Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans


Abstract
1. Introduction
2. Food Sources and Dietary Intakes
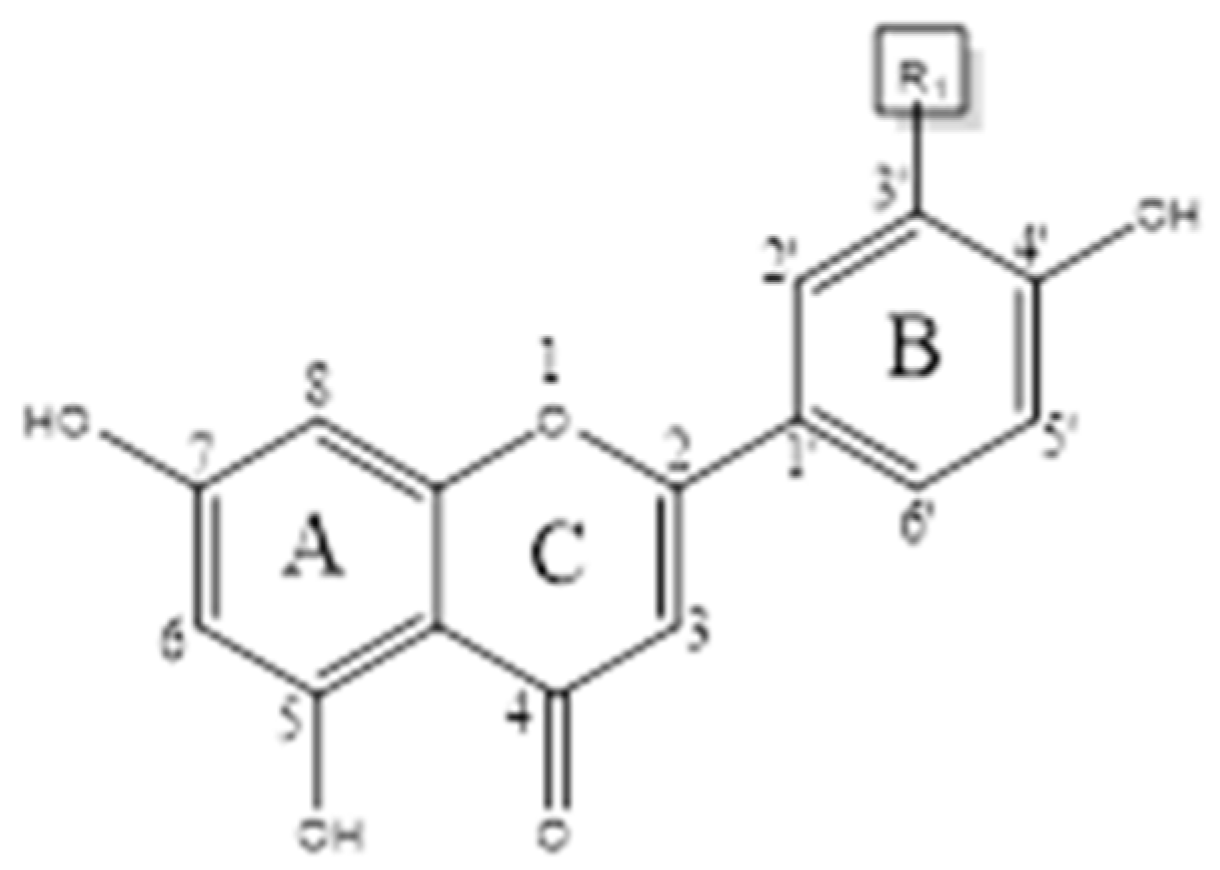
3. Chemical Structure
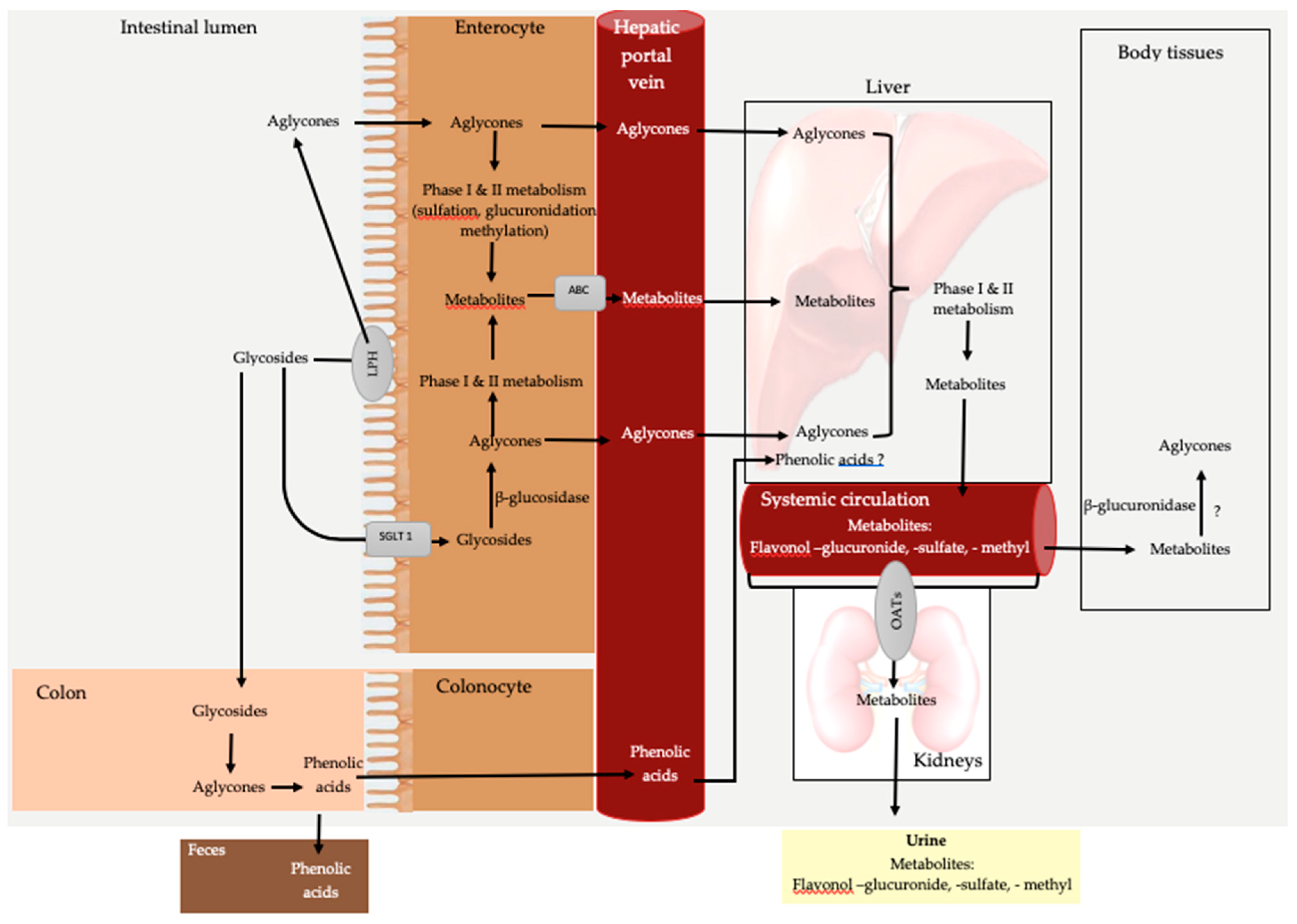
4. Bioavailability
4.1. Quercetin Bioavailability
4.1.1. Digestion and Absorption
4.1.2. Metabolism and Excretion
4.2. Kaempferol Bioavailability
4.2.1. Digestion and Absorption
4.2.2. Metabolism and Excretion
5. Potential Bioactivity in Humans
5.1. Quercetin Bioactivity
5.2. Kaempferol Bioactivity
6. Safety
6.1. Quercetin Safety
6.2. Kaempferol Safety
7. Conclusions
Funding
Conflicts of Interest
References
- Centers for Disease Control and Prevention. Heart Disease Facts. Available online: https://www.cdc.gov/heartdisease/facts.htm (accessed on 24 September 2019).
- Aune, D.; Giovannucci, E.; Boffetta, P.; Fadnes, L.T.; Keum, N.; Norat, T.; Greenwood, D.C.; Riboli, E.; Vatten, L.J.; Tonstad, S. Fruit and vegetable intake and the risk of cardiovascular disease, total cancer and all-cause mortality—A systematic review and dose-response meta-analysis of prospective studies. Int. J. Epidemiol. 2017, 46, 1029–1056. [Google Scholar] [CrossRef] [PubMed]
- McCullough, M.L.; Peterson, J.J.; Patel, R.; Jacques, P.F.; Shah, R.; Dwyer, J.T. Flavonoid intake and cardiovascular disease mortality in a prospective cohort of US adults1234. Am. J. Clin. Nutr. 2012, 95, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Erdman, J.W.; Balentine, D.; Arab, L.; Beecher, G.; Dwyer, J.T.; Folts, J.; Harnly, J.; Hollman, P.; Keen, C.L.; Mazza, G.; et al. Flavonoids and Heart Health: Proceedings of the ILSI North America Flavonoids Workshop, 31 May–1 June 2005, Washington, DC. J. Nutr. 2007, 137, 718S–737S. [Google Scholar] [CrossRef] [PubMed]
- Adebamowo, C.A.; Cho, E.; Sampson, L.; Katan, M.B.; Spiegelman, D.; Willett, W.C.; Holmes, M.D. Dietary flavonols and flavonol-rich foods intake and the risk of breast cancer. Int. J. Cancer 2005, 114, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Crespo, I.; García-Mediavilla, M.V.; Gutiérrez, B.; Sánchez-Campos, S.; Tuñón, M.J.; González-Gallego, J. A comparison of the effects of kaempferol and quercetin on cytokine-induced pro-inflammatory status of cultured human endothelial cells. Br. J. Nutr. 2008, 100, 968–976. [Google Scholar] [CrossRef] [PubMed]
- Espley, R.V.; Butts, C.A.; Laing, W.A.; Martell, S.; Smith, H.; McGhie, T.K.; Zhang, J.; Paturi, G.; Hedderley, D.; Bovy, A.; et al. Dietary flavonoids from modified apple reduce inflammation markers and modulate gut microbiota in mice. J. Nutr. 2014, 144, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Badary, O.A.; Yassin, N.A.Z.; El-Shenawy, S.M.A.; EL-Moneem, M.A.; AL-Shafeiy, H.M. Study of the effect of Allium porrum on hypertension induced in rats. Available online: https://pdfs.semanticscholar.org/c5a4/82740ec2a0bbc33acf5a64996475be279203.pdf (accessed on 20 November 2018).
- Xiao, J. Dietary flavonoid aglycones and their glycosides: Which show better biological significance? Crit. Rev. Food Sci. Nutr. 2017, 57, 1874–1905. [Google Scholar] [CrossRef]
- Rein, M.J.; Renouf, M.; Cruz-Hernandez, C.; Actis-Goretta, L.; Thakkar, S.K.; da Silva Pinto, M. Bioavailability of bioactive food compounds: A challenging journey to bioefficacy. Br. J. Clin. Pharm. 2013, 75, 588–602. [Google Scholar] [CrossRef]
- Hollman, P.C.; Vries, D.H.J.; Leeuwen, V.D.S.; Mengelers, M.J.; Katan, M.B. Absorption of dietary quercetin glycosides and quercetin in healthy ileostomy volunteers. Am. J. Clin. Nutr. 1995, 62, 1276–1282. [Google Scholar] [CrossRef]
- Hollman, P.C.H.; Gaag, M.V.D.; Mengelers, M.J.B.; Van Trijp, J.M.P.; De Vries, J.H.M.; Katan, M.B. Absorption and disposition kinetics of the dietary antioxidant quercetin in man. Free Radic. Biol. Med. 1996, 21, 703–707. [Google Scholar] [CrossRef]
- DuPont, M.S.; Day, A.J.; Bennett, R.N.; Mellon, F.A.; Kroon, P.A. Absorption of kaempferol from endive, a source of kaempferol-3-glucuronide, in humans. Eur. J. Clin. Nutr. 2004, 58, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Minihane, A.-M. The role of metabolism (and the microbiome) in defining the clinical efficacy of dietary flavonoids. Am. J. Clin. Nutr. 2017, 105, 10–22. [Google Scholar] [CrossRef] [PubMed]
- Cuyckens, F.; Claeys, M. Mass spectrometry in the structural analysis of flavonoids. J. Mass Spectrom. 2004, 39, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Panche, A.N.; Diwan, A.D.; Chandra, S.R. Flavonoids: An overview. J. Nutr. Sci 2016, 5, e47. [Google Scholar] [CrossRef] [PubMed]
- Dabeek, W.M.; Kovinich, N.; Walsh, C.; Ventura Marra, M. Characterization and Quantification of Major Flavonol Glycosides in Ramps (Allium tricoccum). Molecules 2019, 24, 3281. [Google Scholar] [CrossRef] [PubMed]
- Institute of Medicine. Standing Committee on the Scientific Evaluation of Dietary Reference Intakes Dietary Reference Intakes: Proposed Definition and Plan for Review of Dietary Antioxidants and Related Compounds; National Academies Press: Washington, DC, USA, 1998; ISBN 978-0-309-06187-2. [Google Scholar]
- Chun, O.K.; Chung, S.J.; Song, W.O. Estimated Dietary Flavonoid Intake and Major Food Sources of U.S. Adults. J. Nutr. 2007, 137, 1244–1252. [Google Scholar] [CrossRef]
- Cassidy, A.; O’Reilly, É.J.; Kay, C.; Sampson, L.; Franz, M.; Forman, J.; Curhan, G.; Rimm, E.B. Habitual intake of flavonoid subclasses and incident hypertension in adults123. Am. J. Clin. Nutr. 2011, 93, 338–347. [Google Scholar] [CrossRef] [PubMed]
- Kesse-Guyot, E.; Fezeu, L.; Andreeva, V.A.; Touvier, M.; Scalbert, A.; Hercberg, S.; Galan, P. Total and Specific Polyphenol Intakes in Midlife Are Associated with Cognitive Function Measured 13 Years Later. J. Nutr. 2012, 142, 76–83. [Google Scholar] [CrossRef]
- Zamora-Ros, R.; Knaze, V.; Luján-Barroso, L.; Slimani, N.; Romieu, I.; Fedirko, V.; de Magistris, M.S.; Ericson, U.; Amiano, P.; Trichopoulou, A.; et al. Estimated dietary intakes of flavonols, flavanones and flavones in the European Prospective Investigation into Cancer and Nutrition (EPIC) 24 hour dietary recall cohort. Br. J. Nutr. 2011, 106, 1915–1925. [Google Scholar] [CrossRef]
- Bai, W.; Wang, C.; Ren, C. Intakes of total and individual flavonoids by US adults. Int. J. Food Sci. Nutr. 2014, 65, 9–20. [Google Scholar] [CrossRef]
- Scalbert, A.; Williamson, G. Dietary intake and bioavailability of polyphenols. J. Nutr. 2000, 130, 2073S–2085S. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Pandey, A.K. Chemistry and Biological Activities of Flavonoids: An Overview. Sci. World J. 2013, 1–16. Available online: https://www.hindawi.com/journals/tswj/2013/162750/ (accessed on 25 March 2019). [CrossRef] [PubMed]
- Sharma, A.; Sharma, P.; Tuli, H.S.; Sharma, A.K. Phytochemical and Pharmacological Properties of Flavonols. In eLS.; American Cancer Society; Wiley: Hoboken, NJ, USA, 2018; pp. 1–12. ISBN 978-0-470-01590-2. [Google Scholar]
- Jiang, H.; Engelhardt, U.H.; Thräne, C.; Maiwald, B.; Stark, J. Determination of flavonol glycosides in green tea, oolong tea and black tea by UHPLC compared to HPLC. Food Chem. 2015, 183, 30–35. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Muzashvili, T.S.; Georgiev, M.I. Advances in the biotechnological glycosylation of valuable flavonoids. Biotechnol. Adv. 2014, 32, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Murota, K.; Terao, J. Antioxidative flavonoid quercetin: Implication of its intestinal absorption and metabolism. Arch. Biochem. Biophys. 2003, 417, 12–17. [Google Scholar] [CrossRef]
- Lee, J.; Mitchell, A.E. Pharmacokinetics of quercetin absorption from apples and onions in healthy humans. J. Agric. Food Chem. 2012, 60, 3874–3881. [Google Scholar] [CrossRef] [PubMed]
- Thilakarathna, S.H.; Rupasinghe, H.P.V. Flavonoid Bioavailability and Attempts for Bioavailability Enhancement. Nutrients 2013, 5, 3367–3387. [Google Scholar] [CrossRef]
- Makris, D.P.; Kallithraka, S.; Kefalas, P. Flavonols in grapes, grape products and wines: Burden, profile and influential parameters. J. Food Compos. Anal. 2006, 19, 396–404. [Google Scholar] [CrossRef]
- Lu, X.; Ross, C.F.; Powers, J.R.; Rasco, B.A. Determination of Quercetins in Onion (Allium cepa) Using Infrared Spectroscopy. J. Agric. Food Chem. 2011, 59, 6376–6382. [Google Scholar] [CrossRef] [PubMed]
- Dragoni, S.; Gee, J.; Bennett, R.; Valoti, M.; Sgaragli, G. Red wine alcohol promotes quercetin absorption and directs its metabolism towards isorhamnetin and tamarixetin in rat intestine in vitro. Br. J. Pharm. 2006, 147, 765–771. [Google Scholar] [CrossRef]
- Carotenuto, A.; De Feo, V.; Fattorusso, E.; Lanzotti, V.; Magno, S.; Cicala, C. The flavonoids of Allium ursinum. Phytochemistry 1996, 41, 531–536. [Google Scholar] [CrossRef]
- Barile, E.; Capasso, R.; Izzo, A.A.; Lanzotti, V.; Sajjadi, S.E.; Zolfaghari, B. Structure-activity relationships for saponins from Allium hirtifolium and Allium elburzense and their antispasmodic activity. Planta Med. 2005, 71, 1010–1018. [Google Scholar] [CrossRef] [PubMed]
- Fattorusso, E.; Lanzotti, V.; Taglialatela-Scafati, O.; Cicala, C. The flavonoids of leek, Allium porrum. Phytochemistry 2001, 57, 565–569. [Google Scholar] [CrossRef]
- Carotenuto, A.; Fattorusso, E.; Lanzotti, V.; Magno, S.; De Feo, V.; Cicala, C. The flavonoids of Allium neapolitanum. Phytochemistry 1997, 44, 949–957. [Google Scholar] [CrossRef]
- Vallejo, F.; Tomás-Barberán, F.A.; Ferreres, F. Characterisation of flavonols in broccoli (Brassica oleracea L. var. italica) by liquid chromatography–UV diode-array detection–electrospray ionisation mass spectrometry. J. Chromatogr. A 2004, 1054, 181–193. [Google Scholar] [CrossRef] [PubMed]
- Aherne, S.A.; O’Brien, N.M. Dietary flavonols: Chemistry, food content, and metabolism. Nutrition 2002, 18, 75–81. [Google Scholar] [CrossRef]
- Williamson, G.; Kay, C.D.; Crozier, A. The Bioavailability, Transport, and Bioactivity of Dietary Flavonoids: A Review from a Historical Perspective. Compr. Rev. Food Sci. Food Saf. 2018, 17, 1054–1112. [Google Scholar] [CrossRef]
- Alvarez, A.I.; Real, R.; Pérez, M.; Mendoza, G.; Prieto, J.G.; Merino, G. Modulation of the activity of ABC transporters (P-glycoprotein, MRP2, BCRP) by flavonoids and drug response. J. Pharm. Sci. 2010, 99, 598–617. [Google Scholar] [CrossRef]
- Day, A.J.; DuPont, M.S.; Ridley, S.; Rhodes, M.; Rhodes, M.J.C.; Morgan, M.R.A.; Williamson, G. Deglycosylation of flavonoid and isoflavonoid glycosides by human small intestine and liver β-glucosidase activity. FEBS Lett. 1998, 436, 71–75. [Google Scholar] [CrossRef]
- Németh, K.; Plumb, G.W.; Berrin, J.-G.; Juge, N.; Jacob, R.; Naim, H.Y.; Williamson, G.; Swallow, D.M.; Kroon, P.A. Deglycosylation by small intestinal epithelial cell beta-glucosidases is a critical step in the absorption and metabolism of dietary flavonoid glycosides in humans. Eur. J. Nutr. 2003, 42, 29–42. [Google Scholar] [CrossRef]
- Walgren, R.A.; Lin, J.T.; Kinne, R.K.; Walle, T. Cellular uptake of dietary flavonoid quercetin 4′-beta-glucoside by sodium-dependent glucose transporter SGLT1. J. Pharm. Exp. 2000, 294, 837–843. [Google Scholar]
- Manach, C.; Regerat, F.; Texier, O.; Agullo, G.; Demigne, C.; Remesy, C. Bioavailability, metabolism and physiological impact of 4-oxo-flavonoids. Nutr. Res. 1996, 16, 517–544. [Google Scholar] [CrossRef]
- O’Leary, K.A.; Day, A.J.; Needs, P.W.; Mellon, F.A.; O’Brien, N.M.; Williamson, G. Metabolism of quercetin-7- and quercetin-3-glucuronides by an in vitro hepatic model: The role of human beta-glucuronidase, sulfotransferase, catechol-O-methyltransferase and multi-resistant protein 2 (MRP2) in flavonoid metabolism. Biochem. Pharm. 2003, 65, 479–491. [Google Scholar] [CrossRef]
- Marín, L.; Miguélez, E.M.; Villar, C.J.; Lombó, F. Bioavailability of Dietary Polyphenols and Gut Microbiota Metabolism: Antimicrobial Properties. Biomed. Res. Int. 2015, 1–18. Available online: https://www.hindawi.com/journals/bmri/2015/905215/ (accessed on 31 January 2019).
- Lu, Q.-Y.; Zhang, L.; Eibl, G.; Go, V.L.W. Overestimation of flavonoid aglycones as a result of the ex vivo deconjugation of glucuronides by the tissue β-glucuronidase. J. Pharm. Biomed. Anal. 2014, 88, 364–369. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Wolkoff, A.W.; Morris, M.E. Flavonoids as a novel class of human organic anion-transporting polypeptide OATP1B1 (OATP-C) modulators. Drug Metab. Dispos. 2005, 33, 1666–1672. [Google Scholar] [CrossRef] [PubMed]
- Wong, C.C.; Botting, N.P.; Orfila, C.; Al-Maharik, N.; Williamson, G. Flavonoid conjugates interact with organic anion transporters (OATs) and attenuate cytotoxicity of adefovir mediated by organic anion transporter 1 (OAT1/SLC22A6). Biochem. Pharm. 2011, 81, 942–949. [Google Scholar] [CrossRef]
- Manach, C.; Scalbert, A.; Morand, C.; Rémésy, C.; Jiménez, L. Polyphenols: Food sources and bioavailability. Am. J. Clin. Nutr. 2004, 79, 727–747. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Rouanet, J.-M.; Auger, C.; Teissèdre, P.-L.; Caldwell, S.T.; Hartley, R.C.; Lean, M.E.J.; Edwards, C.A.; Crozier, A. Bioavailability of [2-(14)C]quercetin-4′-glucoside in rats. J. Agric. Food Chem. 2008, 56, 12127–12137. [Google Scholar] [CrossRef] [PubMed]
- Viskupicova, J.; Ondrejovič, M.; Sturdik, E. Bioavailability and metabolism of flavonoids. J. Food Nutr. Res. 2008, 47, 151–162. [Google Scholar]
- De Vries, J.H.; Hollman, P.C.; Meyboom, S.; Buysman, M.N.; Zock, P.L.; van Staveren, W.A.; Katan, M.B. Plasma concentrations and urinary excretion of the antioxidant flavonols quercetin and kaempferol as biomarkers for dietary intake. Am. J. Clin. Nutr. 1998, 68, 60–65. [Google Scholar] [CrossRef]
- De Vries, J.H.; Hollman, P.C.H.; van Amersfoort, I.; Olthof, M.R.; Katan, M.B. Red Wine Is a Poor Source of Bioavailable Flavonols in Men. J. Nutr. 2001, 131, 745–748. [Google Scholar] [CrossRef] [PubMed]
- Olthof, M.R.; Hollman, P.C.H.; Vree, T.B.; Katan, M.B. Bioavailabilities of Quercetin-3-Glucoside and Quercetin-4′-Glucoside Do Not Differ in Humans. J. Nutr. 2000, 130, 1200–1203. [Google Scholar] [CrossRef] [PubMed]
- Hollman, P.C.; van Trijp, J.M.; Buysman, M.N.; van der Gaag, M.S.; Mengelers, M.J.; de Vries, J.H.; Katan, M.B. Relative bioavailability of the antioxidant flavonoid quercetin from various foods in man. FEBS Lett. 1997, 418, 152–156. [Google Scholar] [CrossRef]
- Shi, Y.; Williamson, G. Comparison of the urinary excretion of quercetin glycosides from red onion and aglycone from dietary supplements in healthy subjects: A randomized, single-blinded, cross-over study. Food Funct. 2015, 6, 1443–1448. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Mah, E.; Davis, C.G.; Jalili, T.; Ferruzzi, M.G.; Chun, O.K.; Bruno, R.S. Dietary fat increases quercetin bioavailability in overweight adults. Mol. Nutr. Food Res. 2013, 57, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Graefe, E.U.; Wittig, J.; Mueller, S.; Riethling, A.-K.; Uehleke, B.; Drewelow, B.; Pforte, H.; Jacobasch, G.; Derendorf, H.; Veit, M. Pharmacokinetics and Bioavailability of Quercetin Glycosides in Humans. J. Clin. Pharmacol. 2001, 41, 492–499. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.; Liu, R.H. Apple phytochemicals and their health benefits. Nutr. J. 2004, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Palafox-Carlos, H.; Ayala-Zavala, J.F.; González-Aguilar, G.A. The Role of Dietary Fiber in the Bioaccessibility and Bioavailability of Fruit and Vegetable Antioxidants. J. Food Sci. 2011, 76, R6–R15. [Google Scholar] [CrossRef] [PubMed]
- Mullen, W.; Edwards, C.A.; Crozier, A. Absorption, excretion and metabolite profiling of methyl-, glucuronyl-, glucosyl- and sulpho-conjugates of quercetin in human plasma and urine after ingestion of onions. Br. J. Nutr. 2006, 96, 107–116. [Google Scholar] [CrossRef] [PubMed]
- Moon, J.-H.; Nakata, R.; Oshima, S.; Inakuma, T.; Terao, J. Accumulation of quercetin conjugates in blood plasma after the short-term ingestion of onion by women. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2000, 279, R461–R467. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, S.E.; Kall, M.; Justesen, U.; Schou, A.; Dragsted, L.O. Human absorption and excretion of flavonoids after broccoli consumption. Cancer Lett. 1997, 114, 173–174. [Google Scholar] [CrossRef]
- Barve, A.; Chen, C.; Hebbar, V.; Desiderio, J.; Saw, C.L.-L.; Kong, A.-N. Metabolism, oral bioavailability and pharmacokinetics of chemopreventive kaempferol in rats. Biopharm. Drug Dispos. 2009, 30, 356–365. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, K.A.; Day, A.J.; Needs, P.W.; Sly, W.S.; O’Brien, N.M.; Williamson, G. Flavonoid glucuronides are substrates for human liver beta-glucuronidase. FEBS Lett. 2001, 503, 103–106. [Google Scholar] [CrossRef]
- Day, A.J.; Mellon, F.; Barron, D.; Sarrazin, G.; Morgan, M.R.; Williamson, G. Human metabolism of dietary flavonoids: Identification of plasma metabolites of quercetin. Free Radic. Res. 2001, 35, 941–952. [Google Scholar] [CrossRef]
- Jaganath, I.B.; Jaganath, I.B.; Mullen, W.; Edwards, C.A.; Crozier, A. The relative contribution of the small and large intestine to the absorption and metabolism of rutin in man. Free Radic. Res. 2006, 40, 1035–1046. [Google Scholar] [CrossRef] [PubMed]
- Conquer, J.A.; Maiani, G.; Azzini, E.; Raguzzini, A.; Holub, B.J. Supplementation with Quercetin Markedly Increases Plasma Quercetin Concentration without Effect on Selected Risk Factors for Heart Disease in Healthy Subjects. J. Nutr. 1998, 128, 593–597. [Google Scholar] [CrossRef] [PubMed]
- Edwards, R.L.; Lyon, T.; Litwin, S.E.; Rabovsky, A.; Symons, J.D.; Jalili, T. Quercetin reduces blood pressure in hypertensive subjects. J. Nutr. 2007, 137, 2405–2411. [Google Scholar] [CrossRef] [PubMed]
- Egert, S.; Bosy-Westphal, A.; Seiberl, J.; Kürbitz, C.; Settler, U.; Plachta-Danielzik, S.; Wagner, A.E.; Frank, J.; Schrezenmeir, J.; Rimbach, G.; et al. Quercetin reduces systolic blood pressure and plasma oxidised low-density lipoprotein concentrations in overweight subjects with a high-cardiovascular disease risk phenotype: A double-blinded, placebo-controlled cross-over study. Br. J. Nutr. Camb. 2009, 102, 1065–1074. [Google Scholar] [CrossRef]
- Zahedi, M.; Ghiasvand, R.; Feizi, A.; Asgari, G.; Darvish, L. Does Quercetin Improve Cardiovascular Risk factors and Inflammatory Biomarkers in Women with Type 2 Diabetes: A Double-blind Randomized Controlled Clinical Trial. Int. J. Prev. Med. 2013, 4, 777–785. [Google Scholar]
- Brüll, V.; Burak, C.; Stoffel-Wagner, B.; Wolffram, S.; Nickenig, G.; Müller, C.; Langguth, P.; Alteheld, B.; Fimmers, R.; Naaf, S.; et al. Effects of a quercetin-rich onion skin extract on 24 h ambulatory blood pressure and endothelial function in overweight-to-obese patients with (pre-)hypertension: A randomised double-blinded placebo-controlled cross-over trial. Br. J. Nutr. 2015, 114, 1263–1277. [Google Scholar] [CrossRef]
- Serban, M.; Sahebkar, A.; Zanchetti, A.; Mikhailidis, D.P.; Howard, G.; Antal, D.; Andrica, F.; Ahmed, A.; Aronow, W.S.; Muntner, P.; et al. Effects of Quercetin on Blood Pressure: A Systematic Review and Meta-Analysis of Randomized Controlled Trials. J. Am. Heart Assoc. 2016, 5, e002713. [Google Scholar] [CrossRef] [PubMed]
- Larson, A.J.; Symons, J.D.; Jalili, T. Therapeutic Potential of Quercetin to Decrease Blood Pressure: Review of Efficacy and Mechanisms. Adv. Nutr. 2012, 3, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Larson, A.; Witman, M.A.H.; Guo, Y.; Ives, S.; Richardson, R.S.; Bruno, R.S.; Jalili, T.; Symons, J.D. Acute, quercetin-induced reductions in blood pressure in hypertensive individuals are not secondary to lower plasma angiotensin-converting enzyme activity or endothelin-1: Nitric oxide. Nutr. Res. 2012, 32, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Sanz, M.J.; Ferrandiz, M.L.; Cejudo, M.; Terencio, M.C.; Gil, B.; Bustos, G.; Ubeda, A.; Gunasegaran, R.; Alcaraz, M.J. Influence of a series of natural flavonoids on free radical generating systems and oxidative stress. Xenobiotica 1994, 24, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Kampkötter, A.; Gombitang Nkwonkam, C.; Zurawski, R.F.; Timpel, C.; Chovolou, Y.; Wätjen, W.; Kahl, R. Effects of the flavonoids kaempferol and fisetin on thermotolerance, oxidative stress and FoxO transcription factor DAF-16 in the model organism Caenorhabditis elegans. Arch. Toxicol. 2007, 81, 849–858. [Google Scholar] [CrossRef] [PubMed]
- Hertog, M.G.; Feskens, E.J.; Hollman, P.C.; Katan, M.B.; Kromhout, D. Dietary antioxidant flavonoids and risk of coronary heart disease: The Zutphen Elderly Study. Lancet 1993, 342, 1007–1011. [Google Scholar] [CrossRef]
- Huxley, R.R.; Neil, H.A.W. The relation between dietary flavonol intake and coronary heart disease mortality: A meta-analysis of prospective cohort studies. Eur. J. Clin. Nutr. 2003, 57, 904–908. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Rexrode, K.M.; Hu, F.; Albert, C.M.; Chae, C.U.; Rimm, E.B.; Stampfer, M.J.; Manson, J.E. Dietary Intakes of Flavonols and Flavones and Coronary Heart Disease in US Women. Am. J. Epidemiol. 2007, 165, 1305–1313. [Google Scholar] [CrossRef] [PubMed]
- Geleijnse, J.M.; Launer, L.J.; Van der Kuip, D.A.M.; Hofman, A.; Witteman, J.C.M. Inverse association of tea and flavonoid intakes with incident myocardial infarction: The Rotterdam Study. Am. J. Clin. Nutr. 2002, 75, 880–886. [Google Scholar] [CrossRef] [PubMed]
- Marniemi, J.; Alanen, E.; Impivaara, O.; Seppänen, R.; Hakala, P.; Rajala, T.; Rönnemaa, T. Dietary and serum vitamins and minerals as predictors of myocardial infarction and stroke in elderly subjects. Nutr. Metab. Cardiovasc. Dis. 2005, 15, 188–197. [Google Scholar] [CrossRef] [PubMed]
- Bobe, G.; Albert, P.S.; Sansbury, L.B.; Lanza, E.; Schatzkin, A.; Colburn, N.H.; Cross, A.J. Interleukin-6 as a Potential Indicator for Prevention of High Risk Adenoma Recurrence by Dietary Flavonols in the Polyp Prevention Trial. Cancer Prev. Res. (Phila.) 2010, 3, 764–775. [Google Scholar] [CrossRef] [PubMed]
- Knekt, P.; Kumpulainen, J.; Järvinen, R.; Rissanen, H.; Heliövaara, M.; Reunanen, A.; Hakulinen, T.; Aromaa, A. Flavonoid intake and risk of chronic diseases. Am. J. Clin. Nutr. 2002, 76, 560–568. [Google Scholar] [CrossRef] [PubMed]
- GRAS Notice Inventory-Agency Response Letter GRAS Notice No. GRN 000341. Available online: https://wayback.archive-it.org/7993/20171031012354/https://www.fda.gov/Food/IngredientsPackagingLabeling/GRAS/NoticeInventory/ucm235935.htm (accessed on 1 April 2019).
- Pfeuffer, M.; Auinger, A.; Bley, U.; Kraus-Stojanowic, I.; Laue, C.; Winkler, P.; Rüfer, C.E.; Frank, J.; Bösch-Saadatmandi, C.; Rimbach, G.; et al. Effect of quercetin on traits of the metabolic syndrome, endothelial function and inflammation in men with different APOE isoforms. Nutr. Metab. Cardiovasc. Dis. 2013, 23, 403–409. [Google Scholar] [CrossRef] [PubMed]
- McAnulty, S.R.; McAnulty, L.S.; Nieman, D.C.; Quindry, J.C.; Hosick, P.A.; Hudson, M.H.; Still, L.; Henson, D.A.; Milne, G.L.; Morrow, J.D.; et al. Chronic quercetin ingestion and exercise-induced oxidative damage and inflammation. Appl. Physiol. Nutr. Metab. 2008, 33, 254–262. [Google Scholar] [CrossRef] [PubMed]
- Lu, N.T.; Crespi, C.M.; Liu, N.M.; Vu, J.Q.; Ahmadieh, Y.; Wu, S.; Lin, S.; McClune, A.; Durazo, F.; Saab, S.; et al. A Phase I Dose Escalation Study Demonstrates Quercetin Safety and Explores Potential for Bioflavonoid Antivirals in Patients with Chronic Hepatitis C. Phytother. Res. 2016, 30, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Pietta, P.G. Flavonoids as antioxidants. J. Nat. Prod. 2000, 63, 1035–1042. [Google Scholar] [CrossRef] [PubMed]
- Terao, J. Dietary Flavonoids as Antioxidants. Food Factors Health Promot. 2009, 61, 87–94. [Google Scholar]
- Nirmala, P.; Ramanathan, M. Effect of kaempferol on lipid peroxidation and antioxidant status in 1,2-dimethyl hydrazine induced colorectal carcinoma in rats. Eur. J. Pharm. 2011, 654, 75–79. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Cheng, Z.; Heller, L.I.; Krasnoff, S.B.; Glahn, R.P.; Welch, R.M. Kaempferol in Red and Pinto Bean Seed (Phaseolus vulgaris L.) Coats Inhibits Iron Bioavailability Using an in Vitro Digestion/Human Caco-2 Cell Model. J. Agric. Food Chem. 2006, 54, 9254–9261. [Google Scholar] [CrossRef] [PubMed]
- Lemos, C.; Peters, G.J.; Jansen, G.; Martel, F.; Calhau, C. Modulation of folate uptake in cultured human colon adenocarcinoma Caco-2 cells by dietary compounds. Eur. J. Nutr. 2007, 46, 329–336. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Source | Quercetin | Kaempferol |
|---|---|---|
| Food | mg/100 g fresh weight | |
| Apples | 4.01 | 0.14 |
| Asparagus | 14.0 | 1.40 |
| Broccoli | 13.7 | 7.20 |
| Chili pepper | 32.6 | - |
| Chinese cabbage | - | 22.5 |
| Kale | 22.6 | 47.0 |
| Leeks | 0.9 | 2.67 |
| Lettuce | 14.7 | 0.84 |
| Onions | 45.0 | 4.50 |
| Spinach | 27.2 | 55.0 |
| Chives | 10.4 | 12.5 |
| Dill | 79.0 | 40.0 |
| Fennel leaves | 46.8 | 6.50 |
| Oregano | 42.0 | - |
| Blueberry | 14.6 | 3.17 |
| Cherry | 17.4 | 5.14 |
| Cranberry | 25.0 | 0.21 |
| Wild leeks (whole) [17] | 8.36 | 5.31 |
| Beverage | mg/100 ml | |
| Black tea | 2.50 | 1.70 |
| Red wine | 3.16 | 0.25 |
| Food Source | Sugar Moieties | Ref. |
|---|---|---|
| Quercetin | ||
| Red wine | -3-glucoside, –rutinoside | [32] |
| Onions | -4′-glucoside, -3,4′-diglucoside | [33] |
| Tea | -3-rutinoside | [34] |
| Apple | -rutinoside, -galactoside, -rhamnoside, –glucoside | [30] |
| Wild leek (ramps) | -sophoroside glucuronide, -hexoside glucuronide, -sophoroside | [17] |
| Kaempferol | ||
| Wild garlic and leeks | -glucopyranoside, -neohesperidose | [35,36,37,38] |
| Black tea | -rutinoside, glucoside | [27] |
| Broccoli | -sophoroside (β-1,2-glucose) | [39] |
| Endive | -3- glucuronide, 3-glucoside | [13] |
| Wild leek (ramps) | -sophoroside glucuronide, -rutinoside glucuronide, -sophoroside | [17] |
| Food Source (amount, g or ml) | Quercetin Dosage | n | Urinary Excretion or Concentration (%) | Maximal Plasma Concentration (µmol/L) | Time to Reach Maximal Concentration (hours) | Ref. |
|---|---|---|---|---|---|---|
| Black tea (1600) | 49 mg | 15 | 0.5 | [55] | ||
| Onions (129) | 13 mg | 1.1 | ||||
| Onions (NR) | 225 µmol | 9 | 1.39 | 0.74 | 0.70 | [58] |
| Applesauce + peel (NR) | 325 µmol | 0.44 | 0.30 | 2.5 | ||
| Rutin | 331 µmol | 0.35 | 0.30 | 9 | ||
| Onions (333) | 89 mg | 9 | 52 | [11] | ||
| Rutin | 220 mg | 17 | ||||
| Dehydrate | 112 | 24 | ||||
| Red wine (750) | 14.2 | 12 | 0.371 μmol/L | 0.026 | [56] | |
| Onions (50) | 15.9 | 0.509 μmol/L | 0.053 | |||
| Black tea (375) | 13.7 | 0.252 μmol/L | 0.026 | |||
| Quercetin-3-glucoside capsule | 151 | 9 | 3.0 | 5.0 | 0.62 | [57] |
| Quercetin-4′-glucoside capsule | 154 | 2.6 | 4.5 | 0.45 | ||
| Dehydrate | 544 | 6 | 1.69 μmol/L | [59] | ||
| Onion soup (100) | 47 | 1.17 μmol/L | ||||
| Dehydrate with fat-free (<0.5) | 1095 | 9 | - | 1.1 | 5.7 | [60] |
| Dehydrate low-fat (4.0) | - | 1.24 | 5.4 | |||
| Dehydrate high-fat (15.4) | - | 1.6 | 6.7 | |||
| Onions (160) | 100 | 12 | 6.4 | 2.31 | 0.68 | [61] |
| Dehydrate | 100 | 4.5 | 2.12 | 0.70 | ||
| Buckwheat tea (NR) | 200 | 1.0 | 0.64 | 4.32 | ||
| Dehydrate | 200 | 0.90 | 0.32 | 6.98 |
| Food Source (g or ml) | Flavonol Glycoside (mg) | Dosage (mg) | Metabolites Detected | Concentration (µmol/L) | Max Time (hours) | Half-life (hours) | Ref. |
|---|---|---|---|---|---|---|---|
| Blood | |||||||
| Fried onions (200) | Quercetin-3,4′-O-diglucoside | 37.1 | Isorhamnetine | 0.11 | 1.5 | - | [69] |
| Quercetin-3-O-glucoside | 0.7 | -glucuronide | 0.26 | 1.5 | - | ||
| Quercetin-4′-O-glucoside | 39.5 | -Sulfate | 0.16 | 1.5 | - | ||
| Isorhamnetin-4′-O-glucoside | 1.8 | ||||||
| Quercetin | 0.1 | ||||||
| Fried onions (270) | Quercetin-3,4′-O-diglucoside | 67 | -3′-sulfate | 0.67 | 0.75 | 1.71 | [64] |
| Quercetin-4′-O-glucoside | 66 | -3-glucoronide | 0.35 | 0.60 | 2.33 | ||
| Isprhamnetin-4′-O-glucoside * | 5.3 | Isorhamnetin-3-glucuronide* | 0.11 | 0.60 | 5.34 | ||
| -glucuronide sulfate | 0.12 | 2.5 | 4.54 | ||||
| -diglucuronide | 0.062 | 0.80 | 1.76 | ||||
| Tomato juice (300) | Quercetin-3-O-rutinoside | 7.3 | -3-glucuronide | 0.0038 | 5 | 5.7 | [70] |
| Urine | |||||||
| Fried onions (270) | Quercetin-3,4′-O-diglucoside | 67 | -diglucuronide | 2.22 | 4-8 | - | [64] |
| quercetin-3′-glucuronide | 1.85 | 0-4 | - | ||||
| Quercetin-4′-O-glucoside | 66 | isorhamnetin-3-glucuronide* | 1.79 | 4-8 | - | ||
| -glucuronide sulfate | 1.38 | 0-4 | - | ||||
| Methylquercetin diglucuronide | 1.00 | 4-8 | - | ||||
| -3-glucuronide | 0.912 | 0-4 | - | ||||
| Isorhamnetin-4′-O-glucoside * | 5.3 | -glucoside sulfate | 0.82 | 0-4 | - | ||
| Isorhamnetin-4′-glucuronide* | 0.70 | 0-4 | - | ||||
| -glucoronide glucoside | 0.16 | 0-4 | - | ||||
| -4′-O-glucuronide | 0.24 | 24 | - | ||||
| Tomato juice (300) | Quercetin-3-O-rutinoside | 7.3 | -3-glucuronide | 0.18 | 24 | - | [70] |
| n | Health Condition | Age (years) | BMI (kg/m2) | Quercetin (mg) | Duration (days) | Blood Pressure Results | Other Results | Ref. |
|---|---|---|---|---|---|---|---|---|
| 27 | Healthy | 42.0 ± 2.6 | 26.0 ± 1.3 | 1000 aglycone + 200 rutin | 28 | No effect | No effect other CVD factors | [71] |
| 41 | Pre-HTN | 47.8 ± 3.5 | 29.7 ± 1.3 | 730 aglycone | 28 | No effect | nor oxidative stress | [72] |
| Stage 1-HTN | 49.2 ± 2.9 | 29.7 ± 1.3 | SBP (−7 ± 2mmHg), DBP(−5 ± 2mmHg) mean arterial pressures (−5 ± 2 mmHg) | no effect on oxidative stress | ||||
| 93 | Baseline mean BP (130±16.4/ 81.6±9.3mmHg) | 25–65 | 25–35 | 150 aglycone | 42 | SBP: entire group (−2.6 mmHg), subgroup with HTN (−2.9 mmHg) Subgroup aged 25-50 (−3.7 mmHg) | Decreased oxidized LDL. No effect on TNF-α and CRP | [73] |
| 62 | Type 2 diabetes | 35-55 | NR | 500 aglycone | 70 | SBP (−8.8 ± 9.3 mmHg), DBP (no effect) | Decreased TNF-α and IL-6 relative to baseline, but not different than placebo | [74] |
| 68 | Pre-HTN (≥120–139 mmHg and/or ≥80–89 mmHg) | 25–65 | 25–35 | 396 onion powder (162 quercetin glucoside) | 42 | ABP (no effect) | sICAM-1 (−8.2 ng/mL) | [75] |
| Stage I HTN (≥140–159 mmHg and/or ≥90–99) | 25–65 | 25–35 | Systolic ABP (−3.6 mmHg), | sICAM-1 (−8.2 ng/mL) No effect on NO, ACE, sVCAM | ||||
| 5 | Normotensive | 24 ± 3 | 24 ± 4 | 1095 aglycone | 1 | No effect | No effect on ACE, ET-1, NO, and brachial artery flow mediated dilation | [78] |
| 12 | Stage 1 HTN | 41 ± 12 | 29 ± 5 | SBP (−5 mmHg) |
| n | Age (years) | Health Status | Dietary Intake (mg/day) | Results | Ref. |
|---|---|---|---|---|---|
| 805 | 65–84 | Healthy | Flavonoids (12.0–41.6) | (-): CHD mortality (0.42, 95% CI 0.20–0.88, p = 0.015) | [81] |
| 4807 | 64–69 | Healthy | Flavonols (14.8–38.5) | (-): fatal MI (0.57, 95% CI 0.33–0.98) | [84] |
| 66,360 | 30–55 | 29.8% HTN | flavonols+flavone (21.2) Kaempferol (4.7) | (-): CHD mortality (0.66, 95% CI 0.48–0.93; p = 0.04) No association with MI | [83] |
| 10,054 | Mean 39.3 | 9.6% HTN | flavonoids (24.2), kaempferol (0.1–0.9) | (-): cerebrovascular disease (0.70, 95% CI 0.56–0.86, p = 0.003), thrombosis (0.63, 95% CI 0.47–0.85, p = 0.004) | [87] |
| 872 | 52–67 | Colorectal adenoma | Flavonols (9.4–20.8), kaempferol (2.54–8.04) | (-): IL-6 levels (95% CI: 0.24–0.93; p = 0.03) | [86] |
| 744 | 65–99 | Healthy | Kaempferol (1.0–1.5) | (-): acute MI (0.48, 95% CI; 0.30–0.77, p = 0.002). | [85] |
| n | Health Status | Dosage (mg) | Duration (weeks) | Adverse Event | Ref. |
|---|---|---|---|---|---|
| 93 | Normotensive and Stage I hypertension | 150 | 6 | No effects on liver, kidneys, electrolytes, and hematology biomarkers | [73] |
| 49 | Healthy | 150 | 8 | Increased TNF-α by 0.11 pg/mL, p < 0.05 | [89] |
| 40 | Athletes | 1000 | 6 | No adverse events reported | [90] |
| 30 | Chronic Hepatitis C | 250–5000 | 4 | No adverse effects on liver function but mild gastrointestinal discomfort | [91] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dabeek, W.M.; Marra, M.V. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients 2019, 11, 2288. https://doi.org/10.3390/nu11102288
Dabeek WM, Marra MV. Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients. 2019; 11(10):2288. https://doi.org/10.3390/nu11102288
Chicago/Turabian StyleDabeek, Wijdan M., and Melissa Ventura Marra. 2019. "Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans" Nutrients 11, no. 10: 2288. https://doi.org/10.3390/nu11102288
APA StyleDabeek, W. M., & Marra, M. V. (2019). Dietary Quercetin and Kaempferol: Bioavailability and Potential Cardiovascular-Related Bioactivity in Humans. Nutrients, 11(10), 2288. https://doi.org/10.3390/nu11102288