Single-Dose SDA-Rich Echium Oil Increases Plasma EPA, DPAn3, and DHA Concentrations


 ,
,
Abstract
1. Introduction
2. Materials and Methods
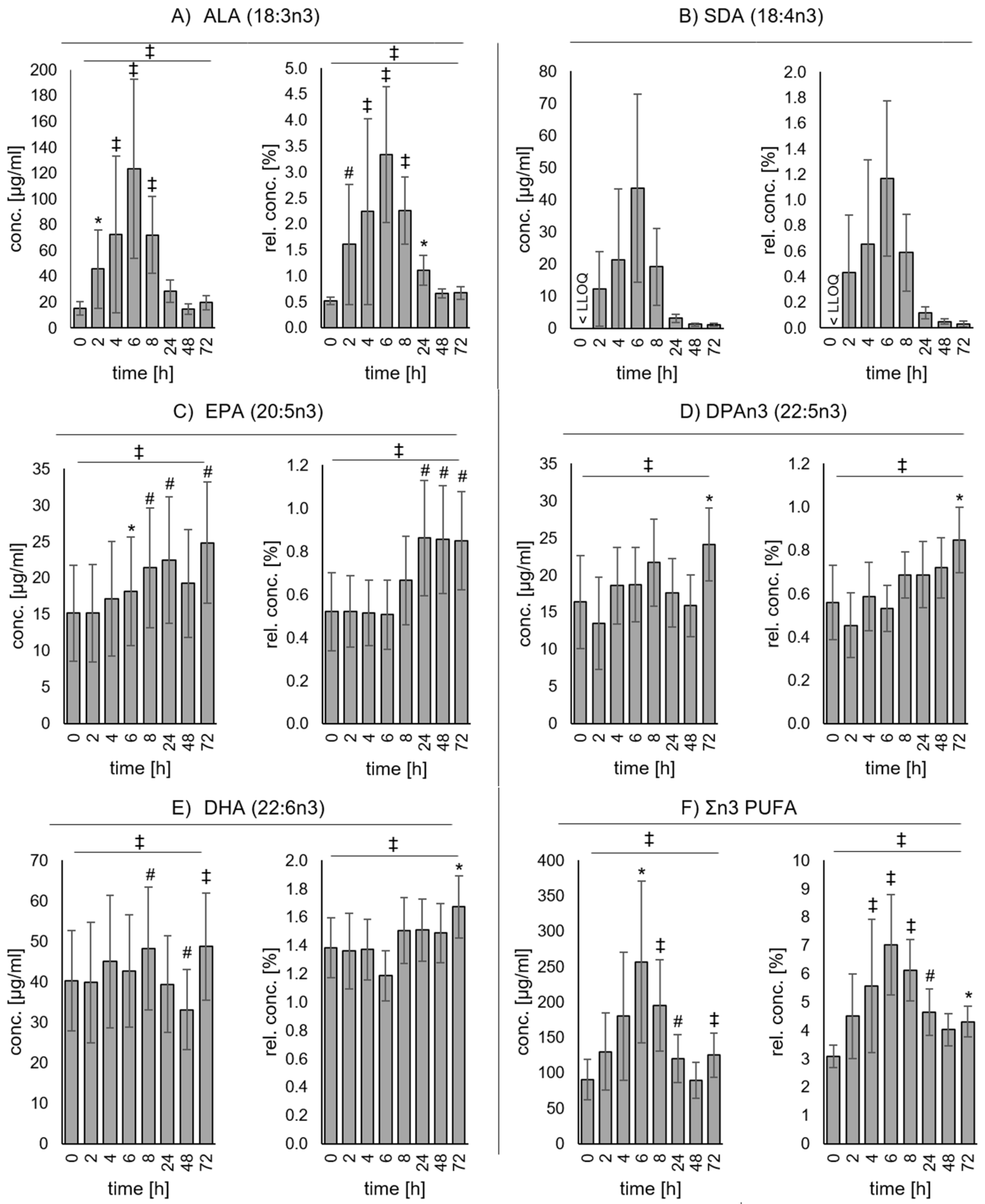
3. Results
3.1. Study Population
3.2. Changes of Plasma Fatty Acid Concentrations
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALA | alpha-linolenic acid (C18:3n3) |
| ALT | Alanine Aminotransferase |
| AST | Aspartate Aminotransferase |
| BMI | body mass index |
| DHA | docosahexaenoic acid (C22:6n3) |
| dias BP | diastolic blood pressure |
| DPAn3 | docosapentaenoic acid (C22:5n3) |
| D6D | delta-6-desaturase |
| EO | echium oil |
| EPA | eicosapentaenoic acid (C20:5n3) |
| FADS2 | fatty acid desaturase 2 |
| FAME | fatty acid methyl esters |
| FID | flame ionization detection |
| GC | gas chromatography |
| GGT | Gamma-glutamyl transpeptidase |
| HDL | high density lipoprotein |
| LA | linoleic acid (C18:2n6) |
| LDL | low density lipoprotein |
| LLOQ | lower limit of quantification |
| MUFA | monounsaturated fatty acids |
| n3 | omega-3 |
| n6 | omega-6 |
| PUFA | polyunsaturated fatty acids |
| SD | standard deviation |
| SDA | stearidonic acid (C18:4n3) |
| SFA | saturated fatty acids |
| sys BP | systolic blood pressure |
| TC | total cholesterol |
| TG | triglycerides |
References
- Calder, P.C. n-3 Fatty acids and cardiovascular disease: Evidence explained and mechanisms explored. Clin. Sci. 2004, 107, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Calder, P.C. Very long chain omega-3 (n-3) fatty acids and human health: Omega-3 fatty acids and health. Eur. J. Lipid Sci. Technol. 2014, 116, 1280–1300. [Google Scholar] [CrossRef]
- Harris, W.S. The omega-3 index: From biomarker to risk marker to risk factor. Curr. Atheroscler. Rep. 2009, 11, 411–417. [Google Scholar] [CrossRef] [PubMed]
- Marik, P.E.; Varon, J. Omega-3 Dietary Supplements and the Risk of Cardiovascular Events: A Systematic Review. Clin. Cardiol. 2009, 32, 365–372. [Google Scholar] [CrossRef] [PubMed]
- Mozaffarian, D.; Wu, J.H.Y. Omega-3 Fatty Acids and Cardiovascular Disease: Effects on risk factors, molecular pathways, and clinical events. J. Am. Coll. Cardiol. 2011, 58, 2047–2067. [Google Scholar] [CrossRef]
- Calder, P.C. Marine omega-3 fatty acids and inflammatory processes: Effects, mechanisms and clinical relevance. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2015, 1851, 469–484. [Google Scholar] [CrossRef]
- Zhang, G.; Panigrahy, D.; Mahakian, L.M.; Yang, J.; Liu, J.-Y.; Stephen Lee, K.S.; Wettersten, H.I.; Ulu, A.; Hu, X.; Tam, S.; et al. Epoxy metabolites of docosahexaenoic acid (DHA) inhibit angiogenesis, tumor growth, and metastasis. Proc. Natl. Acad. Sci. USA 2013, 110, 6530–6535. [Google Scholar] [CrossRef]
- SanGiovanni, J.P.; Parra-Cabrera, S.; Colditz, G.A.; Berkey, C.S.; Dwyer, J.T. Meta-analysis of dietary essential fatty acids and long-chain polyunsaturated fatty acids as they relate to visual resolution acuity in healthy preterm infants. Pediatrics 2000, 105, 1292–1298. [Google Scholar] [CrossRef]
- Kaur, G.; Cameron-Smith, D.; Garg, M.; Sinclair, A.J. Docosapentaenoic acid (22:5n-3): A review of its biological effects. Prog. Lipid Res. 2011, 50, 28–34. [Google Scholar] [CrossRef]
- Kaur, G.; Guo, X.-F.; Sinclair, A.J. Short update on docosapentaenoic acid: A bioactive long-chain n-3 fatty acid. Curr. Opin. Clin. Nutr. Metab. Care 2016, 19, 88–91. [Google Scholar] [CrossRef]
- Drouin, G.; Rioux, V.; Legrand, P. The n-3 docosapentaenoic acid (DPA): A new player in the n-3 long chain polyunsaturated fatty acid family. Biochimie 2019, 159, 36–48. [Google Scholar] [CrossRef] [PubMed]
- Micha, R.; Khatibzadeh, S.; Shi, P.; Fahimi, S.; Lim, S.; Andrews, K.G.; Engell, R.E.; Powles, J.; Ezzati, M.; Mozaffarian, D.; et al. Global, regional, and national consumption levels of dietary fats and oils in 1990 and 2010: A systematic analysis including 266 country-specific nutrition surveys. BMJ 2014, 348, g2272. [Google Scholar] [CrossRef] [PubMed]
- Stark, K.D.; Van Elswyk, M.E.; Higgins, M.R.; Weatherford, C.A.; Salem, N. Global survey of the omega-3 fatty acids, docosahexaenoic acid and eicosapentaenoic acid in the blood stream of healthy adults. Prog. Lipid Res. 2016, 63, 132–152. [Google Scholar] [CrossRef] [PubMed]
- Lands, B. Consequences of essential fatty acids. Nutrients 2012, 4, 1338–1357. [Google Scholar] [CrossRef] [PubMed]
- Wood, K.E.; Mantzioris, E.; Gibson, R.A.; Ramsden, C.E.; Muhlhausler, B.S. The effect of modifying dietary LA and ALA intakes on omega-3 long chain polyunsaturated fatty acid (n-3 LCPUFA) status in human adults: A systematic review and commentary. Prostaglandins Leukot. Essent. Fatty Acids (PLEFA) 2015, 95, 47–55. [Google Scholar] [CrossRef] [PubMed]
- Arterburn, L.M.; Hall, E.B.; Oken, H. Distribution, interconversion, and dose response of n- 3 fatty acids in humans. Am. J. Clin. Nutr. 2006, 83, S1467–S1476. [Google Scholar] [CrossRef]
- Greupner, T.; Kutzner, L.; Nolte, F.; Strangmann, A.; Kohrs, H.; Hahn, A.; Schebb, N.H.; Schuchardt, J.P. Effects of a 12-week high-α-linolenic acid intervention on EPA and DHA concentrations in red blood cells and plasma oxylipin pattern in subjects with a low EPA and DHA status. Food Funct. 2018, 9, 1587–1600. [Google Scholar] [CrossRef]
- Greupner, T.; Kutzner, L.; Pagenkopf, S.; Kohrs, H.; Hahn, A.; Schebb, N.H.; Schuchardt, J.P. Effects of a low and a high dietary LA/ALA ratio on long-chain PUFA concentrations in red blood cells. Food Funct. 2018, 9, 4742–4754. [Google Scholar] [CrossRef]
- Bernert, J.T.; Sprecher, H. Studies to determine the role rates of chain elongation and desaturation play in regulating the unsaturated fatty acid composition of rat liver lipids. Biochim. Biophys. Acta (BBA)-Lipids Lipid Metab. 1975, 398, 354–363. [Google Scholar] [CrossRef]
- Sprecher, H.; Chen, Q.; Yin, F.Q. Regulation of the biosynthesis of 22:5n-6 and 22:6n-3: A complex intracellular process. Lipids 1999, 34, S153–S156. [Google Scholar] [CrossRef]
- Whelan, J.; Gouffon, J.; Zhao, Y. Effects of Dietary Stearidonic Acid on Biomarkers of Lipid Metabolism. J. Nutr. 2012, 142, 630S–634S. [Google Scholar] [CrossRef] [PubMed]
- James, M.J.; Ursin, V.M.; Cleland, L.G. Metabolism of stearidonic acid in human subjects: Comparison with the metabolism of other n-3 fatty acids. Am. J. Clin. Nutr. 2003, 77, 1140–1145. [Google Scholar] [CrossRef] [PubMed]
- Miles, E.A.; Banerjee, T.; Dooper, M.M.B.W.; M’Rabet, L.; Graus, Y.M.F.; Calder, P.C. The influence of different combinations of γ-linolenic acid, stearidonic acid and EPA on immune function in healthy young male subjects. Br. J. Nutr. 2004, 91, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Lemke, S.L.; Vicini, J.L.; Su, H.; Goldstein, D.A.; Nemeth, M.A.; Krul, E.S.; Harris, W.S. Dietary intake of stearidonic acid–enriched soybean oil increases the omega-3 index: Randomized, double-blind clinical study of efficacy and safety. Am. J. Clin. Nutr. 2010, 92, 766–775. [Google Scholar] [CrossRef] [PubMed]
- Krul, E.S.; Lemke, S.L.; Mukherjea, R.; Taylor, M.L.; Goldstein, D.A.; Su, H.; Liu, P.; Lawless, A.; Harris, W.S.; Maki, K.C. Effects of duration of treatment and dosage of eicosapentaenoic acid and stearidonic acid on red blood cell eicosapentaenoic acid content. Prostaglandins Leukot. Essent. Fat. Acids 2012, 86, 51–59. [Google Scholar] [CrossRef] [PubMed]
- Baker, E.J.; Miles, E.A.; Burdge, G.C.; Yaqoob, P.; Calder, P.C. Metabolism and functional effects of plant-derived omega-3 fatty acids in humans. Prog. Lipid Res. 2016, 64, 30–56. [Google Scholar] [CrossRef] [PubMed]
- Ostermann, A.I.; Müller, M.; Willenberg, I.; Schebb, N.H. Determining the fatty acid composition in plasma and tissues as fatty acid methyl esters using gas chromatography–A comparison of different derivatization and extraction procedures. Prostaglandins Leukot. Essent. Fat. Acids (PLEFA) 2014, 91, 235–241. [Google Scholar] [CrossRef]
- Harris, W.S.; Thomas, R.M. Biological variability of blood omega-3 biomarkers. Clin. Biochem. 2010, 43, 338–340. [Google Scholar] [CrossRef]
- Patenaude, A.; Rodriguez-Leyva, D.; Edel, A.L.; Dibrov, E.; Dupasquier, C.M.C.; Austria, J.A.; Richard, M.N.; Chahine, M.N.; Malcolmson, L.J.; Pierce, G.N. Bioavailability of α-linolenic acid from flaxseed diets as a function of the age of the subject. Eur. J. Clin. Nutr. 2009, 63, 1123–1129. [Google Scholar] [CrossRef]
- Abdelmagid, S.A.; Clarke, S.E.; Nielsen, D.E.; Badawi, A.; El-Sohemy, A.; Mutch, D.M.; Ma, D.W.L. Comprehensive Profiling of Plasma Fatty Acid Concentrations in Young Healthy Canadian Adults. PLoS ONE 2015, 10, e0116195. [Google Scholar] [CrossRef]
- Flock, M.R.; Skulas-Ray, A.C.; Harris, W.S.; Etherton, T.D.; Fleming, J.A.; Kris-Etherton, P.M. Determinants of Erythrocyte Omega-3 Fatty Acid Content in Response to Fish Oil Supplementation: A Dose-Response Randomized Controlled Trial. J. Am. Heart Assoc. 2013, 2, e000513. [Google Scholar] [CrossRef] [PubMed]
- Gellert, S.; Schuchardt, J.P.; Hahn, A. Low long chain omega-3 fatty acid status in middle-aged women. Prostaglandin Leukot. Essent. Fat. Acids 2017, 117, 54–59. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Pottala, J.V.; Lacey, S.M.; Vasan, R.S.; Larson, M.G.; Robins, S.J. Clinical correlates and heritability of erythrocyte eicosapentaenoic and docosahexaenoic acid content in the Framingham Heart Study. Atherosclerosis 2012, 225, 425–431. [Google Scholar] [CrossRef] [PubMed]
- Plourde, M.; Chouinard-Watkins, R.; Vandal, M.; Zhang, Y.; Lawrence, P.; Brenna, J.T.; Cunnane, S.C. Plasma incorporation, apparent retroconversion and β-oxidation of 13C-docosahexaenoic acid in the elderly. Nutr. Metab. 2011, 8, 5. [Google Scholar] [CrossRef] [PubMed]
- Bakewell, L.; Burdge, G.C.; Calder, P.C. Polyunsaturated fatty acid concentrations in young men and women consuming their habitual diets. Br. J. Nutr. 2006, 96, 93. [Google Scholar] [CrossRef] [PubMed]
- Crowe, F.L.; Murray Skeaff, C.; Green, T.J.; Gray, A.R. Serum n-3 long-chain PUFA differ by sex and age in a population-based survey of New Zealand adolescents and adults. Br. J. Nutr. 2008, 99, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Metherel, A.H.; Armstrong, J.M.; Patterson, A.C.; Stark, K.D. Assessment of blood measures of n-3 polyunsaturated fatty acids with acute fish oil supplementation and washout in men and women. Prostaglandins Leukot. Essent. Fat. Acids 2009, 81, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Langlois, K.; Ratnayake, W.M.N. Omega-3 Index of Canadian adults. Health Rep. 2015, 26, 11. [Google Scholar]
- Sands, S.A.; Reid, K.J.; Windsor, S.L.; Harris, W.S. The impact of age, body mass index, and fish intake on the EPA and DHA content of human erythrocytes. Lipids 2005, 40, 343. [Google Scholar] [CrossRef]
- Sala-Vila, A.; Harris, W.S.; Cofán, M.; Pérez-Heras, A.M.; Pintó, X.; Lamuela-Raventós, R.M.; Covas, M.-I.; Estruch, R.; Ros, E. Determinants of the omega-3 index in a Mediterranean population at increased risk for CHD. Br. J. Nutr. 2011, 106, 425–431. [Google Scholar] [CrossRef]
- Surette, M.E.; Edens, M.; Chilton, F.H.; Tramposch, K.M. Dietary Echium oil increases plasma and neutrophil long-chain (n-3) fatty acids and lowers serum triacylglycerols in hypertriglyceridemic humans. J. Nutr. 2004, 134, 1406–1411. [Google Scholar] [CrossRef] [PubMed]
- Harris, W.S.; Lemke, S.L.; Hansen, S.N.; Goldstein, D.A.; DiRienzo, M.A.; Su, H.; Nemeth, M.A.; Taylor, M.L.; Ahmed, G.; George, C. Stearidonic Acid-Enriched Soybean Oil Increased the Omega-3 Index, an Emerging Cardiovascular Risk Marker. Lipids 2008, 43, 805–811. [Google Scholar] [CrossRef] [PubMed]
- Lemke, S.L.; Maki, K.C.; Hughes, G.; Taylor, M.L.; Krul, E.S.; Goldstein, D.A.; Su, H.; Rains, T.M.; Mukherjea, R. Consumption of Stearidonic Acid−Rich Oil in Foods Increases Red Blood Cell Eicosapentaenoic Acid. J. Acad. Nutr. Diet. 2013, 113, 1044–1056. [Google Scholar] [CrossRef] [PubMed]
- Arm, J.P.; Boyce, J.A.; Wang, L.; Chhay, H.; Zahid, M.; Patil, V.; Govindarajulu, U.; Ivester, P.; Weaver, K.L.; Sergeant, S.; et al. Impact of botanical oils on polyunsaturated fatty acid metabolism and leukotriene generation in mild asthmatics. Lipids Health Dis. 2013, 12, 141. [Google Scholar] [CrossRef] [PubMed]
- Kuhnt, K.; Fuhrmann, C.; Kohler, M.; Kiehnmiddlef, M.; Jahreis, G. Dietary Echium Oil Increases Long-Chain n-3 PUFAs, Including Docosapentaenoic Acid, in Blood Fractions and Alters Biochemical Markers for Cardiovascular Disease Independently of Age, Sex, and Metabolic Syndrome. J. Nutr. 2014, 144, 447–460. [Google Scholar] [CrossRef] [PubMed]
- Pieters, D.J.M.; Mensink, R.P. Effects of stearidonic acid on serum triacylglycerol concentrations in overweight and obese subjects: A randomized controlled trial. Eur. J. Clin. Nutr. 2015, 69, 121–126. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, M.; Jahreis, G.; Bothor, K.; Drechsel, C.; Kiehnmiddlef, M.; Blüher, M.; Dawczynski, C. Benefits of foods supplemented with vegetable oils rich in α-linolenic, stearidonic or docosahexaenoic acid in hypertriglyceridemic subjects: A double-blind, randomized, controlled trail. Eur. J. Nutr. 2015, 54, 881–893. [Google Scholar] [CrossRef]
- Kuhnt, K.; Weiß, S.; Kiehnmiddlef, M.; Jahreis, G. Consumption of echium oil increases EPA and DPA in blood fractions more efficiently compared to linseed oil in humans. Lipids Health Dis. 2016, 15, 32. [Google Scholar] [CrossRef]
- Cho, H.P.; Nakamura, M.T.; Clarke, S.D. Cloning, Expression, and Nutritional Regulation of the Mammalian Δ-6 Desaturase. J. Biol. Chem. 1999, 274, 471–477. [Google Scholar] [CrossRef]
- Simopoulos, A.P. Genetic variants in the metabolism of omega-6 and omega-3 fatty acids: Their role in the determination of nutritional requirements and chronic disease risk. Exp. Biol. Med. 2010, 235, 785–795. [Google Scholar] [CrossRef]
- Sprecher, H. Metabolism of highly unsaturated n-3 and n-6 fatty acids. Biochim. Biophys. Acta (BBA)-Mol. Cell Biol. Lipids 2000, 1486, 219–231. [Google Scholar] [CrossRef]
- Portolesi, R.; Powell, B.C.; Gibson, R.A. Competition between 24:5n-3 and ALA for 6 desaturase may limit the accumulation of DHA in HepG2 cell membranes. J. Lipid Res. 2007, 48, 1592–1598. [Google Scholar] [CrossRef] [PubMed]
- Miller, E.; Kaur, G.; Larsen, A.; Loh, S.P.; Linderborg, K.; Weisinger, H.S.; Turchini, G.M.; Cameron-Smith, D.; Sinclair, A.J. A short-term n-3 DPA supplementation study in humans. Eur. J. Nutr. 2013, 52, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Williams, C.M.; Burdge, G. Long-chain n-3 PUFA: Plant v. marine sources. Proc. Nutr. Soc. 2006, 65, 42–50. [Google Scholar] [CrossRef] [PubMed]
- Childs, C.E.; Romeu-Nadal, M.; Burdge, G.C.; Calder, P.C. Gender differences in the n-3 fatty acid content of tissues. Proc. Nutr. Soc. 2008, 67, 19–27. [Google Scholar] [CrossRef]


{kind=link}
| Fatty Acid | Common Name | Concentration (g/100 g) |
|---|---|---|
| 16:0 | Palmitic acid | 6.95 |
| 16:1n7 | Palmitoleic acid | 0.0925 |
| 18:0 | Stearic acid | 3.62 |
| 18:1n9 | Oleic acid | 15.5 |
| 18:1n7 | Vaccenic acid | 0.910 |
| 18:2n6 | Linoleic acid | 15.2 |
| 18:3n6 | γ-Linolenic acid | 10.4 |
| 18:3n3 | α-Linolenic acid | 30.2 |
| 18:4n3 | Stearidonic acid | 11.8 |
| 20:0 | Arachidic acid | 0.0980 |
| 20:1n9 | Gondoic acid | 0.592 |
| 22:0 | Behenic acid | 0.117 |
| 22:1n9 | Erucic acid | 0.129 |
| SFA | 10.8 | |
| MUFA | 17.3 | |
| PUFA | 67.6 | |
| Σn3 PUFA | 42.0 | |
| Σn6 PUFA | 25.6 |
| Parameter | mean | ± | SD |
|---|---|---|---|
| Age (years) | 24.6 | ± | 2.43 |
| Weight (kg) | 84.5 | ± | 8.79 |
| BMI (kg/m2) | 24.6 | ± | 2.06 |
| Sys BP (mmHg) | 123 | ± | 13.9 |
| Dias BP (mmHg) | 74.2 | ± | 5.57 |
| AST (U/l) | 25.4 | ± | 5.28 |
| ALT (U/l) | 23.0 | ± | 13.5 |
| GGT (U/l) | 20.3 | ± | 8.54 |
| TC (mg/dl) | 166 | ± | 38.4 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Greupner, T.; Koch, E.; Kutzner, L.; Hahn, A.; Schebb, N.H.; Schuchardt, J.P. Single-Dose SDA-Rich Echium Oil Increases Plasma EPA, DPAn3, and DHA Concentrations. Nutrients 2019, 11, 2346. https://doi.org/10.3390/nu11102346
Greupner T, Koch E, Kutzner L, Hahn A, Schebb NH, Schuchardt JP. Single-Dose SDA-Rich Echium Oil Increases Plasma EPA, DPAn3, and DHA Concentrations. Nutrients. 2019; 11(10):2346. https://doi.org/10.3390/nu11102346
Chicago/Turabian StyleGreupner, Theresa, Elisabeth Koch, Laura Kutzner, Andreas Hahn, Nils Helge Schebb, and Jan Philipp Schuchardt. 2019. "Single-Dose SDA-Rich Echium Oil Increases Plasma EPA, DPAn3, and DHA Concentrations" Nutrients 11, no. 10: 2346. https://doi.org/10.3390/nu11102346
APA StyleGreupner, T., Koch, E., Kutzner, L., Hahn, A., Schebb, N. H., & Schuchardt, J. P. (2019). Single-Dose SDA-Rich Echium Oil Increases Plasma EPA, DPAn3, and DHA Concentrations. Nutrients, 11(10), 2346. https://doi.org/10.3390/nu11102346