentowicz-Wikarek, M.; Dulawa, J.; Mossakowska, M.; Zdrojewski, T.; Skalska, A.; Wiecek, A. Inflammation but not obesity or insulin resistance is associated with increased plasma fibroblast growth factor 23 concentration in the elderly. Clin. Endocrinol. (Oxf.) 2015, 82, 900–909. [Google Scholar] [CrossRef] [PubMed]
- Wojcik, M.; Janus, D.; Dolezal-Oltarzewska, K.; Drozdz, D.; Sztefko, K.; Starzyk, J.B. The association of FGF23 levels in obese adolescents with insulin sensitivity. J. Pediatr. Endocrinol. Metab. 2012, 25, 687–690. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Real, J.M.; Puig, J.; Serrano, M.; Sabater, M.; Rubio, A.; Moreno-Navarrete, J.M.; Fontan, M.; Casamitjana, R.; Xifra, G.; Ortega, F.J.; et al. Iron and obesity status-associated insulin resistance influence circulating fibroblast-growth factor-23 concentrations. PLoS ONE 2013, 8, e58961. [Google Scholar] [CrossRef] [PubMed]
- Gursoy Calan, O.; Calan, M.; Yesil Senses, P.; Unal Kocabas, G.; Ozden, E.; Sari, K.R.; Kocar, M.; Imamoglu, C.; Senses, Y.M.; Bozkaya, G.; et al. Increased adipsin is associated with carotid intima media thickness and metabolic disturbances in polycystic ovary syndrome. Clin. Endocrinol. (Oxf.) 2016, 85, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Kahraman, S.; Altinova, A.E.; Yalcin, M.M.; Gulbahar, O.; Arslan, B.; Akturk, M.; Cakir, N.; Toruner, F.B. Association of serum betatrophin with fibroblast growth factor-21 in women with polycystic ovary syndrome. J. Endocrinol. Investig. 2018, 41, 1069–1074. [Google Scholar] [CrossRef]
- Olszanecka-Glinianowicz, M.; Madej, P.; Wdowczyk, M.; Owczarek, A.; Chudek, J. Circulating FGF21 levels are related to nutritional status and metabolic but not hormonal disturbances in polycystic ovary syndrome. Eur. J. Endocrinol. 2015, 172, 173–179. [Google Scholar] [CrossRef] [Green Version]
- Sahin, S.B.; Ayaz, T.; Cure, M.C.; Sezgin, H.; Ural, U.M.; Balik, G.; Sahin, F.K. Fibroblast growth factor 21 and its relation to metabolic parameters in women with polycystic ovary syndrome. Scand. J. Clin. Lab. Investig. 2014, 74, 465–469. [Google Scholar] [CrossRef]
- Vejrazkova, D.; Lischkova, O.; Vankova, M.; Stanicka, S.; Vrbikova, J.; Lukasova, P.; Vcelak, J.; Vacinova, G.; Bendlova, B. Distinct response of fat and gastrointestinal tissue to glucose in gestational diabetes mellitus and polycystic ovary syndrome. Physiol. Res. 2017, 66, 283–292. [Google Scholar]
- Cakal, E.; Ozkaya, M.; Engin-Ustun, Y.; Ustun, Y. Serum lipocalin-2 as an insulin resistance marker in patients with polycystic ovary syndrome. J. Endocrinol. Investig. 2011, 34, 97–100. [Google Scholar]
- Diamanti-Kandarakis, E.; Livadas, S.; Kandarakis, S.A.; Margeli, A.; Papassotiriou, I. Serum concentrations of atherogenic proteins neutrophil gelatinase-associated lipocalin and its complex with matrix metalloproteinase-9 are significantly lower in women with polycystic ovary syndrome: Hint of a protective mechanism? Eur. J. Endocrinol. 2008, 158, 525–531. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Garcia, M.A.; Montes-Nieto, R.; Fernandez-Duran, E.; Insenser, M.; Luque-Ramirez, M.; Escobar-Morreale, H.F. Evidence for masculinization of adipokine gene expression in visceral and subcutaneous adipose tissue of obese women with polycystic ovary syndrome (PCOS). J. Clin. Endocrinol. Metab. 2013, 98, E388–E396. [Google Scholar] [CrossRef] [PubMed]
- Huang, R.; Yue, J.; Sun, Y.; Zheng, J.; Tao, T.; Li, S.; Liu, W. Increased serum chemerin concentrations in patients with polycystic ovary syndrome: Relationship between insulin resistance and ovarian volume. Clin. Chim. Acta. 2015, 450, 366–369. [Google Scholar] [CrossRef] [PubMed]
- Koiou, E.; Dinas, K.; Tziomalos, K.; Toulis, K.; Kandaraki, E.A.; Kalaitzakis, E.; Katsikis, I.; Panidis, D. The phenotypes of polycystic ovary syndrome defined by the 1990 diagnostic criteria are associated with higher serum vaspin levels than the phenotypes introduced by the 2003 criteria. Obes. Facts 2011, 4, 145–150. [Google Scholar] [CrossRef]
- Tang, Y.L.; Yu, J.; Zeng, Z.G.; Liu, Y.; Liu, J.Y.; Xu, J.X. Circulating omentin-1 levels in women with polycystic ovary syndrome: A meta-analysis. Gynecol. Endocrinol. 2017, 33, 244–249. [Google Scholar] [CrossRef]
- Toulis, K.A.; Goulis, D.G.; Mintziori, G.; Kintiraki, E.; Eukarpidis, E.; Mouratoglou, S.A.; Pavlaki, A.; Stergianos, S.; Poulasouchidou, M.; Tzellos, T.G.; et al. Meta-analysis of cardiovascular disease risk markers in women with polycystic ovary syndrome. Hum. Reprod. Update 2011, 17, 741–760. [Google Scholar] [CrossRef] [Green Version]
- Akbarzadeh, S.; Ghasemi, S.; Kalantarhormozi, M.; Nabipour, I.; Abbasi, F.; Aminfar, A.; Jaffari, S.M.; Motamed, N.; Movahed, A.; Mirzaei, M.; et al. Relationship among plasma adipokines, insulin and androgens level as well as biochemical glycemic and lipidemic markers with incidence of PCOS in women with normal BMI. Gynecol. Endocrinol. 2012, 28, 521–524. [Google Scholar] [CrossRef]
- Cassar, S.; Teede, H.J.; Harrison, C.L.; Joham, A.E.; Moran, L.J.; Stepto, N.K. Biomarkers and insulin sensitivity in women with Polycystic Ovary Syndrome: Characteristics and predictive capacity. Clin. Endocrinol. (Oxf.) 2015, 83, 50–58. [Google Scholar] [CrossRef]
- de Mendonca-Louzeiro, M.R.; Annichino-Bizzacchi, J.M.; Benetti-Pinto, C.L. Android fat distribution affects some hemostatic parameters in women with polycystic ovary syndrome compared with healthy control subjects matched for age and body mass index. Fertil. Steril. 2015, 104, 467–473. [Google Scholar] [CrossRef]
- Guvenc, Y.; Var, A.; Goker, A.; Kuscu, N.K. Assessment of serum chemerin, vaspin and omentin-1 levels in patients with polycystic ovary syndrome. J. Int. Med. Res. 2016, 44, 796–805. [Google Scholar] [CrossRef] [Green Version]
- Tarkun, I.; Canturk, Z.; Arslan, B.C.; Turemen, E.; Tarkun, P. The plasminogen activator system in young and lean women with polycystic ovary syndrome. Endocr. J. 2004, 51, 467–472. [Google Scholar] [CrossRef] [PubMed]
- Paton, C.M.; Rogowski, M.P.; Kozimor, A.L.; Stevenson, J.L.; Chang, H.; Cooper, J.A. Lipocalin-2 increases fat oxidation in vitro and is correlated with energy expenditure in normal weight but not obese women. Obesity (Silver Spring) 2013, 21, E640–E648. [Google Scholar] [CrossRef] [PubMed]
- Tholstrup, T.; Miller, G.J.; Bysted, A.; Sandstrom, B. Effect of individual dietary fatty acids on postprandial activation of blood coagulation factor VII and fibrinolysis in healthy young men. Am. J. Clin. Nutr. 2003, 77, 1125–1132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teng, K.T.; Chang, L.F.; Vethakkan, S.R.; Nesaretnam, K.; Sanders, T.A.B. Effects of exchanging carbohydrate or monounsaturated fat with saturated fat on inflammatory and thrombogenic responses in subjects with abdominal obesity: A randomized controlled trial. Clin. Nutr. 2017, 36, 1250–1258. [Google Scholar] [CrossRef] [Green Version]
- Mortensen, L.S.; Thomsen, C.; Hermansen, K. Effects of different protein sources on plasminogen inhibitor-1 and factor VII coagulant activity added to a fat-rich meal in type 2 diabetes. Rev. Diabet. Stud. 2010, 7, 233–240. [Google Scholar]
- Gaal, S.; Kerr, M.A.; Ward, M.; McNulty, H.; Livingstone, M.B.E. Breakfast Consumption in the UK: Patterns, Nutrient Intake and Diet Quality. A Study from the International Breakfast Research Initiative Group. Nutrients 2018, 10, 999. [Google Scholar] [CrossRef]
- Barr, S.I.; Vatanparast, H.; Smith, J. Breakfast in Canada: Prevalence of consumption, contribution to nutrient and food group intakes, and variability across tertiles of daily diet quality. A study from the International Breakfast Research Initiative. Nutrients 2018, 10, 985. [Google Scholar] [CrossRef]
- Moreira, A.P.; Texeira, T.F.; Ferreira, A.B.; Peluzio Mdo, C.; Alfenas Rde, C. Influence of a high-fat diet on gut microbiota, intestinal permeability and metabolic endotoxaemia. Br. J. Nutr. 2012, 108, 801–809. [Google Scholar] [CrossRef]
- Zietek, T.; Rath, E. Inflammation Meets Metabolic Disease: Gut Feeling Mediated by GLP-1. Front. Immunol. 2016, 7, 154. [Google Scholar] [CrossRef] [Green Version]
- Schmid, A.; Leszczak, S.; Ober, I.; Karrasch, T.; Schaffler, A. Short-term and divergent regulation of FGF-19 and FGF-21 during oral lipid tolerance test but not oral glucose tolerance test. Exp. Clin. Endocrinol. Diabetes 2015, 123, 88–94. [Google Scholar] [CrossRef]
- Maruyama, R.; Shimizu, M.; Hashidume, T.; Inoue, J.; Itoh, N.; Sato, R. FGF21 Alleviates Hepatic Endoplasmic Reticulum Stress under Physiological Conditions. J. Nutr. Sci. Vitam. (Tokyo) 2018, 64, 200–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matikainen, N.; Taskinen, M.R.; Stennabb, S.; Lundbom, N.; Hakkarainen, A.; Vaaralahti, K.; Raivio, T. Decrease in circulating fibroblast growth factor 21 after an oral fat load is related to postprandial triglyceride-rich lipoproteins and liver fat. Eur. J. Endocrinol. 2012, 166, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Westerink, J.; Hajer, G.R.; Kranendonk, M.E.; Schipper, H.S.; Monajemi, H.; Kalkhoven, E.; Visseren, F.L. An oral mixed fat load is followed by a modest anti-inflammatory adipocytokine response in overweight patients with metabolic syndrome. Lipids 2014, 49, 247–254. [Google Scholar] [CrossRef] [PubMed]
- Jeong, E.; Youn, B.S.; Kim, D.W.; Kim, E.H.; Park, J.W.; Namkoong, C.; Jeong, J.Y.; Yoon, S.Y.; Park, J.Y.; Lee, K.U.; et al. Circadian rhythm of serum vaspin in healthy male volunteers: Relation to meals. J. Clin. Endocrinol. Metab. 2010, 95, 1869–1875. [Google Scholar] [CrossRef]
- van Nierop, F.S.; Scheltema, M.J.; Eggink, H.M.; Pols, T.W.; Sonne, D.P.; Knop, F.K.; Soeters, M.R. Clinical relevance of the bile acid receptor TGR5 in metabolism. Lancet Diabetes Endocrinol. 2017, 5, 224–233. [Google Scholar] [CrossRef]
- Chabot, F.; Caron, A.; Laplante, M.; St-Pierre, D.H. Interrelationships between ghrelin, insulin and glucose homeostasis: Physiological relevance. World J. Diabetes 2014, 5, 328–341. [Google Scholar] [CrossRef]
- Fritsche, K.L. The science of fatty acids and inflammation. Adv. Nutr. 2015, 6, 293S–301S. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Control Women | Women with PCOS (Polycystic Ovary Syndrome) | Control Men | Group | Obesity | Interaction | ||||
|---|---|---|---|---|---|---|---|---|---|
| Non-obese (n = 9) | Obese (n = 8) | Non-obese (n = 9) | Obese (n = 8) | Non-obese (n = 10) | Obese (n = 9) | P | P | P | |
| Age (years) | 26 ± 5 | 27 ± 6 | 24 ± 8 | 30 ± 4 | 24 ± 5 | 25 ± 4 | 0.342 | 0.101 | 0.319 |
| Body mass index (kg/m2) | 23 ± 2 | 36 ± 4 | 24 ± 2 | 37 ± 5 | 23 ± 2 | 34 ± 3 | 0.226 | <0.001 | 0.782 |
| Waist circumference (cm) | 76 ± 9 | 100 ± 17 | 72 ± 7 | 105 ± 11 | 81 ± 5 | 110 ± 13 | 0.049 B,C | <0.001 | 0.377 |
| Waist to hip ratio | 0.75 ± 0.08 | 0.83 ± 0.12 | 0.73 ± 0.05 | 0.85 ± 0.06 | 0.83 ± 0.04 | 0.90 ± 0.05 | 0.002 B, C | <0.001 | 0.436 |
| Hirsutism score | 1.4 ± 1.3 | 1.8 ± 1.2 | 9.7 ± 4.5 | 9.3 ± 4.5 | - | - | <0.001 A | 0.738 | 0.876 |
| Total testosterone (ng/dL) | 46 ± 9 | 58 ± 14 | 72 ± 20 | 69 ± 29 | 534 ± 96 | 499 ± 104 | <0.001 A,B,C | 0.776 | 0.196 |
| Free testosterone (ng/dL) | 0.6 ± 0.2 | 0.9 ± 0.0 | 1.0 ± 0.4 | 1.3 ± 0.7 | 13.0 ± 3.0 | 13.4 ± 2.7 | <0.001 A,B,C | 0.024 | 0.265 |
| Total estradiol (pg/mL) | 41 ± 17 | 75 ± 55 | 50 ± 55 | 41 ± 13 | 19 ± 4 | 26 ± 7 | <0.001 B,C | 0.024 | 0.422 |
| Free estradiol (pg/mL) | 0.74 ± 0.30 | 1.44 ± 0.82 | 0.98 ± 1.17 | 0.93 ± 0.38 | 0.49 ± 0.14 | 0.71 ± 0.19 | 0.010 B | 0.003 | 0.520 |
| Ratio FT/FE2 | 8.6 ± 0.9 | 7.6 ± 1.6 | 17.4 ± 4.0 | 14.0 ± 1.9 | 263.4 ± 22.6 | 186.4 ± 12.7 | <0.001 A,B,C | 0.119 | 0.750 |
| SHBG (µg/dL) | 504 ± 225 | 387 ± 126 | 450 ± 189 | 288 ± 117 | 243 ± 90 | 180 ± 54 | <0.001 B,C | 0.008 | 0.568 |
| Androstendione (ng/mL) | 2.6 ± 0.8 | 2.7 ± 0.8 | 4.2 ± 1.2 | 3.8 ± 1.8 | 2.0 ± 0.5 | 2.7 ± 1.1 | <0.001 A,C | 0.712 | 0.371 |
| DHEAS (ng/mL) | 1990 ± 442 | 1953 ± 553 | 2800 ± 1437 | 2100 ± 774 | 2211 ± 663 | 3021 ± 1068 | 0.094 | 0.852 | 0.081 |
| hsCRP (mg/L) | 2.8 ± 2.3 | 4.0 ± 2.9 | 2.1 ± 2.2 | 6.8 ± 7.7 | 3.3 ± 2.6 | 3.2 ± 1.3 | 0.998 | 0.009 | 0.207 |
| Fasting insulin (µIU/mL) | 7.9 ± 2.9 | 11.1 ± 3.0 | 7.5 ± 4.2 | 13.5 ± 3.6 | 5.8 ± 1.6 | 10.8 ± 3.9 | 0.123 | <0.001 | 0.440 |
| Fasting glucose (mg/dL) | 85 ± 7 | 96 ± 7 | 81 ± 9 | 87 ± 9 | 88 ± 9 | 94 ± 7 | 0.011 A,C | 0.001 | 0.408 |
| HOMA-IR | 1.6 ± 0.6 | 2.6 ± 0.7 | 1.5 ± 0.9 | 2.9 ± 0.7 | 1.3 ± 0.4 | 2.5 ± 1.0 | 0.424 | <0.001 | 0.742 |
| Insulin sensitivity index | 6.6 ± 2.9 | 3.3 ± 1.2 | 8.1 ± 4.7 | 3.5 ± 1.4 | 7.3 ± 2.8 | 3.8 ± 1.6 | 0.640 | <0.001 | 0.909 |
| Triglycerides (mg/dL) | 75 ± 32 | 81 ± 30 | 80 ± 22 | 101 ± 42 | 79 ± 26 | 102 ± 35 | 0.399 | 0.054 | 0.711 |
| Total cholesterol (mg/dL) | 170 ± 35 | 181 ± 35 | 166± 39 | 170 ± 35 | 158 ± 23 | 185 ± 35 | 0.844 | 0.133 | 0.485 |
| HDL-cholesterol (mg/dL) | 54 ± 12 | 51 ± 8 | 54 ± 8 | 46 ± 8 | 46 ± 8 | 39 ± 4 | 0.001 B, C | 0.002 | 0.512 |
| LDL-cholesterol (mg/dL) | 104 ± 31 | 112 ± 23 | 97 ± 31 | 104 ± 31 | 93 ± 15 | 127 ± 31 | 0.507 | 0.034 | 0.237 |
| Control Women | Women with PCOS | Control Men | Group | Obesity | Interaction | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| Non-obese (n = 9) | Obese (n = 8) | Non-obese (n = 9) | Obese (n = 8) | Non-obese (n = 9) | Obese (n = 9) | P | P | P | ||
| Omentin-1 (ng/mL) | 215 ± 48 | 201 ± 75 | 211 ± 45 | 196 ± 86 | 259 ± 47 | 201 ± 89 | 0.573 | 0.044 | 0.561 | |
| Vaspin (pg/mL) | 324 ± 159 | 249 ± 185 | 349 ± 482 | 278 ± 180 | 169 ± 123 | 133 ± 61 | 0.022 A,B | 0.472 | 0.672 | |
| Lipocalin-2 (ng/mL) | 108 ± 23 | 187 ± 72 | 180 ± 95 | 150 ± 65 | 122 ± 54 | 121 ± 29 | 0.152 | 0.203 | 0.049 | |
| Adipsin (µg/mL) | 2.87 ± 0.51 | 3.46 ± 0.50 | 3.42 ± 0.89 | 3.58 ± 0.56 | 3.20 ± 0.46 | 3.47 ± 0.84 | 0.359 | 0.054 | 0.533 | |
| PAI-1 (ng/mL) | 28.2 ± 15.9 | 60.0 ± 13.8 | 33.2 ± 15.0 | 48.8 ± 22.8 | 21.2 ± 14.6 | 44.4 ± 26.6 | 0.093 | < 0.001 | 0.424 | |
| Chemerin (ng/mL) | 8.14 ± 1.54 | 10.48 ± 2.02 | 8.49 ± 1.91 | 9.32 ± 1.30 | 8.13 ± 1.73 | 8.53 ± 1.60 | 0.240 | 0.015 | 0.224 | |
| FGF-21 (pg/mL) | 392 ± 169 | 338 ± 144 | 285 ± 78 | 349 ± 84 | 258 ± 128 | 308 ± 179 | 0.102 | 0.489 | 0.346 | |
| FGF-23 (pg/mL) | 312 ± 172 | 187 ± 41 | 237 ± 134 | 195 ± 45 | 222 ± 111 | 171 ± 19 | 0.282 | 0.013 | 0.394 | |
| Macronutrient | Omentin-1 | Vaspin | Lipocalin-2 | Adipsin | PAI-1 | Chemerin | FGF-21 | FGF-23 | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| r | p | r | p | r | p | r | p | r | p | r | p | r | p | r | p | |
| All subjects as a whole | ||||||||||||||||
| Glucose | −0.359 | 0.009 | −0.573 | <0.001 | −0.319 | 0.021 | −0.612 | <0.001 | −0.567 | <0.001 | −0.379 | 0.006 | −0.272 | 0.051 | −0.395 | 0.004 |
| Lipids | −0.456 | 0.001 | -0.505 | <0.001 | −0.249 | 0.076 | −0.661 | <0.001 | −0.426 | 0.002 | −0.430 | 0.001 | - | - | −0.567 | <0.001 |
| Proteins | - | - | -0.594 | <0.001 | −0.460 | 0.001 | −0.512 | <0.001 | −0.594 | <0.001 | −0.284 | 0.041 | −0.437 | 0.001 | −0.349 | 0.011 |
| Non-obese subjects | ||||||||||||||||
| Glucose | - | - | −0.615 | 0.001 | - | - | −0.751 | <0.001 | −0.490 | 0.009 | - | - | - | - | −0.346 | 0.077 |
| Lipids | −0.589 | 0.001 | −0.688 | <0.001 | - | - | −0.636 | <0.001 | - | - | −0.514 | 0.006 | - | - | −0.638 | <0.001 |
| Proteins | −0.473 | 0.013 | −0.734 | <0.001 | −0.330 | 0.092 | −0.419 | 0.030 | - | - | - | - | −0.400 | 0.039 | −0.563 | 0.002 |
| Obese subjects | ||||||||||||||||
| Glucose | −0.456 | 0.022 | −0.590 | 0.002 | −0.455 | 0.022 | −0.418 | 0.037 | −0.657 | <0.001 | −0.528 | 0.007 | −0.536 | 0.006 | −0.702 | <0.001 |
| Lipids | - | - | - | - | −0.604 | 0.001 | −0.651 | <0.001 | −0.502 | 0.011 | −0.370 | 0.069 | - | - | −0.398 | 0.049 |
| Proteins | 0.494 | 0.012 | −0.579 | 0.002 | −0.598 | 0.002 | −0.633 | 0.001 | −0.733 | <0.001 | −0.369 | 0.069 | −0.476 | 0.016 | - | - |
| Serum Concentrations | Fasting | Postprandial (AUC) |
|---|---|---|
| Omentin-1 | ↓ in obese | ↓ after all macronutrient loads (mainly in non-obese after proteins) |
| Vaspin | ↓ in men | ↓ during lipid challenge and ↓↓ during protein load Global effect of obesity (smaller response in obese) |
| Lipocalin-2 | ↑ in obese only in control women PCOS > control women only in non-obese | ↓ during glucose and protein loads |
| Adipsin | Trends towards ↑ in obese | ↓ after all macronutrient loads |
| PAI-1 | ↑↑ in obese | ↓ during lipid challenge and ↓↓ during protein load Global effect of obesity (larger response in obese) |
| Chemerin | ↑ in obese | ↓ during glucose and lipid challenges |
| FGF-21 | - | ↑ during glucose, ↓ during lipid challenge and ↓↓ during protein loads |
| FGF-23 | ↓ in obese | ↓ during protein load, only in non-obese individuals |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martínez-García, M.Á.; Moncayo, S.; Insenser, M.; Álvarez-Blasco, F.; Luque-Ramírez, M.; Escobar-Morreale, H.F. Metabolic Cytokines at Fasting and During Macronutrient Challenges: Influence of Obesity, Female Androgen Excess and Sex. Nutrients 2019, 11, 2566. https://doi.org/10.3390/nu11112566
Martínez-García MÁ, Moncayo S, Insenser M, Álvarez-Blasco F, Luque-Ramírez M, Escobar-Morreale HF. Metabolic Cytokines at Fasting and During Macronutrient Challenges: Influence of Obesity, Female Androgen Excess and Sex. Nutrients. 2019; 11(11):2566. https://doi.org/10.3390/nu11112566
Chicago/Turabian StyleMartínez-García, M. Ángeles, Samuel Moncayo, María Insenser, Francisco Álvarez-Blasco, Manuel Luque-Ramírez, and Héctor F. Escobar-Morreale. 2019. "Metabolic Cytokines at Fasting and During Macronutrient Challenges: Influence of Obesity, Female Androgen Excess and Sex" Nutrients 11, no. 11: 2566. https://doi.org/10.3390/nu11112566