Lifelong n-3 Polyunsaturated Fatty Acid Exposure Modulates Size of Mammary Epithelial Cell Populations and Expression of Caveolae Resident Proteins in Fat-1 Mice



Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. Animals, Diets and Phenotyping
2.3. Euthanization and Tissue Collection
2.4. Epithelial Cell Characterization and Caveolae Isolation
2.5. Protein Expression
2.6. Phospholipid Fatty Acid Analysis
2.7. Statistical Analysis
3. Results
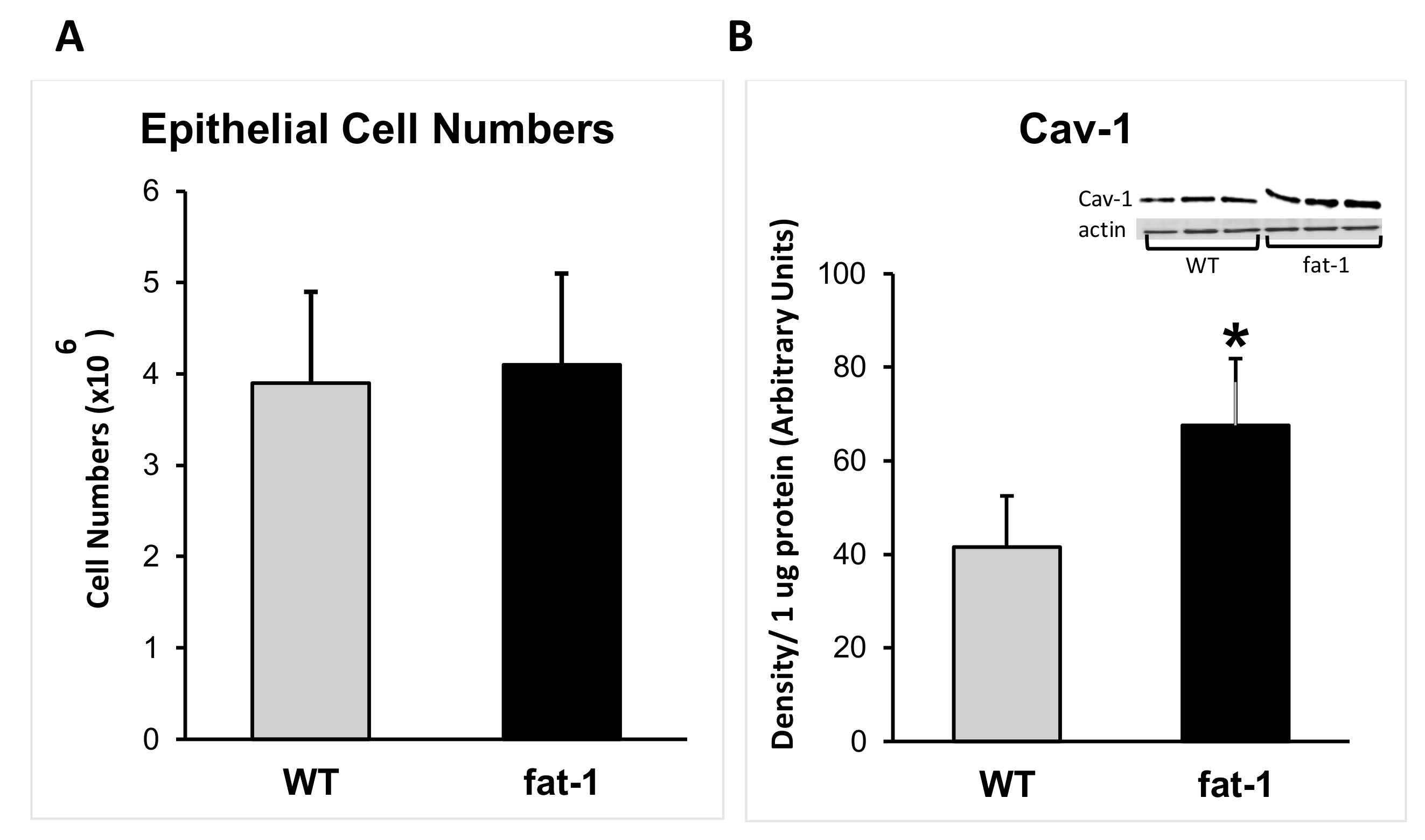
3.1. Epithelial Cell Populations
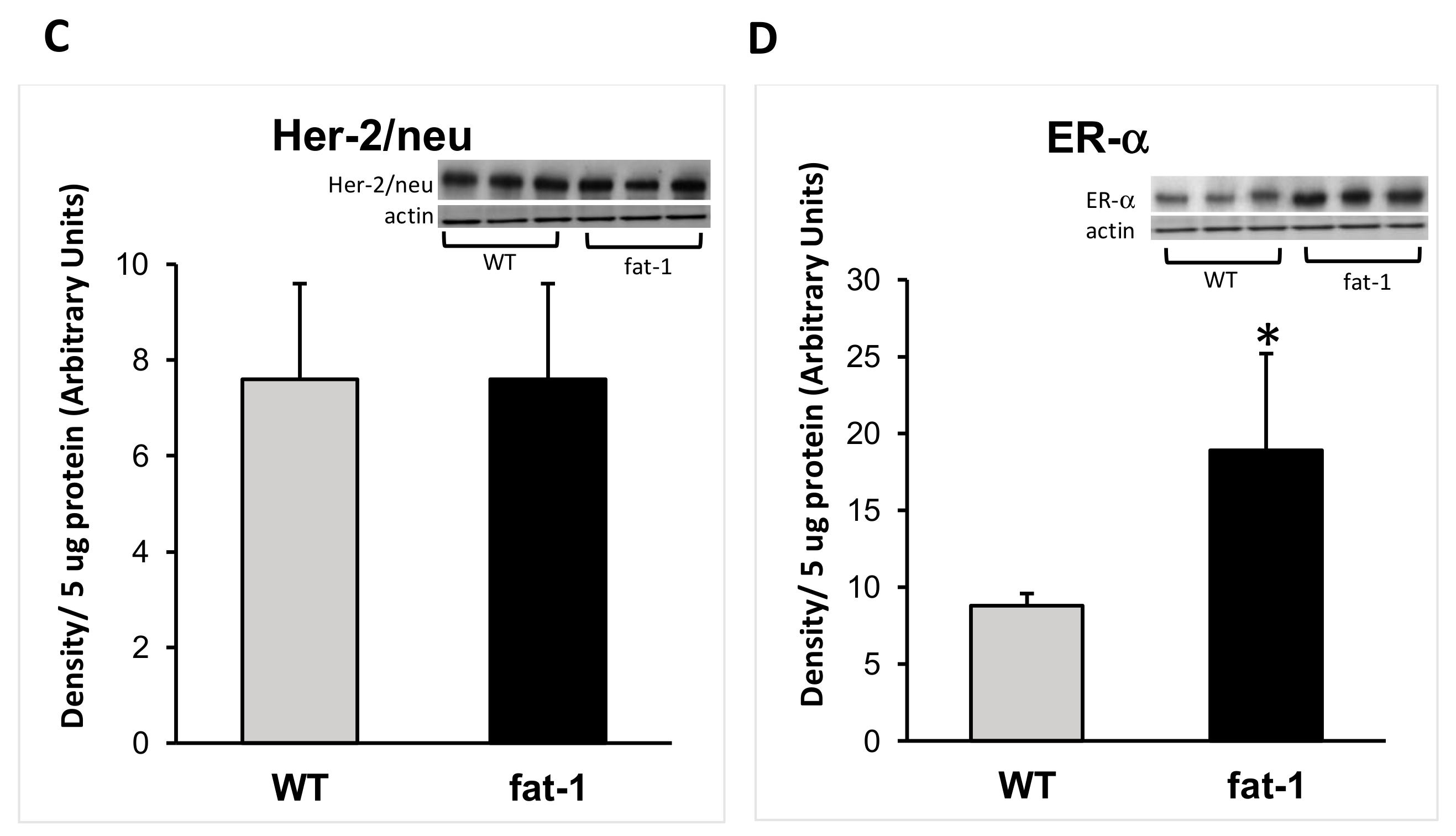
3.2. Protein Expression
3.3. Phospholipid Fatty Acids
4. Discussion
Author Contributions
Funding
Conflicts of Interest
References
- Parkin, D.M.; Bray, F.I.; Devesa, S.S. Cancer burden in the year 2000. The global picture. Eur. J. Cancer 2001, 37 (Suppl. 8), 4–66. [Google Scholar] [CrossRef]
- Howlader, N.; Noone, A.M.; Krapcho, M.; Miller, D.; Brest, A.; Yu, M.; Ruhl, J.; Tatalovich, Z.; Mariotto, A.; Lewis, D.R.; et al. SEER Cancer Statistics Review, 1975–2011; National Cancer Institute: Bethesda, MD, USA, 2014. Available online: https://seer.cancer.gov/archive/csr/1975_2011/ (accessed on 11 December 2018).
- Ferlay, J.; Shin, H.R.; Bray, F.; Forman, D.; Mathers, C.; Parkin, D.M. Estimates of worldwide burden of cancer in 2008: Globocan 2008. Int. J. Cancer 2010, 127, 2893–2917. [Google Scholar] [CrossRef] [PubMed]
- Saika, K.; Sobue, T. Cancer statisitcs in the world. Gan Kagaku Ryoho Cancer Chemother. 2013, 40, 2475–2480. [Google Scholar]
- Jemal, A.; Center, M.M.; DeSantis, C.; Ward, E.M. Global patterns of cancer incidence and mortality rates and trends. Cancer Epidemiol. Biomarkers Prev. 2010, 19, 1893–1907. [Google Scholar] [CrossRef] [PubMed]
- Kachuri, L.; De, P.; Ellison, L.F.; Semenciw, R.; Advisory Committee on Canadian Cancer Statistics. Cancer incidence, mortality and survival trends in Canada, 1970–2007. Chronic Dis. Inj. Can. 2013, 33, 69–80. [Google Scholar] [PubMed]
- Iwasaki, M.; Tsugane, S. Risk factors for breast cancer: Epidemiological evidence from Japanese studies. Cancer Sci. 2011, 102, 1607–1614. [Google Scholar] [CrossRef] [PubMed]
- MacLennan, M.B.; Clarke, S.E.; Perez, K.; Wood, G.A.; Muller, W.J.; Kang, J.X.; Ma, D.W. Mammary tumor development is directly inhibited by lifelong n-3 polyunsaturated fatty acids. J. Nutr. Biochem. 2013, 24, 388–395. [Google Scholar] [CrossRef]
- Leslie, M.A.; Abdelmagid, S.A.; Perez, K.; Muller, W.J.; Ma, D.W. Mammary tumour development is dose-dependently inhibited by n-3 polyunsaturated fatty acids in the MMTV-neu(ndl)-YD5 transgenic mouse model. Lipids Health Dis. 2014, 13, 96. [Google Scholar] [CrossRef]
- MacLennan, M.B.; Anderson, B.M.; Ma, D.W. Differential mammary gland development in FVB and C57Bl/6 mice: Implications for breast cancer research. Nutrients 2011, 3, 929–936. [Google Scholar] [CrossRef]
- Russo, J.; Tay, L.K.; Ciocca, D.R.; Russo, I.H. Molecular and cellular basis of the mammary gland susceptibility to carcinogenesis. Environ. Health Perspect. 1983, 49, 185–199. [Google Scholar] [CrossRef]
- Olivo, S.E.; Hilakivi-Clarke, L. Opposing effects of prepubertal low- and high-fat n-3 polyunsaturated fatty acid diets on rat mammary tumorigenesis. Carcinogenesis 2005, 26, 1563–1572. [Google Scholar] [CrossRef] [PubMed]
- Gudjonsson, T.; Adriance, M.C.; Sternlicht, M.D.; Petersen, O.W.; Bissell, M.J. Myoepithelial cells: Their origin and function in breast morphogenesis and neoplasia. J. Mammary Gland Biol. Neoplasia 2005, 10, 261–272. [Google Scholar] [CrossRef] [PubMed]
- Sternlicht, M.D.; Kedeshian, P.; Shao, Z.M.; Safarians, S.; Barsky, S.H. The human myoepithelial cell is a natural tumor suppressor. Clin. Cancer Res. 1997, 3, 1949–1958. [Google Scholar] [PubMed]
- Gusterson, B.A.; Warburton, M.J.; Mitchell, D.; Ellison, M.; Neville, A.M.; Rudland, P.S. Distribution of myoepithelial cells and basement membrane proteins in the normal breast and in benign and malignant breast diseases. Cancer Res. 1982, 42, 4763–4770. [Google Scholar] [PubMed]
- Gudjonsson, T.; Rønnov-Jessen, L.; Villadsen, R.; Rank, F.; Bissell, M.J.; Petersen, O.W. Normal and tumor-derived myoepithelial cells differ in their ability to interact with luminal breast epithelial cells for polarity and basement membrane deposition. J. Cell Sci. 2002, 115, 39–50. [Google Scholar]
- Rudland, P.S. Stem cells and the development of mammary cancers in experimental rats and in humans. Cancer Metastasis Rev. 1987, 6, 55–83. [Google Scholar] [CrossRef]
- Aoki, M.N.; Amarante, M.K.; Oda, J.M.; Watanabe, M.A. Caveolin involvement and modulation in breast cancer. Mini Rev. Med. Chem. 2011, 11, 1143–1152. [Google Scholar] [CrossRef]
- Hnasko, R.; Lisanti, M.P. The biology of caveolae: Lessons from caveolin knockout mice and implications for human disease. Mol. Interv. 2003, 3, 445–464. [Google Scholar] [CrossRef]
- Sotgia, F.; Martinez-Outschoorn, U.E.; Howell, A.; Pestell, R.G.; Pavlides, S.; Lisanti, M.P. Caveolin-1 and cancer metabolism in the tumor microenvironment: Markers, models, and mechanisms. Annu. Rev. Pathol. 2012, 7, 423–467. [Google Scholar] [CrossRef]
- Patani, N.; Martin, L.A.; Reis-Filho, J.S.; Dowsett, M. The role of caveolin-1 in human breast cancer. Breast Cancer Res. Treat. 2012, 131, 1–15. [Google Scholar] [CrossRef]
- Giacinti, L.; Claudio, P.P.; Lopez, M.; Giordano, A. Epigenetic information and estrogen receptor alpha expression in breast cancer. Oncologist 2006, 11, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Molina, R.; Escudero, J.M.; Muñoz, M.; Augé, J.M.; Filella, X. Circulating levels of Her-2/neu oncoprotein in breast cancer. Clin. Chem. Lab. Med. 2012, 50, 5–21. [Google Scholar] [CrossRef] [PubMed]
- Kang, J.X.; Wang, J.; Wu, L.; Kang, Z.B. Transgenic mice: Fat-1 mice convert n-6 to n-3 fatty acids. Nature 2004, 427, 504. [Google Scholar] [CrossRef] [PubMed]
- Marcondes, F.K.; Bianchi, F.J.; Tanno, A.P. Determination of the estrous cycle phases of rats: Some helpful considerations. Braz. J. Biol. 2002, 62, 609–614. [Google Scholar] [CrossRef]
- Prater, M.; Shehata, M.; Watson, C.J.; Stingl, J. Enzymatic dissociation, flow cytometric analysis, and culture of normal mouse mammary tissue. Methods Mol. Biol. 2013, 946, 395–409. [Google Scholar]
- Macdonald, J.L.; Pike, L.J. A simplified method for the preparation of detergent-free lipid rafts. J. Lipid Res. 2005, 46, 1061–1067. [Google Scholar] [CrossRef]
- Folch, J.L.M. A simple method for the isolation and purification of total lipids from animal tissues. J. Biol. Chem. 1957, 226, 497–509. [Google Scholar]
- Touchstone, J.C.; Chen, J.C.; Beaver, K.M. Improved separation of phospholipids in thin layer chromatography. Lipids 1980, 15, 61–62. [Google Scholar] [CrossRef]
- Razandi, M.; Oh, P.; Pedram, A.; Schnitzer, J.; Levin, E.R. ERs associate with and regulate the production of caveolin: Implications for signaling and cellular actions. Mol. Endocrinol. 2002, 16, 100–115. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | g/Kg |
|---|---|
| Casein | 200 |
| L-cystine | 3.0 |
| Cornstarch | 336.7 |
| Maltodextrin | 132 |
| Sucrose | 100 |
| Cellulose | 50 |
| Safflower oil | 97 |
| t-butylhydroquinone | 0.019 |
| Mineral mix S10022G | 35 |
| Vitamin Mix V10037 | 10 |
| Choline bitartrate | 2.5 |
| PC | PE | |||||
|---|---|---|---|---|---|---|
| WT | Fat-1 | p Value | WT | Fat-1 | p Value | |
| 18:0, stearic | 44.1 * | 34.6 | ≤0.05 | 50.9 | 47.1 | 0.116 |
| 18:1, oleic | 19.4 | 16.7 | 0.371 | 29.2 | 32.7 | 0.214 |
| 18:2n6, linoleic | 24.0 * | 15.2 | ≤0.05 | 11.3 * | 4.7 | ≤0.05 |
| 18:3n3, alpha linolenic | 0.2 | 3.9 * | ≤0.05 | 0 | 1.4 * | ≤0.05 |
| 20:4n6, arachidonic | 12.3 * | 8.6 | ≤0.05 | 8.6 * | 2.6 | ≤0.05 |
| 20:5n3, eicosapentaenoic | 0 | 8.8 * | ≤0.05 | 0 | 5.6 * | ≤0.05 |
| 22:6n3, docosahexaenoic | 0 | 12.1 * | ≤0.05 | 0 | 5.9 * | ≤0.05 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hillyer, L.M.; Kang, J.X.; Ma, D.W.L. Lifelong n-3 Polyunsaturated Fatty Acid Exposure Modulates Size of Mammary Epithelial Cell Populations and Expression of Caveolae Resident Proteins in Fat-1 Mice. Nutrients 2019, 11, 2477. https://doi.org/10.3390/nu11102477
Hillyer LM, Kang JX, Ma DWL. Lifelong n-3 Polyunsaturated Fatty Acid Exposure Modulates Size of Mammary Epithelial Cell Populations and Expression of Caveolae Resident Proteins in Fat-1 Mice. Nutrients. 2019; 11(10):2477. https://doi.org/10.3390/nu11102477
Chicago/Turabian StyleHillyer, Lyn M., Jing X. Kang, and David W.L. Ma. 2019. "Lifelong n-3 Polyunsaturated Fatty Acid Exposure Modulates Size of Mammary Epithelial Cell Populations and Expression of Caveolae Resident Proteins in Fat-1 Mice" Nutrients 11, no. 10: 2477. https://doi.org/10.3390/nu11102477
APA StyleHillyer, L. M., Kang, J. X., & Ma, D. W. L. (2019). Lifelong n-3 Polyunsaturated Fatty Acid Exposure Modulates Size of Mammary Epithelial Cell Populations and Expression of Caveolae Resident Proteins in Fat-1 Mice. Nutrients, 11(10), 2477. https://doi.org/10.3390/nu11102477