Inflammation and Oxidative Stress in an Obese State and the Protective Effects of Gallic Acid

 ,
,  ,
,
Abstract
1. Introduction
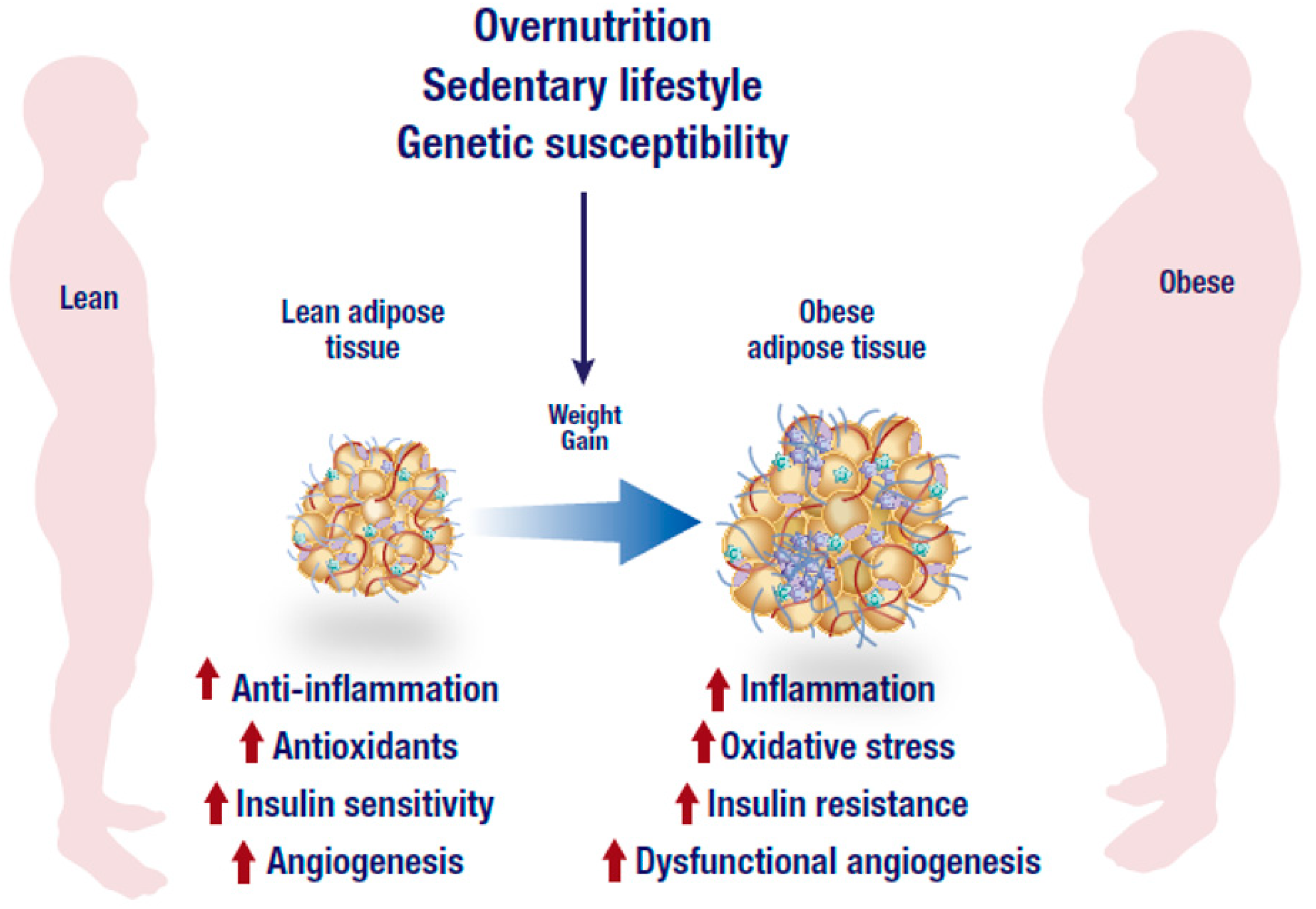
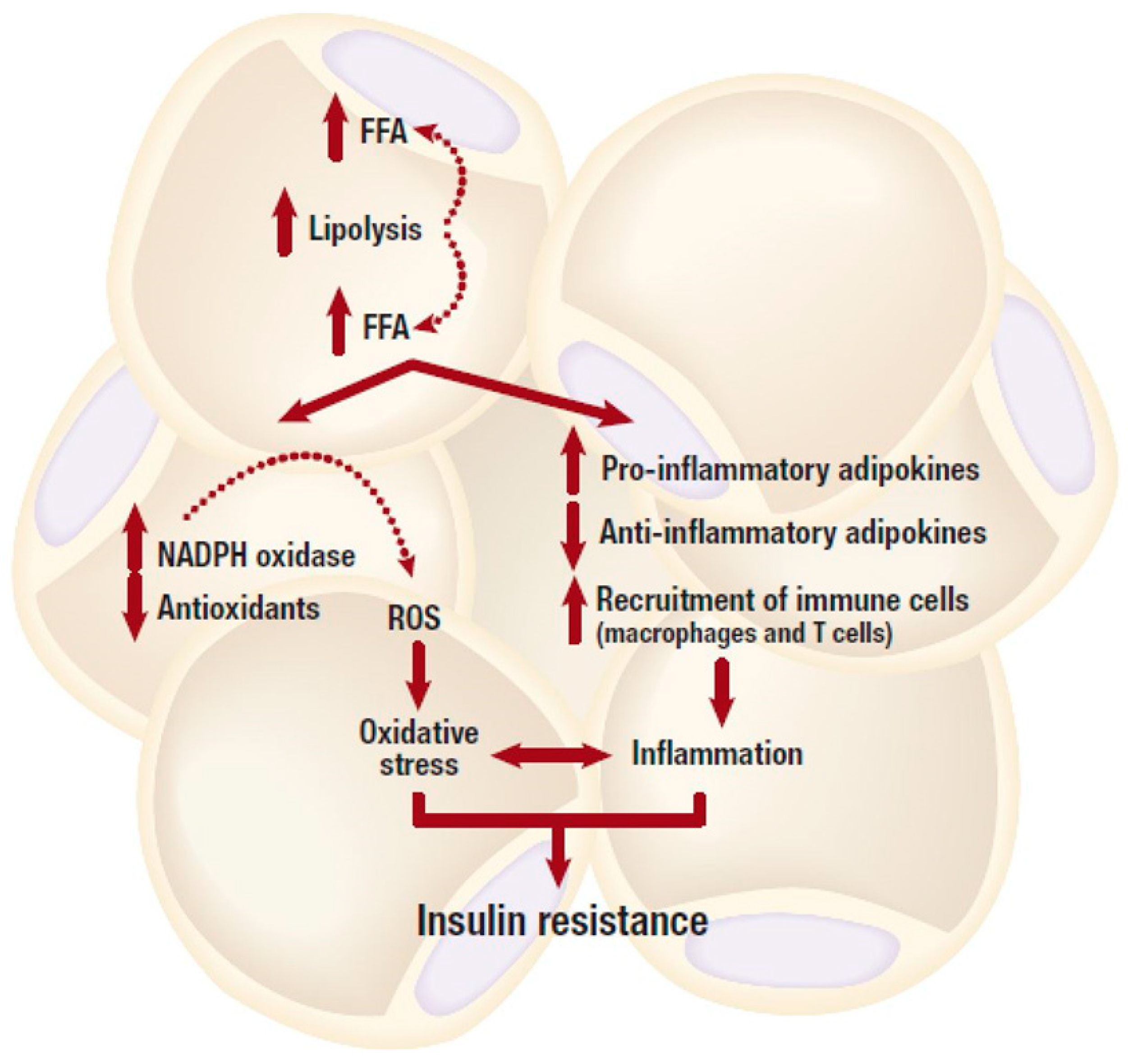
2. Inflammation and Insulin Resistance in Adipose Tissue
3. Oxidative Stress in Adipose Tissue
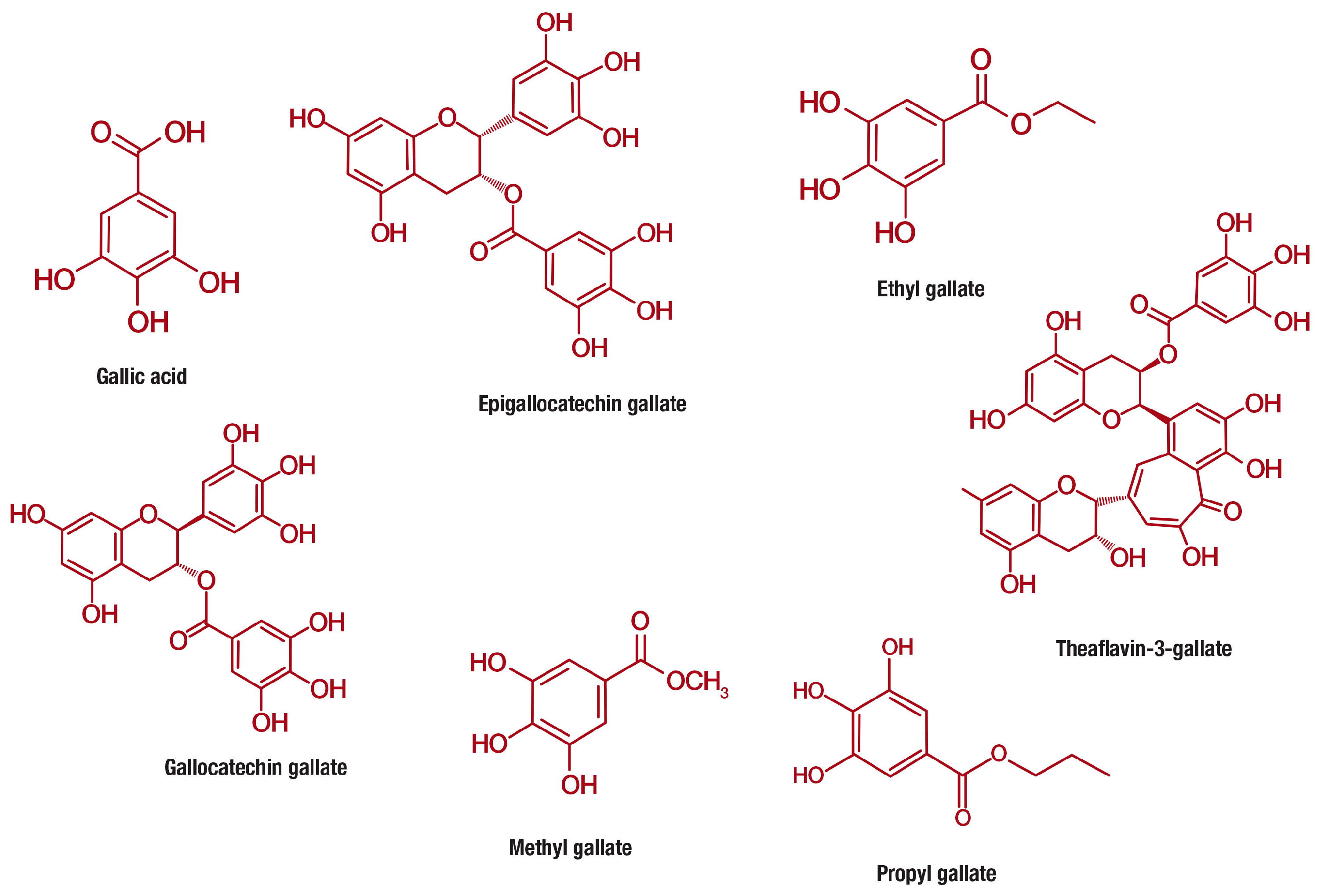
4. A Brief Overview of the Classification, Occurrence, and Bioavailability of Gallic Acid
5. Experimental Models Investigating the Anti-Obesity Effects of Gallic Acid
6. Evidence on the Anti-Obesity Properties of Gallic Acid
7. Evidence on the Anti-Obesity Effects of Gallic Acid Derived Compounds
8. Evidence on the Anti-Obesity Properties of Tea and Fruits Containing Gallic Acid
9. Anti-Obesity Properties of other Plants Rich in Gallic Acid
10. Human Studies Reporting on the Therapeutic Potential of Gallic Acid against Obesity-Associated Complications
11. Concluding Remarks
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AMPK | AMP-activated protein kinase |
| ·OH | hydroxyl radical |
| O2 • − | superoxide anion |
| C/EBP | CCAAT/enhancer-binding proteins |
| FAS | fatty acid synthase |
| FASL | FAS ligand |
| FFAs | free fatty acids |
| HFD | high fat diet |
| H2O2 | hydrogen peroxide |
| iNOS | inducible nitric oxide synthase |
| IKK | inhibitor κB kinase |
| JNK | c-JUN NH2-terminal kinase |
| IRS-1 | insulin receptor substrate 1 |
| IL-6 | interleukin |
| MCP-1 | monocyte chemoattractant 1 |
| NO | nitric oxide |
| NOX | nicotinamide adenine dinucleotide phosphate (NADPH) oxidase |
| NF-κB | nuclear factor kappa-light -chain-enhancer of activated B cells |
| PPAR | peroxisome proliferator-activated receptor |
| PGC1α | peroxisome proliferator activated receptor gamma coactivator 1 alpha |
| PI3K | phosphatidylinositol 3-kinase |
| PKC | phosphokinase C |
| Akt | protein kinase B |
| ROS | reactive oxygen species |
| SIRT1 | NAD-dependent deacetylase sirtuin-1 |
| TH17 | T-helper cell 17 |
| TNF-α | tumor necrosis factor alpha |
| p53 | tumor protein 53 |
| T2D | type 2 diabetes |
References
- World Health Organization. Obesity. Available online: http://www.who.int/en/news-room/fact- sheets/detail/obesity-and-overweight (accessed on 9 August 2018).
- Williams, C.B.; Mackenzie, K.C.; Gahagan, S. The effect of maternal obesity on the offspring. Clin. Obstet. Gynecol. 2014, 57, 508–515. [Google Scholar] [CrossRef] [PubMed]
- Jung, U.J.; Choi, M.S. Obesity and its metabolic complications: The role of adipokines and the relationship between obesity, inflammation, insulin resistance, dyslipidemia and nonalcoholic fatty liver disease. Int. J. Mol. Sci. 2014, 15, 6184–6223. [Google Scholar] [CrossRef] [PubMed]
- Francisqueti, F.V.; Chiaverini, L.C.; Santos, K.C.; Minatel, I.O.; Ronchi, C.B.; Ferron, A.J.; Ferreira, A.L.; Corrêa, C.R. The role of oxidative stress on the pathophysiology of metabolic syndrome. Rev. Assoc. Med. Bras. 2017, 63, 85–91. [Google Scholar] [CrossRef] [PubMed]
- Lumeng, C.N.; Saltiel, A.R. Inflammatory links between obesity and metabolic disease. J. Clin. Investig. 2011, 121, 2111–2117. [Google Scholar] [CrossRef]
- Jo, J.; Gavrilova, O.; Pack, S.; Jou, W.; Mullen, S.; Sumner, A.E.; Cushman, S.W.; Periwal, V. Hypertrophy and/or hyperplasia: Dynamics of adipose tissue growth. PLoS Comput. Biol. 2009, 5, e1000324. [Google Scholar] [CrossRef] [PubMed]
- Ussar, S.; Lee, K.Y.; Dankel, S.N.; Boucher, J.; Haering, M.F.; Kleinridders, A.; Thomou, T.; Xue, R.; Macotela, Y.; Cypess, A.M.; et al. ASC-1, PAT2, and P2RX5 are cell surface markers for white, beige, and brown adipocytes. Sci. Transl. Med. 2014, 6, 247ra103. [Google Scholar] [CrossRef] [PubMed]
- Fernandez-Sanchez, A.; Madrigal-Santillán, E.; Bautista, M.; Esquivel-Soto, J.; Morales-González, A.; Esquivel-Chirino, C.; Durante-Montiel, I.; Sánchez-Rivera, G.; Valadez-Vega, C.; Morales-González, J.A. Inflammation, oxidative stress, and obesity. Int. J. Mol. Sci. 2011, 12, 3117–3132. [Google Scholar] [CrossRef]
- Petrovska, B.B. Historical review of medicinal plants’ usage. Pharmacogn. Rev. 2012, 6, 1–5. [Google Scholar] [CrossRef]
- Lin, D.; Xiao, M.; Zhao, J.; Li, Z.; Xing, B.; Li, X.; Kong, M.; Li, L.; Zhang, Q.; Liu, Y.; et al. An overview of plant phenolic compounds and their importance in human nutrition and management of type 2 diabetes. Molecules 2016, 21, 10. [Google Scholar] [CrossRef]
- Esposito, D.; Damsud, T.; Wilson, M.; Grace, M.H.; Strauch, R.; Li, X.; Lila, M.A.; Komarnytsky, S. Black currant anthocyanins attenuate weight gain and improve glucose metabolism in diet-induced obese mice with intact, but not disrupted, gut microbiome. J. Agric. Food Chem. 2015, 63, 6172–6180. [Google Scholar] [CrossRef]
- Saibabu, V.; Fatima, Z.; Khan, L.A.; Hameed, S. Therapeutic potential of dietary phenolic acids. Adv. Pharmacol. Sci. 2015, 2015, 823539. [Google Scholar] [CrossRef]
- Wang, S.S.; Wang, D.M.; Pu, W.J.; Li, D.W. Phytochemical profiles, antioxidant and antimicrobial activities of three Potentilla species. BMC Complement. Altern. Med. 2013, 13, 321. [Google Scholar] [CrossRef] [PubMed]
- Ediriweera, M.K.; Tennekoon, K.H.; Samarakoon, S.R.; Thabrew, I.; de Silva, E.D. Protective effects of six selected dietary compounds against leptin-induced proliferation of oestrogen receptor positive (MCF-7) breast cancer cells. Medicines 2017, 4, 56. [Google Scholar] [CrossRef]
- Ikeda, I.; Hamamoto, R.; Uzu, K.; Imaizumi, K.; Nagao, K.; Yanagita, T.; Suzuki, Y.; Kobayashi, M.; Kakuda, T. Dietary gallate esters of tea catechins reduce deposition of visceral fat, hepatic triacylglycerol, and activities of hepatic enzymes related to fatty acid synthesis in rats. Biosci. Biotechnol. Biochem. 2005, 69, 1049–1053. [Google Scholar] [CrossRef] [PubMed]
- Makihara, H.; Koike, Y.; Ohta, M.; Horiguchi-Babamoto, E.; Tsubata, M.; Kinoshita, K.; Akase, T.; Goshima, Y.; Aburada, M.; Shimada, T. Gallic acid, the active ingredient of Terminalia bellirica, enhances adipocyte differentiation and adiponectin secretion. Biol. Pharm. Bull. 2016, 39, 1137–1143. [Google Scholar] [CrossRef]
- Pandey, A.; Bani, S.; Sangwan, P.L. Anti-obesity potential of gallic acid from Labisia pumila, through augmentation of adipokines in high fat diet induced obesity in C57BL/6 mice. Adv. Res. 2014, 2, 556–570. [Google Scholar] [CrossRef]
- Chao, J.; Huo, T.I.; Cheng, H.Y.; Tsai, J.C.; Liao, J.W.; Lee, M.S.; Qin, X.M.; Hsieh, M.T.; Pao, L.H.; Peng, W.H. Gallic acid ameliorated impaired glucose and lipid homeostasis in high fat diet-induced NAFLD mice. PLoS ONE 2014, 9, e96969. [Google Scholar] [CrossRef]
- Donado-Pestana, C.M.; Dos Santos-Donado, P.R.; Daza, L.D.; Belchior, T.; Festuccia, W.T.; Genovese, M.I. Cagaita fruit (Eugenia dysenterica DC.) and obesity: Role of polyphenols on already established obesity. Food Res. Int. 2018, 103, 40–47. [Google Scholar] [CrossRef]
- Gandhi, G.R.; Jothi, G.; Antony, P.J.; Balakrishna, K.; Paulraj, M.G.; Ignacimuthu, S.; Stalin, A.; Al-Dhabi, N.A. Gallic acid attenuates high-fat diet fed-streptozotocin-induced insulin resistance via partial agonism of PPARgamma in experimental type 2 diabetic rats and enhances glucose uptake through translocation and activation of GLUT4 in PI3K/p-Akt signaling pathway. Eur. J. Pharmacol. 2014, 745, 201–216. [Google Scholar] [CrossRef]
- Hsu, C.L.; Lo, W.H.; Yen, G.C. Gallic acid induces apoptosis in 3T3-L1 pre-adipocytes via a fas- and mitochondrial-mediated pathway. J. Agric. Food Chem. 2007, 55, 7359–7365. [Google Scholar] [CrossRef]
- Totani, N.; Tateishi, S.; Takimoto, T.; Maeda, Y.; Sasaki, H. Gallic acid glycerol ester promotes weight-loss in rats. J. Oleo. Sci. 2011, 60, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Locatelli, C.; Filippin-Monteiro, F.B.; Creczynski-Pasa, T.B. Alkyl esters of gallic acid as anticancer agents: A review. Eur. J. Med. Chem. 2013, 60, 233–239. [Google Scholar] [CrossRef] [PubMed]
- Badhani, B.; Sharma, N.; Kakkar, R. Gallic acid: A versatile antioxidant with promising therapeutic and industrial applications. RSC Adv. 2015, 5, 27540–27557. [Google Scholar] [CrossRef]
- Choubey, S.; Varughese, L.R.; Kumar, V.; Beniwal, V. Medicinal importance of gallic acid and its ester derivatives: A patent review. Pharm. Pat. Anal. 2015, 4, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, F.H.; Salgado, H.R. Gallic acid: Review of the methods of determination and quantification. Crit. Rev. Anal. Chem. 2016, 46, 257–265. [Google Scholar] [CrossRef] [PubMed]
- Nayeem, N.; SMB, A.; Salem, H.; AHEl-Alfqy, S. Gallic acid: A promising lead molecule for drug development. J. Appl. Pharm. 2016, 8, 213. [Google Scholar] [CrossRef]
- Kosuru, R.Y.; Roy, A.; Das, S.K.; Bera, S. Gallic acid and gallates in human health and disease: Do mitochondria hold the key to success? Mol. Nutr. Food Res. 2018, 62. [Google Scholar] [CrossRef]
- Dludla, P.V.; Nkambule, B.B.; Dias, S.C.; Johnson, R. Cardioprotective potential of N-acetyl cysteine against hyperglycaemia-induced oxidative damage: A protocol for a systematic review. Syst. Rev. 2017, 6, 96. [Google Scholar] [CrossRef]
- Weisberg, S.P.; McCann, D.; Desai, M.; Rosenbaum, M.; Leibel, R.L.; Ferrante, A.W., Jr. Obesity is associated with macrophage accumulation in adipose tissue. J. Clin. Investig. 2003, 112, 1796–1808. [Google Scholar] [CrossRef]
- Cao, Y. Angiogenesis modulates adipogenesis and obesity. J. Clin. Investig. 2007, 117, 2362–2368. [Google Scholar] [CrossRef]
- Ye, J. Adipose tissue vascularization: Its role in chronic inflammation. Curr. Diab. Rep. 2011, 11, 203–210. [Google Scholar] [CrossRef] [PubMed]
- Hausman, G.J.; Richardson, R.L. Adipose tissue angiogenesis. J. Anim. Sci. 2004, 82, 925–934. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y. Angiogenesis and vascular functions in modulation of obesity, adipose metabolism, and insulin sensitivity. Cell MeTable 2013, 18, 478–489. [Google Scholar] [CrossRef] [PubMed]
- Sears, B.; Perry, M. The role of fatty acids in insulin resistance. Lipids Health Dis. 2015, 14, 121. [Google Scholar] [CrossRef] [PubMed]
- Stolarczyk, E. Adipose tissue inflammation in obesity: A metabolic or immune response? Curr. Opin. Pharmacol. 2017, 37, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Hao, Z.; Münzberg, H.; Rezai-Zadeh, K.; Keenan, M.; Coulon, D.; Lu, H.; Berthoud, H.R.; Ye, J. Leptin deficient ob/ob mice and diet-induced obese mice responded differently to Roux-en-Y bypass surgery. Int. J. Obes. 2015, 39, 798–805. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef]
- Katsuki, A.; Sumida, Y.; Murashima, S.; Murata, K.; Takarada, Y.; Ito, K.; Fujii, M.; Tsuchihashi, K.; Goto, H.; Nakatani, K.; et al. Serum levels of tumor necrosis factor-alpha are increased in obese patients with noninsulin-dependent diabetes mellitus. J. Clin. Endocrinol. MeTable 1998, 83, 859–862. [Google Scholar] [CrossRef]
- Perego, L.; Pizzocri, P.; Corradi, D.; Maisano, F.; Paganelli, M.; Fiorina, P.; Barbieri, M.; Morabito, A.; Paolisso, G.; Folli, F.; et al. Circulating leptin correlates with left ventricular mass in morbid (grade III) obesity before and after weight loss induced by bariatric surgery: A potential role for leptin in mediating human left ventricular hypertrophy. J. Clin. Endocrinol. MeTable 2005, 90, 4087–4093. [Google Scholar] [CrossRef]
- Dludla, P.V.; Essop, M.F.; Gabuza, K.B.; Muller, C.J.F.; Louw, J.; Johnson, R. Age-dependent development of left ventricular wall thickness in type 2 diabetic (db/db) mice is associated with elevated low-density lipoprotein and triglyceride serum levels. Heart Vessels 2017, 32, 1025–1031. [Google Scholar] [CrossRef]
- Mangge, H.; Summers, K.L.; Meinitzer, A.; Zelzer, S.; Almer, G.; Prassl, R.; Schnedl, W.J.; Reininghaus, E.; Paulmichl, K.; Weghuber, D.; et al. Obesity-related dysregulation of the tryptophan-kynurenine metabolism: Role of age and parameters of the metabolic syndrome. Obesity 2014, 22, 195–201. [Google Scholar] [CrossRef] [PubMed]
- Brandacher, G.; Hoeller, E.; Fuchs, D.; Weiss, H.G. Chronic immune activation underlies morbid obesity: Is IDO a key player? Curr. Drug MeTable 2007, 8, 289–295. [Google Scholar] [CrossRef]
- Curat, C.A.; Wegner, V.; Sengenès, C.; Miranville, A.; Tonus, C.; Busse, R.; Bouloumié, A. Macrophages in human visceral adipose tissue: Increased accumulation in obesity and a source of resistin and visfatin. Diabetologia 2006, 49, 744–747. [Google Scholar] [CrossRef] [PubMed]
- Nio, Y.; Yamauchi, T.; Iwabu, M.; Okada-Iwabu, M.; Funata, M.; Yamaguchi, M.; Ueki, K.; Kadowaki, T. Monocyte chemoattractant protein-1 (MCP-1) deficiency enhances alternatively activated M2 macrophages and ameliorates insulin resistance and fatty liver in lipoatrophic diabetic A-ZIP transgenic mice. Diabetologia 2012, 55, 3350–3358. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, K.; Hori, M.; Ouchi, N.; Kihara, S.; Funahashi, T.; Matsuzawa, Y.; Miyazaki, A.; Nakayama, H.; Horiuchi, S. Adiponectin down-regulates acyl-coenzyme A: Cholesterol acyltransferase-1 in cultured human monocyte-derived macrophages. Biochem. Biophys. Res. Commun. 2004, 317, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Ehsan, M.; Singh, K.K.; Lovren, F.; Pan, Y.; Quan, A.; Mantella, L.E.; Sandhu, P.; Teoh, H.; Al-Omran, M.; Verma, S. Adiponectin limits monocytic microparticle-induced endothelial activation by modulation of the AMPK, Akt and NFkappaB signaling pathways. Atherosclerosis 2016, 245, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Saucillo, D.C.; Gerriets, V.A.; Sheng, J.; Rathmell, J.C.; Maciver, N.J. Leptin metabolically licenses T cells for activation to link nutrition and immunity. J. Immunol. 2014, 192, 136–144. [Google Scholar] [CrossRef]
- Ip, B.; Cilfone, N.A.; Belkina, A.C.; DeFuria, J.; Jagannathan-Bogdan, M.; Zhu, M.; Kuchibhatla, R.; McDonnell, M.E.; Xiao, Q.; Kepler, T.B.; et al. Th17 cytokines differentiate obesity from obesity-associated type 2 diabetes and promote TNFalpha production. Obesity 2016, 24, 102–112. [Google Scholar] [CrossRef]
- Gonzalez, Y.; Herrera, M.T.; Soldevila, G.; Garcia-Garcia, L.; Fabián, G.; Pérez-Armendariz, E.M.; Bobadilla, K.; Guzmán-Beltrán, S.; Sada, E.; Torres, M. High glucose concentrations induce TNF-alpha production through the down-regulation of CD33 in primary human monocytes. BMC Immunol. 2012, 13, 19. [Google Scholar] [CrossRef]
- Lajaunias, F.; Dayer, J.M.; Chizzolini, C. Constitutive repressor activity of CD33 on human monocytes requires sialic acid recognition and phosphoinositide 3-kinase-mediated intracellular signaling. Eur. J. Immunol. 2005, 35, 243–251. [Google Scholar] [CrossRef]
- Jin, W.; Dong, C. IL-17 cytokines in immunity and inflammation. Emerg. Microbes. Infect. 2013, 2, e60. [Google Scholar] [CrossRef] [PubMed]
- Capurso, C.; Capurso, A. From excess adiposity to insulin resistance: The role of free fatty acids. Vascul. Pharmacol. 2012, 57, 91–97. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.; Dludla, P.V.; Muller, C.J.; Huisamen, B.; Essop, M.F.; Louw, J. The transcription profile unveils the cardioprotective effect of aspalathin against lipid toxicity in an in vitro H9c2 model. Molecules 2017, 22, 22. [Google Scholar] [CrossRef] [PubMed]
- Glass, C.K.; Olefsky, J.M. Inflammation and lipid signaling in the etiology of insulin resistance. Cell MeTable 2012, 15, 635–645. [Google Scholar] [CrossRef] [PubMed]
- Johnson, A.M.; Olefsky, J.M. The origins and drivers of insulin resistance. Cell 2013, 152, 673–684. [Google Scholar] [CrossRef]
- Soskic, S.S.; Dobutović, B.D.; Sudar, E.M.; Obradović, M.M.; Nikolić, D.M.; Djordjevic, J.D.; Radak, D.J.; Mikhailidis, D.P.; Isenović, E.R. Regulation of inducible nitric oxide synthase (iNOS) and its potential role in insulin resistance, diabetes and heart failure. Open Cardiovasc. Med. J. 2011, 5, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, T.; Tokunaga, E.; Ota, H.; Sugita, H.; Martyn, J.A.; Kaneki, M. S-nitrosylation-dependent inactivation of Akt/protein kinase B in insulin resistance. J. Biol. Chem. 2005, 280, 7511–7518. [Google Scholar] [CrossRef]
- Kaneto, H.; Xu, G.; Fujii, N.; Kim, S.; Bonner-Weir, S.; Weir, G.C. Involvement of c-Jun N-terminal kinase in oxidative stress-mediated suppression of insulin gene expression. J. Biol. Chem. 2002, 277, 30010–30018. [Google Scholar] [CrossRef]
- Manna, P.; Jain, S.K. Obesity, oxidative Stress, adipose tissue dysfunction, and the associated health risks: Causes and therapeutic strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef]
- Hajjar, D.P.; Gotto, A.M., Jr. Biological relevance of inflammation and oxidative stress in the pathogenesis of arterial diseases. Am. J. Pathol. 2013, 182, 1474–1481. [Google Scholar] [CrossRef]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Dludla, P.V.; Joubert, E.; Muller, C.J.F.; Louw, J.; Johnson, R. Hyperglycemia-induced oxidative stress and heart disease-cardioprotective effects of rooibos flavonoids and phenylpyruvic acid-2-O-beta-D-glucoside. Nutr. MeTable 2017, 14, 45. [Google Scholar] [CrossRef] [PubMed]
- Han, C.Y. Roles of reactive oxygen species on insulin resistance in adipose tissue. Diabetes Metab. J. 2016, 40, 272–279. [Google Scholar] [CrossRef] [PubMed]
- Jankovic, A.; Korac, A.; Buzadzic, B.; Otasevic, V.; Stancic, A.; Daiber, A.; Korac, B. Redox implications in adipose tissue (dys)function—A new look at old acquaintances. Redox Biol. 2015, 6, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, S.; Fujita, T.; Shimabukuro, M.; Iwaki, M.; Yamada, Y.; Nakajima, Y.; Nakayama, O.; Makishima, M.; Matsuda, M.; Shimomura, I. Increased oxidative stress in obesity and its impact on metabolic syndrome. J. Clin. Investig. 2004, 114, 1752–1761. [Google Scholar] [CrossRef]
- Den Hartigh, L.J.; Omer, M.; Goodspeed, L.; Wang, S.; Wietecha, T.; O’Brien, K.D.; Han, C.Y. Adipocyte-specific deficiency of NADPH oxidase 4 delays the onset of insulin resistance and attenuates adipose tissue inflammation in obesity. Arterioscler. Thromb. Vasc. Biol. 2017, 37, 466–475. [Google Scholar] [CrossRef] [PubMed]
- Han, C.Y.; Umemoto, T.; Omer, M.; Den Hartigh, L.J.; Chiba, T.; LeBoeuf, R.; Buller, C.L.; Sweet, I.R.; Pennathur, S.; Abel, E.D.; et al. NADPH oxidase-derived reactive oxygen species increases expression of monocyte chemotactic factor genes in cultured adipocytes. J. Biol. Chem. 2012, 287, 10379–10393. [Google Scholar] [CrossRef]
- Murphy, M.P. How mitochondria produce reactive oxygen species. Biochem. J. 2009, 417, 1–13. [Google Scholar] [CrossRef]
- Dludla, P.V.; Nkambule, N.B.; Tiano, L.; Louw, J.; Jastroch, M.; Mazibuko-Mbeje, S.E. Uncoupling proteins as a therapeutic target to protect the diabetic heart. Pharmacol. Res. 2018, 137, 11–24. [Google Scholar] [CrossRef]
- Hurrle, S.; Hsu, W.H. The etiology of oxidative stress in insulin resistance. Biomed. J. 2017, 40, 257–262. [Google Scholar] [CrossRef]
- De Marchi, E.; Baldassari, F.; Bononi, A.; Wieckowski, M.R.; Pinton, P. Oxidative stress in cardiovascular diseases and obesity: Role of p66Shc and protein kinase C. Oxid. Med. Cell. Longev. 2013, 2013, 564961. [Google Scholar] [CrossRef] [PubMed]
- Amiot, M.J.; Riva, C.; Vinet, A. Effects of dietary polyphenols on metabolic syndrome features in humans: A systematic review. Obes. Rev. 2016, 17, 573–586. [Google Scholar] [CrossRef] [PubMed]
- Gregorio, B.M.; De Souza, D.B.; de Morais Nascimento, F.A.; Pereira, L.M.; Fernandes-Santos, C. The potential role of antioxidants in metabolic syndrome. Curr. Pharm. Des. 2016, 22, 859–869. [Google Scholar] [CrossRef]
- Jack, B.U.; Malherbe, C.J.; Willenburg, E.L.; de Beer, D.; Huisamen, B.; Joubert, E.; Muller, C.J.F.; Louw, J.; Pheiffer, C. Polyphenol-enriched fractions of Cyclopia intermedia selectively affect lipogenesis and lipolysis in 3T3-L1 adipocytes. Planta Med. 2018, 84, 100–110. [Google Scholar] [CrossRef] [PubMed]
- Pollack, R.M.; Donath, M.Y.; LeRoith, D.; Leibowitz, G. Anti-inflammatory agents in the treatment of diabetes and its vascular complications. Diabetes Care 2016, 39, S244–S252. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information: PubChem CID: 370. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Gallic_acid (accessed on 9 August 2018).
- Nabavi, S.F.; Habtemariam, S.; Di Lorenzo, A.; Sureda, A.; Khanjani, S.; Nabavi, S.M.; Daglia, M. Post-stroke depression modulation and in vivo antioxidant activity of gallic acid and its synthetic derivatives in a murine model system. Nutrients 2016, 8, 248. [Google Scholar] [CrossRef] [PubMed]
- National Center for Biotechnology Information: PubChem CID: 245086. Available online: https://pubchem.ncbi.nlm.nih.gov/substance/24724290 (accessed on 9 August 2018).
- National Center for Biotechnology Information: PubChem CID: 13250. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/ethyl_gallate (accessed on 9 August 2018).
- National Center for Biotechnology Information: PubChem CID: 199472. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/Gallocatechin_gallate (accessed on 9 August 2018).
- National Center for Biotechnology Information: PubChem CID: 7428. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/7428 (accessed on 9 August 2018).
- National Center for Biotechnology Information: PubChem CID: 4947. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/propyl_gallate (accessed on 9 August 2018).
- National Center for Biotechnology Information: PubChem CID: 169167. Available online: https://pubchem.ncbi.nlm.nih.gov/compound/169167 (accessed on 9 August 2018).
- Karakaya, S. Bioavailability of phenolic compounds. Crit. Rev. Food Sci. Nutr. 2004, 44, 453–464. [Google Scholar] [CrossRef] [PubMed]
- Ma, F.W.; Deng, Q.F.; Zhou, X.; Gong, X.J.; Zhao, Y.; Chen, H.G.; Zhao, C. The tissue distribution and urinary excretion study of gallic acid and protocatechuic acid after oral administration of Polygonum aapitatum extract in rats. Molecules 2016, 21, 399. [Google Scholar] [CrossRef]
- Athukuri, B.L.; Neerati, P. Enhanced oral bioavailability of diltiazem by the influence of gallic acid and ellagic acid in male Wistar rats: Involvement of CYP3A and P-gp inhibition. Phytother. Res. 2017, 31, 1441–1448. [Google Scholar] [CrossRef]
- Shahrzad, S.; Aoyagi, K.; Winter, A.; Koyama, A.; Bitsch, I. Pharmacokinetics of gallic acid and its relative bioavailability from tea in healthy humans. J. Nutr. 2001, 131, 1207–1210. [Google Scholar] [CrossRef]
- Konishi, Y.; Hitomi, Y.; Yoshioka, E. Intestinal absorption of p-coumaric and gallic acids in rats after oral administration. J. Agric. Food Chem. 2004, 52, 2527–2532. [Google Scholar] [CrossRef] [PubMed]
- Daglia, M.; Di Lorenzo, A.; Nabavi, S.F.; Talas, Z.S.; Nabavi, S.M. Polyphenols: Well beyond the antioxidant capacity: Gallic acid and related compounds as neuroprotective agents: You are what you eat! Curr. Pharm. Biotechnol. 2014, 15, 362–372. [Google Scholar] [CrossRef] [PubMed]
- Ferruzzi, M.G.; Lobo, J.K.; Janle, E.M.; Cooper, B.; Simon, J.E.; Wu, Q.L.; Welch, C.; Ho, L.; Weaver, C.; Pasinetti, G.M. Bioavailability of gallic acid and catechins from grape seed polyphenol extract is improved by repeated dosing in rats: Implications for treatment in Alzheimer’s disease. J. Alzheimers Dis. 2009, 18, 113–124. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharyya, S.; Ahammed, S.M.; Saha, B.P.; Mukherjee, P.K. The gallic acid-phospholipid complex improved the antioxidant potential of gallic acid by enhancing its bioavailability. AAPS Pharm. Sci. Tech. 2013, 14, 1025–1033. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Song, F.; Jin, Y.C.; Zhang, W.M.; Zhang, Y.; Liu, E.J.; Zhou, D.; Bi, L.L.; Yang, Q.; Li, H.; et al. Comparative pharmacokinetics of gallic acid after oral administration of gallic acid monohydrate in normal and isoproterenol-induced myocardial infarcted rats. Front. Pharmacol. 2018, 9, 328. [Google Scholar] [CrossRef] [PubMed]
- Lutz, T.A.; Woods, S.C. Overview of animal models of obesity. Curr. Protoc. Pharmacol. 2012. [Google Scholar] [CrossRef]
- Oi, Y.; Hou, I.C.; Fujita, H.; Yazawa, K. Antiobesity effects of Chinese black tea (Pu-erh tea) extract and gallic acid. Phytother. Res. 2012, 26, 475–481. [Google Scholar] [CrossRef]
- Hsu, C.L.; Yen, G.C. Effects of flavonoids and phenolic acids on the inhibition of adipogenesis in 3T3-L1 adipocytes. J. Agric. Food Chem. 2007, 55, 8404–8410. [Google Scholar] [CrossRef]
- Hsieh, C.F.; Tsuei, Y.W.; Liu, C.W.; Kao, C.C.; Shih, L.J.; Ho, L.T.; Wu, L.Y.; Wu, C.P.; Tsai, P.H.; Chang, H.H. Green tea epigallocatechin gallate inhibits insulin stimulation of adipocyte glucose uptake via the 67-kilodalton laminin receptor and AMP-activated protein kinase pathways. Planta Med. 2010, 76, 1694–1698. [Google Scholar] [CrossRef]
- Makihara, H.; Shimada, T.; Machida, E.; Oota, M.; Nagamine, R.; Tsubata, M.; Kinoshita, K.; Takahashi, K.; Aburada, M. Preventive effect of Terminalia bellirica on obesity and metabolic disorders in spontaneously obese type 2 diabetic model mice. J. Nat. Med. 2012, 66, 459–467. [Google Scholar] [CrossRef]
- Yuda, N.; Tanaka, M.; Suzuki, M.; Asano, Y.; Ochi, H.; Iwatsuki, K. Polyphenols extracted from black tea (Camellia sinensis) residue by hot-compressed water and their inhibitory effect on pancreatic lipase in vitro. J. Food Sci. 2012, 77, H254–H261. [Google Scholar] [CrossRef] [PubMed]
- De la Garza, A.L.; Milagro, F.I.; Boque, N.; Campión, J.; Martínez, J.A. Natural inhibitors of pancreatic lipase as new players in obesity treatment. Planta Med. 2011, 77, 773–785. [Google Scholar] [CrossRef] [PubMed]
- Zengin, G.; Uysal, A.; Aktumsek, A.; Mocan, A.; Mollica, A.; Locatelli, M.; Custodio, L.; Neng, N.R.; Nogueira, J.M.F.; Aumeeruddy-Elalfi, Z.; et al. Euphorbia denticulata Lam.: A promising source of phyto-pharmaceuticals for the development of novel functional formulations. Biomed. Pharmacother. 2017, 87, 27–36. [Google Scholar] [CrossRef] [PubMed]
- Chang, P.C.; Wang, J.D.; Lee, M.M.; Chang, S.S.; Tsai, T.Y.; Chang, K.W.; Tsai, F.J.; Chen, C.Y. Lose weight with traditional chinese medicine? Potential suppression of fat mass and obesity-associated protein. J. Biomol. Struct. Dyn. 2011, 29, 471–483. [Google Scholar] [CrossRef]
- Strobel, P.; Allard, C.; Perez-Acle, T.; Calderon, R.; Aldunate, R.; Leighton, F. Myricetin, quercetin and catechin-gallate inhibit glucose uptake in isolated rat adipocytes. Biochem. J. 2005, 386, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Hsu, C.L.; Huang, S.L.; Yen, G.C. Inhibitory effect of phenolic acids on the proliferation of 3T3-L1 preadipocytes in relation to their antioxidant activity. J. Agric. Food Chem. 2006, 54, 4191–4197. [Google Scholar] [CrossRef]
- Hsu, C.L.; Yen, G.C. Effect of gallic acid on high fat diet-induced dyslipidaemia, hepatosteatosis and oxidative stress in rats. Br. J. Nutr. 2007, 98, 727–735. [Google Scholar] [CrossRef]
- Jang, A.; Srinivasan, P.; Lee, N.Y.; Song, H.P.; Lee, J.W.; Lee, M.; Jo, C. Comparison of hypolipidemic activity of synthetic gallic acid-linoleic acid ester with mixture of gallic acid and linoleic acid, gallic acid, and linoleic acid on high-fat diet induced obesity in C57BL/6 Cr Slc mice. Chem. Biol. Interact. 2008, 174, 109–117. [Google Scholar] [CrossRef]
- Booth, A.; Amen, R.J.; Scott, M.; Greenway, F.L. Oral dose-ranging developmental toxicity study of an herbal supplement (NT) and gallic acid in rats. Adv. Ther. 2010, 27, 250–255. [Google Scholar] [CrossRef]
- Punithavathi, V.R.; Stanely Mainzen Prince, P.; Kumar, M.R.; Selvakumari, C.J. Protective effects of gallic acid on hepatic lipid peroxide metabolism, glycoprotein components and lipids in streptozotocin-induced type II diabetic Wistar rats. J. Biochem. Mol. Toxicol. 2011, 25, 68–76. [Google Scholar] [CrossRef]
- Bak, E.J.; Kim, J.; Jang, S.; Woo, G.H.; Yoon, H.G.; Yoo, Y.J.; Cha, J.H. Gallic acid improves glucose tolerance and triglyceride concentration in diet-induced obesity mice. Scand. J. Clin. Lab. Investig. 2013, 73, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Ou, T.T.; Lin, M.C.; Wu, C.H.; Lin, W.L.; Wang, C.J. Gallic acid attenuates oleic acid-induced proliferation of vascular smooth muscle cell through regulation of AMPK-eNOS-FAS signaling. Curr. Med. Chem. 2013, 20, 3944–3953. [Google Scholar] [CrossRef] [PubMed]
- Doan, K.V.; Ko, C.M.; Kinyua, A.W.; Yang, D.J.; Choi, Y.H.; Oh, I.Y.; Nguyen, N.M.; Ko, A.; Choi, J.W.; Jeong, Y. Gallic acid regulates body weight and glucose homeostasis through AMPK activation. Endocrinology 2015, 156, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Chang, W.C.; Yang, H.J.; Wu, J.S.; Shen, S.C. Gallic acid alleviates hypertriglyceridemia and fat accumulation via modulating glycolysis and lipolysis pathways in perirenal adipose tissues of rats fed a high-fructose diet. Int. J. Mol. Sci. 2018, 19, 254. [Google Scholar] [CrossRef] [PubMed]
- Ong, K.C.; Khoo, H.E.; Das, N.P. Tannic acid inhibits insulin-stimulated lipogenesis in rat adipose tissue and insulin receptor function in vitro. Experientia 1995, 51, 577–584. [Google Scholar] [CrossRef]
- Ren, Y.; Himmeldirk, K.; Chen, X. Synthesis and structure-activity relationship study of antidiabetic penta-O-galloyl-D-glucopyranose and its analogues. J. Med. Chem. 2006, 49, 2829–2837. [Google Scholar] [CrossRef] [PubMed]
- Sergent, T.V.J.; Winand, J.; Beguin, P.; Schneider, Y.J. Phenolic compounds and plant extracts as potential natural anti-obesity substances. Food Chem. 2012, 135, 68–73. [Google Scholar] [CrossRef]
- Park, Y.K.; Lee, J.; Hong, V.S.; Choi, J.S.; Lee, T.Y.; Jang, B.C. Identification of KMU-3, a novel derivative of gallic acid, as an inhibitor of adipogenesis. PLoS ONE 2014, 9, e109344. [Google Scholar] [CrossRef]
- Yang, Y.; Qiao, L.; Zhang, X.; Wu, Z.; Weng, P. Effect of methylated tea catechins from Chinese oolong tea on the proliferation and differentiation of 3T3-L1 preadipocyte. Fitoterapia 2015, 104, 45–49. [Google Scholar] [CrossRef]
- Jeon, M.; Rahman, N.; Kim, Y.S. Wnt/beta-catenin signaling plays a distinct role in methyl gallate-mediated inhibition of adipogenesis. Biochem. Biophys. Res. Commun. 2016, 479, 22–27. [Google Scholar] [CrossRef]
- Amin, K.A.; Nagy, M.A. Effect of Carnitine and herbal mixture extract on obesity induced by high fat diet in rats. Diabetol. Metab. Syndr. 2009, 1, 17. [Google Scholar] [CrossRef] [PubMed]
- Hogan, S.; Canning, C.; Sun, S.; Sun, X.; Zhou, K. Effects of grape pomace antioxidant extract on oxidative stress and inflammation in diet induced obese mice. J. Agric. Food Chem. 2010, 58, 11250–11256. [Google Scholar] [CrossRef]
- Cao, Z.H.; Gu, D.H.; Lin, Q.Y.; Xu, Z.Q.; Huang, Q.C.; Rao, H.; Liu, E.W.; Jia, J.J.; Ge, C.R. Effect of pu-erh tea on body fat and lipid profiles in rats with diet-induced obesity. Phytother. Res. 2011, 25, 234–238. [Google Scholar] [CrossRef] [PubMed]
- Koh, G.Y.; McCutcheon, K.; Zhang, F.; Liu, D.; Cartwright, C.A.; Martin, R.; Yang, P.; Liu, Z. Improvement of obesity phenotype by Chinese sweet leaf tea (Rubus suavissimus) components in high-fat diet-induced obese rats. J. Agric. Food Chem. 2011, 59, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.H.; Liu, L.K.; Chuang, C.M.; Chyau, C.C.; Huang, C.N.; Wang, C.J. Mulberry water extracts possess an anti-obesity effect and ability to inhibit hepatic lipogenesis and promote lipolysis. J. Agric. Food Chem. 2011, 59, 2663–2671. [Google Scholar] [CrossRef] [PubMed]
- Monika, P.; Geetha, A. The modulating effect of Persea americana fruit extract on the level of expression of fatty acid synthase complex, lipoprotein lipase, fibroblast growth factor-21 and leptin--A biochemical study in rats subjected to experimental hyperlipidemia and obesity. Phytomedicine 2015, 22, 939–945. [Google Scholar] [CrossRef] [PubMed]
- Colantuono, A.; Ferracane, R.; Vitaglione, P. In vitro bioaccessibility and functional properties of polyphenols from pomegranate peels and pomegranate peels-enriched cookies. Food Funct. 2016, 7, 4247–4258. [Google Scholar] [CrossRef]
- de Camargo, A.C.; Regitano-d’Arce, M.A.; Biasoto, A.C.; Shahidi, F. Enzyme-assisted extraction of phenolics from winemaking by-products: Antioxidant potential and inhibition of alpha-glucosidase and lipase activities. Food Chem. 2016, 212, 395–402. [Google Scholar] [CrossRef]
- Park, B.; Lee, S.; Lee, B.; Kim, I.; Baek, N.; Lee, T.H.; Lee, S.Y.; Son, M.; Park, H. New ethanol extraction improves the anti-obesity effects of black tea. Arch. Pharm. Res. 2016, 39, 310–320. [Google Scholar] [CrossRef]
- Septembre-Malaterre, A.; Stanislas, G.; Douraguia, E.; Gonthier, M.P. Evaluation of nutritional and antioxidant properties of the tropical fruits banana, litchi, mango, papaya, passion fruit and pineapple cultivated in Reunion French Island. Food Chem. 2016, 212, 225–233. [Google Scholar] [CrossRef]
- Torabi, S.; DiMarco, N.M. Polyphenols extracted from grape powder induce lipogenesis and glucose uptake during differentiation of murine preadipocytes. Exp. Biol. Med. 2016, 241, 1776–1785. [Google Scholar] [CrossRef] [PubMed]
- Pascual-Serrano, A.; Arola-Arnal, A.; Suárez-García, S.; Bravo, F.I.; Suárez, M.; Arola, L.; Bladé, C. Grape seed proanthocyanidin supplementation reduces adipocyte size and increases adipocyte number in obese rats. Int. J. Obes. 2017, 41, 1246–1255. [Google Scholar] [CrossRef] [PubMed]
- Simao, A.A.; Marques, T.R.; Marcussi, S.; Corrêa, A.D. Aqueous extract of Psidium guajava leaves: Phenolic compounds and inhibitory potential on digestive enzymes. An. Acad. Bras. Cienc. 2017, 89, 2155–2165. [Google Scholar] [CrossRef] [PubMed]
- Ge, Q.; Chen, L.; Tang, M.; Zhang, S.; Liu, L.; Gao, L. Analysis of mulberry leaf components in the treatment of diabetes using network pharmacology. Eur. J. Pharmacol. 2018, 833, 50–62. [Google Scholar] [CrossRef]
- Sandoval-Gallegos, E.M.; Ramírez-Moreno, E.; Lucio, J.G.; Arias-Rico, J.; Cruz-Cansino, N.; Ortiz, M.I.; Cariño-Cortés, R. In vitro bioaccessibility and effect of Mangifera indica (Ataulfo) leaf extract on induced dyslipidemia. J. Med. Food 2018, 21, 47–56. [Google Scholar] [CrossRef]
- Wu, S.; Tian, L. A new flavone glucoside together with known ellagitannins and flavones with anti-diabetic and anti-obesity activities from the flowers of pomegranate (Punica granatum). Nat. Prod. Res. 2018, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Wu, C.H.; Yang, M.Y.; Chan, K.C.; Chung, P.J.; Ou, T.T.; Wang, C.J. Improvement in high-fat diet-induced obesity and body fat accumulation by a Nelumbo nucifera leaf flavonoid-rich extract in mice. J. Agric. Food Chem. 2010, 58, 7075–7081. [Google Scholar] [CrossRef]
- Batista, A.G.; Lenquiste, S.A.; Cazarin, C.B.B.; da Silva, J.K.; Luiz-Ferreira, A.; Bogusz, S., Jr.; Hantao, L.W.; de Souza, R.N.; Augusto, F.; Prado, M.A.; et al. Intake of jaboticaba peel attenuates oxidative stress in tissues and reduces circulating saturated lipids of rats with high-fat diet-induced obesity. J. Funct. Foods 2014, 6, 450–461. [Google Scholar] [CrossRef]
- Foddai, M.; Kasabri, V.; Petretto, G.L.; Azara, E.; Sias, A.; Afifi, F.U. In vitro inhibitory effects of Limonium contortirameum and L. virgatum extracts from sardinia on alpha-amylase, alpha-glucosidase and pancreatic lipase. Nat. Prod. Commun. 2014, 9, 181–184. [Google Scholar]
- Irondi, E.A.; Agboola, S.O.; Oboh, G.; Boligon, A.A. Inhibitory effect of leaves extracts of Ocimum basilicum and Ocimum gratissimum on two key enzymes involved in obesity and hypertension in vitro. J. Intercult. Ethnopharmacol. 2016, 5, 396–402. [Google Scholar] [CrossRef]
- Abeysekera, W.; Arachchige, S.P.G. Bark extracts of Ceylon cinnamon possess antilipidemic activities and bind bile acids in vitro. Evid. Based Complement. Alternat. 2017, 2017, 7347219. [Google Scholar] [CrossRef]
- Bayes, W. Gallic acid in haemoptysis. Prov. Med. Surg. J. 1852, 16, 498–499. [Google Scholar] [CrossRef] [PubMed]
- Roberts, A.T. Gallic Acid: Inhibiting Angiogenesis in Adipose Tissue. Master’s Thesis, Louisiana State University and Agricultural and Mechanical College, Barton Rouge, LA, USA, 2006. Available online: https://digitalcommons.lsu.edu/gradschool_theses/1261/ (accessed on 12 August 2018).
- Greenway, F.L.; Liu, Z.; Martin, C.K.; Kai-yuan, W.; Nofziger, J.; Rood, J.C. Safety and efficacy of NT, an herbal supplement, in treating human obesity. Int. J. Obes. 2006, 30, 1737–1741. [Google Scholar] [CrossRef] [PubMed]
- Heber, D.; Seeram, N.P.; Wyatt, H.; Henning, S.M.; Zhang, Y.; Ogden, L.G.; Dreher, M.; Hill, J.O. Safety and antioxidant activity of a pomegranate ellagitannin-enriched polyphenol dietary supplement in overweight individuals with increased waist size. J. Agric. Food Chem. 2007, 55, 10050–10054. [Google Scholar] [CrossRef] [PubMed]
- Skrzypczak-Jankun, E.; Jankun, J. Theaflavin digallate inactivates plasminogen activator inhibitor: Could tea help in Alzheimer’s disease and obesity? Int. J. Mol. Med. 2010, 26, 45–50. [Google Scholar] [PubMed]
- Kubota, K.; Sumi, S.; Tojo, H.; Sumi-Inoue, Y.; I-Chin, H.; Oi, Y.; Fujita, H.; Urata, H. Improvements of mean body mass index and body weight in preobese and overweight Japanese adults with black Chinese tea (Pu-Erh) water extract. Nutr. Res. 2011, 31, 421–428. [Google Scholar] [CrossRef]
- Hernandez, J.R.; Rizzo, J.F.; Díaz, Y.C.; Bubi, E.D.; Cabrillana, J.M.; López-Tomassetti Fernández, E.M. Effect of bismuth subgallate on the quality of life in patients undergoing Scopinaro’s biliopancreatic diversion. Surg. Obes. Relat. Dis. 2015, 11, 436–441. [Google Scholar] [CrossRef]
- Wang, C.; Shi, C.; Yang, X.; Yang, M.; Sun, H.; Wang, C. Celastrol suppresses obesity process via increasing antioxidant capacity and improving lipid metabolism. Eur. J. Pharmacol. 2014, 744, 52–58. [Google Scholar] [CrossRef]
- Wang, S.; Liang, X.; Yang, Q.; Fu, X.; Rogers, C.J.; Zhu, M. Resveratrol induces brown-like adipocyte formation in white fat through activation of AMP-activated protein kinase (AMPK) alpha1. Int. J. Obes. 2015, 39, 967–976. [Google Scholar] [CrossRef]
- Yang, C.S.; Zhang, J.; Zhang, L.; Huang, J.; Wang, Y. Mechanisms of body weight reduction and metabolic syndrome alleviation by tea. Mol. Nutr. Food Res. 2016, 60, 160–174. [Google Scholar] [CrossRef]
- Graham, H.N. Green tea composition, consumption, and polyphenol chemistry. Prev. Med. 1992, 21, 334–350. [Google Scholar] [CrossRef]
- Lin, J.K.; Lin, L.C.; Liang, Y.C.; Lin-Shiau, S.Y.; Juan, I.M. Survey of catechins, gallic acid, and methylxanthines in green, oolong, Pu-erh, and black teas. J. Agric. Food Chem. 1998, 46, 3635–3642. [Google Scholar] [CrossRef]
- Wolfram, S.; Wang, Y.; Thielecke, F. Anti-obesity effects of green tea: From bedside to bench. Mol. Nutr. Food Res. 2006, 50, 176–187. [Google Scholar] [CrossRef]
- Butler, M.S.; Robertson, A.A.; Cooper, M.A. Natural product and natural product derived drugs in clinical trials. Nat. Prod. Rep. 2014, 31, 1612–1661. [Google Scholar] [CrossRef] [PubMed]
- Paller, C.J.; Denmeade, S.R.; Carducci, M.A. Challenges of conducting clinical trials of natural products to combat cancer. Clin. Adv. Hematol. Oncol. 2016, 14, 447–455. [Google Scholar]
- Patel, O.; Muller, C.; Joubert, E.; Louw, J.; Rosenkranz, B.; Awortwe, C. Inhibitory interactions of Aspalathus linearis (rooibos) extracts and compounds, aspalathin and Z-2-(beta-d-glucopyranosyloxy)-3-phenylpropenoic acid, on cytochromes metabolizing hypoglycemic and hypolipidemic drugs. Molecules 2016, 21, 1515. [Google Scholar] [CrossRef]
- Liu, Y.; Sun, M.; Yao, H.; Liu, Y.; Gao, R. Herbal medicine for the treatment of obesity: An overview of scientific evidence from 2007 to 2017. Evid. Based Complement. Alternat. Med. 2017, 2017, 8943059. [Google Scholar] [CrossRef]
- Awortwe, C.; Makiwane, M.; Reuter, H.; Muller, C.; Louw, J.; Rosenkranz, B. Critical evaluation of causality assessment of herb-drug interactions in patients. Br. J. Clin. Pharmacol. 2018, 84, 679–693. [Google Scholar] [CrossRef]
- Wang, S.; Zhu, M.J.; Du, M. Prevention of obesity by dietary resveratrol: How strong is the evidence? Expert. Rev. Endocrinol. MeTable 2015, 10, 561–564. [Google Scholar] [CrossRef]
- Christenson, J.; Whitby, S.J.; Mellor, D.; Thomas, J.; McKune, A.; Roach, P.D. The effects of resveratrol supplementation in overweight and obese humans: A Systematic review of randomized trials. Metab. Syndr. Relat. Disord. 2016, 14, 323–333. [Google Scholar] [CrossRef]
- Fu, C.; Jiang, Y.; Guo, J.; Su, Z. Natural products with anti-obesity effects and different mechanisms of action. J. Agric. Food Chem. 2016, 9571–9585. [Google Scholar] [CrossRef] [PubMed]
- Deng, H.; Fang, Y. Anti-inflammatory gallic acid and wedelolactone are G protein-coupled receptor-35 agonists. Pharmacology 2012, 89, 211–219. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Author, Year | Experimental Model, Dose Used, and Intervention Period | Comparative/Combination Therapy | Experimental Outcome and Proposed Mechanism |
|---|---|---|---|
| Strobel et al., 2005 [103] | Adipocytes from epididymal fat pads from male Wistar rats treated with gallic acid at 0.1–100 µM | Catechin, myricetin and quercetin were used at 0.1–100 µM, together with 1 µM insulin for 30 s | All compounds inhibited glucose uptake through interfering with the function of glucose transporter (GLUT) 4 |
| Hsu et al., 2006 [104] | 3T3-L1 pre-adipocytes treated with gallic acid at 43.3 µM for 24, 48 and 72 h | Chlorogenic acid, o-coumaric acid and m-coumaric acid were used at 72.3, 48.2, and 49.2 µM, respectively, for 24, 48, and 72 h | All phenolic acids, at varying degree, improved the antioxidant status and inhibited proliferation |
| Hsu et al., 2007 [21] | 3T3-L1 pre-adipocytes treated with 0.1–250 µM gallic acid for 24, 48, and 72 h | None | Inhibited proliferation by blocking histone deacetylase activity. Further enhanced protein expression of fatty acid synthase (FAS), FAS ligand (FasL), as well as tumor protein 53 (p53) and activated caspase 3/9 |
| Hsu and Yen, 2007 [105] | High fat diet (HFD) fed male Wistar rats received 50 and 100 mg/kg body weight of gallic acid for 10 weeks | None | Reduced body weight, organ weight of the liver and adipose tissue weights. Further improved hepatic glutathione levels |
| Jang et al., 2008 [106] | HFD fed female C57BL/6 Cr Slc mice treated with gallic acid, at 1% of diet for 7 weeks | Linoleic acid and a mixture of gallic acid and linoleic acid were mixed with diet | All compounds showed hypolipidemic effects through reducing body weights and hepatic oil droplets, while improving lipid profiles |
| Booth et al., 2010 [107] | Male and female BR VAF/Plus rats given a combination of rhubarb, astragalus, red sage, ginger, and turmeric, together with gallic acid at 215, 430 and 860 mg/kg body weight for 20 days | None | Significantly reduced body weights |
| Punithavathi et al., 2011 [108] | Streptozotocin-induced diabetic male Wistar rats treated with gallic acid at 10 and 20 mg/kg body weight for 21 days | None | Reduced blood glucose and hepatic lipid peroxidation products, glycoprotein components, lipids, and the activity of β-Hydroxy β-methylglutaryl-CoA (HMG-CoA) reductase. |
| Oi et al., 2012 [95] | HFD fed female ddY mice treated with gallic acid at 15, 45 mg/kg body weight for 12 weeks | Black tea extract was used at 50, 100 mg/kg body weight for 12 weeks | Reduced body weights, as well as inhibited pancreatic lipase activity |
| Bak et al., 2013 [109] | HFD fed male C57BL/6 mice treated with gallic acid at 10 mg/kg body weight for 2 weeks | None | Reduction in adipocyte size was associated with upregulation of peroxisome proliferator-activated receptor gamma (PPAR)γ expression and activation of protein kinase B (Akt) signaling pathway |
| Ou et al., 2013 [110] | Oleic acid-induced proliferation of vascular smooth muscle cells treated with gallic acid at 10–30 µM for 48 h | None | Displayed anti-atherogenic effects, inhibited fatty acid synthase (FAS), blocked endothelial nitric oxide synthase and activated 5’ adenosine monophosphate-activated protein kinase (AMPK) |
| Chao et al., 2014 [18] | HFD fed male C57BL/6 mice treated with gallic acid at 50 and 100 mg/kg body weight for 16 weeks | None | Partially reversed metabolic disturbances, including lipid and glucose metabolism, amino acids metabolism, choline metabolism and gut-microbiota-associated metabolism |
| Doan et al., 2014 [111] | HFD fed male C57BL/6 mice treated with gallic acid at 10 mg/kg of body weight for 9 weeks | None | Induced browning of adipose tissue through activation of AMPK/Nicotinamide adenine dinucleotide (NAD)-dependent deacetylase sirtuin-1 (SIRT1)/peroxisome proliferator activated receptor gamma coactivator 1 alpha (PGC1α) pathway. Also regulated uncoupling protein 1 |
| Gandhi et al., 2014 [20] | HFD fed and streptozotocin induced diabetic male Wistar rats treated with gallic acid at 20 mg/kg body weight for 28 days | Pioglitazone was used at 10 mg/kg body weight for 28 days | Improved insulin sensitivity through translocation and activation of GLUT4 in phosphatidylinositol -3-kinase (PI3K)/p-Akt dependent pathway. Furthermore, it moderately enhanced PPARγ expression |
| Pandey et al., 2014 [17] | HFD induced male C57BL/6 mice were treated with gallic acid at 2, 4 and 8mg/kg body weight for 28 days | None | Lowered serum levels of triglycerides, and low-density lipoprotein, while increasing high density lipoprotein concentrations |
| 3T3-L1 adipocytes treated with gallic acid at 3.12, 6.25, 12.5, 25, 50 and 100 µM for 48 h | Aqueous extract Labisia pumila and pyrogallol were used at 3.12–100 µM for 48 h | Both compounds and extract showed inhibitory effect on fat droplet formation and triglyceride accumulation | |
| Makihara et al., 2016 [16] | 3T3-L1 adipocytes were treated with gallic acid at 10–30 µM during differentiation period | Troglitazone was used at 10 μM, while Terminalia bellirica hot water extract was used at 0.1, 1.0 and 10 during differentiation | The extract and gallic acid enhanced adipocyte differentiation and adiponectin secretion, partially through increasing adiponectin and fatty acid binding protein-4 levels |
| Huang et al., 2018 [112] | HFD fed male Wistar rats were treated with gallic acid at 10 or 30 mg/kg body weight for 8 weeks | Pioglitazone was used at 30 mg/kg body weight for 8 weeks | Decreased the perirenal adipose tissues and restored expression of insulin receptor and GLUT4 in the perirenal adipose tissues |
| Author, Year | Experimental Model, Dose Used, and Intervention Period | Comparative/Combination Therapy | Experimental Outcome and Proposed Mechanism |
|---|---|---|---|
| Ong et al., 1995 [113] | Adipocytes from epididymal fat pads from male Wistar rats treated with gallic acid at 1–1000 µM for various times from 20 min to 2 h | Tannic acid was used at 1–1000 µM for various times from 20 min to 2 h | Tannic acid inhibited insulin stimulated lipogenesis through promoting activation of insulin-receptor-associated tyrosine kinase phosphorylation. Whereas, gallic acid showed no effect |
| Ren et al., 2006 [114] | 3T3-L1 pre-adipocytes incubated with 6-deoxytetra-O-galloyl-α-d-glucopyranose, tetra-O-galloyl-α-d-xylopyranose and 6-chloro-6-deoxy-1,2,3,4-tetra-O-galloyl-α-d-glucopyranose at 30 µM for 15 min | None | Improved glucose uptake |
| Hsu and Yen, 2007 [96] | 3T3-L1 adipocytes were treated with gallic acid at 1–250 μM for 72 h | o-coumaric acid and rutin were used at with 1–250 μM for 72 h | o-coumaric acid and rutin demonstrated better effect in inhibiting glycerol-3-phosphate dehydrogenase activity, and the expression of peroxisome proliferator activated receptor (PPAR)γ, CAAT/enhancer-binding proteins (C/EBPR) and leptin. While also upregulating adiponectin levels |
| Hsieh et al., 2010 [97] | 3T3-L1 and C3H10T1/2 adipocytes treated with gallic acid at 5–10 μM for 2 h | Compound C, n-acetyl-l-cysteine, epigallocatechin gallate and other catechins, such as epicatechin, epigallocatechin, and epicatechin 3-gallate were used at 5–10 μM for 2 h | Epigallocatechin gallate performed better than other compounds in inhibiting insulin stimulated glucose uptake, with mechanistic involvement of 5’ adenosine monophosphate -activated protein kinase (AMPK) pathways |
| Totani et al., 2011 [22] | High fat diet fed male Wistar rats treated with gallic acid at 90 ppm in diet for 12 weeks | (z)-3-(3,4,5-trihydroxybenzoyloxy) propane-1,2-diyl dioleate (DOGGA) and octyl gallate (OG) were both used at 90 ppm in diet for 12 weeks | DOGGA showed pronounced effect than OG in reducing the body weight in rats. Gallic acid showed no effect |
| Sergent et al., 2012 [115] | In vitro bioassays testing epigallocatechin-3-gallate at 0.8 µM | Kaempferol and quercetin were effective at 13.4 and 21.5 µM, respectively | Epigallocatechin-3-gallate presented pronounced pancreatic lipase inhibitory effect than both kaempferol and quercetin |
| Park et al., 2014 [116] | 3T3-L1 adipocytes treated with gallic acid at 30, 60 and 90 µM during differentiation period | KMU-3, a derivative of gallic acid, was used at 1, 5 and 10 µM during differentiation period | KMU-3 outperformed gallic acid in suppressing lipid accumulation in cells. Mechanistically, it inhibited expressions of C/EBP-A, PPARγ, and Fas, as well as some pro-inflammatory markers |
| Yang et al., 2015 [117] | 3T3-L1 pre-adipocyte treated with epigallocatechin 3-O-(3-O-methyl) gallate and epicatechin-3-gallate at 20, 40 and 80 μg/mL for 48 h | None | Epigallocatechin 3-O-(3-O-methyl) gallate presented higher activity than epicatechin-3-gallate in inhibiting adipogenesis and proliferation |
| Jeon et al., 2016 [118] | 3T3-L1 adipocytes treated with methyl gallate at 25, 50 and 75 µM for 48 h | None | Inhibited adipogenesis through stabilizing β-catenin suppression of PPARγ expression. Further stimulated canonical Wnt/β-catenin signaling |
| Ediriweera et al., 2017 [14] | MCF-7 cells treated with gallic acid at 90 µM for 48 h | Ascorbic acid (6.5 µM), catechin (583 µM), curcumin (3.5), epigallocatechin gallate (7.5 µM), and quercetin (70 µM) for 48 h | Only quercetin, curcumin and epigallocatechin gallate showed significant protective effects against leptin-induced proliferation |
| Zengin et al., 2017 [101] | In vitro docking experiments assessing lipase inhibitory effect of gallic acid | p-OH-benzoic acid, catechin, epigallocatechin gallate, epicatechin, and rosmarinic acid | Epigallogatechin gallate and rosmarinic acid displayed best docking scores for the inhibition of α-glucosidase, α-glucosidase and lipase activities |
| Author, Year | Experimental Model, Dose Used, and Intervention Period | Comparative/Combination Therapy | Experimental Outcome and Proposed Mechanism |
|---|---|---|---|
| Ikeda et al., 2005 [15] | High fat diet fed male Sprague Dawley rats treated with tea catechins or heat-treated catechins extracts, which are rich in epigallocatechin gallate and epicatechin gallate at 1% in diet and fed for 23 days | None | Tea and the extracts markedly reduced visceral fat deposition and hepatic triglyceride levels. The activities of fatty acid synthase and malic enzyme were also decreased |
| Amin and Nagy, 2009 [119] | High fat diet fed male albino rats treated with herbal mixture extract rich in gallic acid at 790 mg/kg body weight for 4 weeks | l-carnitine was used at 250 mg/kg body weight for 4 weeks | The extract and carnitine improved disturbed lipid profile, defective antioxidant stability, and high values of insulin resistance parameters |
| Hogan et al., 2010 [120] | High fat diet fed male C57BLK/6J mice treated with Norton grape pomace extract rich in garlic acid at 2.4 g/kg of feed in order to dose each mouse at approximately 250 mg GPE/kg body weight for 12 weeks | None | The extract lowered plasma C-reactive protein levels. However, the extract did not improve oxidative stress as determined by plasma Oxygen Radical Absorbance Capacity (ORAC) assay, glutathione peroxidase, and liver lipid peroxidation |
| Cao et al., 2011 [121] | High fat diet fed male Sprague-Dawley rats treated with Pu-erh tea extract at 0.5 g, 2 g and 4 g/kg body weight for 8 weeks | None | The extract significantly lowered plasma total cholesterol, triglyceride concentrations and low-density lipoprotein-cholesterol levels. It further enhanced mRNA levels of hormone-sensitive lipase |
| Chang et al., 2011 [102] | In vitro molecular docking screening of traditional Chinese medicine, rich in gallic acid, for inhibition of fat mass and obesity-associated protein activity | (S)-tryptophan-betaxanthin, 3-methoxytyramine-betaxanthin, 4-O-methylgallic acid, syringic acid, ethacrynic acid, ferulic acid, caffeic acid, canavanine, and 3-methylthymidine | Gallic acid, together with (S)-tryptophan-betaxanthin, 3-methoxytyramine-betaxanthin and 4-O-methylgallic acid were among the leading compounds shown to inhibit fat mass and obesity-associated protein activity |
| Koh et al., 2011 [122] | High fat diet fed male Sprague Dawley rats treated with Chinese sweet leaf tea (Rubus suavissimus), rich in gallic acid, at 0.22 g/kg body weight for 9 weeks | None | Significantly reduced body weight gain and abdominal fat gain. Although food intake was not affected, blood glucose was lowered, serum triglycerides and cholesterol were significantly reduced |
| Peng et al., 2011 [123] | High fat diet fed male Syrian golden hamsters treated with mulberry water extracts, rich in gallic acid, at 0.5%, 1% and 2% of extract supplemented in diet for 12 weeks | None | The extracts lowered body weight and visceral fat, accompanied with hypolipidemic effects by reducing serum triacylglycerol, cholesterol, free fatty acid, and the low-density lipoprotein/high-density lipoprotein ratio |
| Makihara et al., 2012 [98] | Type 2 diabetic obese male TSOD mice treated with a hot water extract of Terminalia bellirica, rich in gallic acid, at 1% and 3% supplemented in diet for 8 weeks | None | The extract displayed preventive effect on obesity, insulin resistance, and hyperlipidemia. It suppressed absorption of triacylglycerol in an olive oil loading test (in vivo test) |
| In vitro pancreatic lipase activity inhibitory assay | Demonstrated inhibitory effect on pancreatic lipase activity | ||
| Yuda et al., 2012 [99] | In vitro pancreatic lipase inhibitory assay for black tea (Camellia sinensis) extracts rich in gallic acid | Theaflavin 3-O-gallate, theaflavin 3’-O-gallate, theaflavin 3,3’-O-gallate, epigallocatechin gallate, and epicatechin gallate | All extracts inhibited pancreatic lipase but extracts obtained at 100 to 140 °C showed the greatest lipase inhibition (IC50s of 0.9 to 1.3 μg/mL) |
| Esposito et al., 2015 [11] | High fat diet fed male C57BL/6J mice treated blackcurrant (Ribes nigrum L), rich in gallic acid, at 1% supplemented diet for 8 weeks | None | The extract reduced body weight gain and improved glucose metabolism |
| Monika and Geetha, 2015 [124] | High fat diet fed male Sprague Dawley rats treated with hydro-alcoholic fruit extract of avocado, rich in gallic acid, at 100 mg/kg body weight for 11 weeks | None | The extract reduced body mass index, adiposity index, total fat pad mass, blood cholesterol, triglycerides, and low-density lipoprotein. In addition, mRNA expression levels of fatty acid synthase, lipoprotein lipase, and leptin in adipose tissue was reduced |
| Colantuono et al., 2016 [125] | In vitro α-glucosidase, α-amylase and lipase inhibitory assays to assess pomegranate peels enriched cookies containing high levels of gallic acid and its derivatives | None | Showed inhibitory activity against α-glucosidase, α-amylase and α-lipase activities |
| De Camargo et al., 2016 [126] | In vitro antioxidant assays, as well as α-glucosidase and lipase inhibitory activities for phenolics from winemaking by-products rich in gallic acid | None | In addition to strong antioxidant potential, extracts showed inhibition of α-glucosidase and lipase activities |
| Park et al., 2016 [127] | High fat diet fed male C57BL/6 mice treated with an aqueous ethanol extraction of black tea, rich in gallic acid, at 100 and 300 mg/kg body weight for 8 weeks. 3T3-L1 adipocytes were exposed to 100 and 300 µg/mL during differentiation | None | Reduced body weight and body fat, improved fatty liver, regulated blood glucose, and decreased blood cholesterol. However, it did not have an effect on PPARγ protein expression |
| Septembre-Malaterre et al., 2016 [128] | 3T3-L1 pre-adipocytes treated with pineapple and mango extracts, rich in garlic acid, at 25 µM for 1 h | None | Inhibited hydrogen peroxide induced production of reactive oxygen species |
| Torabi and DiMarco, 2016 [129] | 3T3-F442A pre-adipocytes treated with grape powder extract, rich in gallic acid, at 125–500 mg GP/mL during differentiation period | None | The extract dose dependently induced adipocyte differentiation via upregulation of glucose transported (GLUT) 4, phosphatidylinositol-4,5- bisphosphate 3-kinase (PI3K) and adipogenic genes |
| Pascual-Serrano et al., 2017 [130] | High fat diet fed male Wistar rats treated with grape seed proanthocyanidin, rich in gallic acid, at 25 mg GSPE/kg body weight for 3 weeks | Gallic acid was used at 7 mg gallic acid/kg body weight for 3 weeks | Treatments did not reduce weight gain or reverse adiposity. However, the extract induced antihypertrophic and hyperplasic activities in white adipose tissue through enhancing perilipin-1 and fatty acid binding protein 4 expression and restoring adiponectin |
| Simao et al., 2017 [131] | In vitro α-amylase, α-glycosidase, lipase, and trypsin enzymes assays on aqueous extract from three cultivars of Psidium guajava L. (Pedro Sato, Paluma and Século XXI) rich in gallic acid | None | In presence of simulated gastric fluid, all cultivars showed increase in the inhibition of lipase and α-glycosidase, and decrease in inhibition of α-amylase and trypsin enzymes |
| Ge et al., 2018 [132] | The network-based pharmacological analysis was used to assess mulberry leaves rich in gallic acid | None | The extract regulated Tnf-α, PPARγ, glycogen synthase kinase-3 beta (GSK3B), insulin receptor substrate 1 (IRS1), interleukin 6 (IL-6) and other proteins involved in diabetes and obesity associated complications |
| Sandoval-Gallegos et al., 2018 [133] | High fat diet fed male Wistar rats treated with methanolic acid extract of Mangifera indica L. leaves, rich in gallic acid, at 100, 200 and 400 mg/kg for 32 days | None | In addition to increasing antioxidant capacity, the extract improved hyperlipidemic markers such as cholesterol, triglycerides, and atherogenic index |
| Wu and Tian, 2018 [134] | In vitro α-glucosidase, α-amylase and lipase inhibitory activity of flowers of pomegranate (Punica granatum) rich in gallic acid | Acarbose | The extract showed enhanced effect of suppress α-glucosidase, α-amylase, and lipase activities |
| Author, Year | Experimental Model, Dose Used, and Intervention Period | Comparative/Combination Therapy | Experimental Outcome and Proposed Mechanism |
|---|---|---|---|
| Wu et al., 2010 [135] | High fat diet fed male C57BL/6 mice treated with Nelumbo nucifera leaf extract-rich in gallic acid, supplemented at 0.5% in diet for 6 weeks | Simvastatin was used at 1 mg/kg body weight, while silymarin was used at 100 mg/kg body weight for 6 weeks | The extract performed comparable to simvastatin and silymarin in reducing body weight, body lipid accumulation, and activities of fatty acid synthase, glutamic oxaloacetic transaminase, and glutamic pyruvic transaminase |
| Batista et al., 2014 [136] | High fat diet fed male Sprague Dawley rats treated freeze-dried jaboticaba peel extract, rich in gallic acid, at 1%, 2% and 4% supplemented diet for 6 weeks | None | In addition to reducing circulating saturated free fatty acids, the extract prevented lipid peroxidation in the liver and increased its antioxidant defenses |
| Foddai et al., 2014 [137] | In vitro pancreatic triacylglycerol lipase, α-amylase and α-glucosidase inhibitory assays for Limonium spp (Plumbaginaceae) rich in epigallocatechins | Compared with acarbose, aqueous extracts of L. contortirameum and L. virgatum | All extract showed inhibitory activity on pancreatic triacylglycerol lipase, α-amylase and α-glucosidase |
| Irondi et al., 2016 [138] | In vitro pancreatic lipase and angiotensin 1-converting enzyme inhibitory assays for Ocimum basilicum extracts containing gallic acid | Ocimum gratissimum extracts | All extracts displayed high antioxidant properties. However, Ocimum basilicum displayed slightly lower activity than Ocimum gratissimum to inhibit pancreatic lipase and angiotensin 1-converting enzyme |
| Abeysekera et al., 2017 [139] | In vitro antilipidemic assays assessing potential of bark extracts of Ceylon Cinnamon rich in gallic acid | None | The extract showed inhibitory effect against HMG-CoA reductase, lipase, cholesterol esterase, and cholesterol micellization |
| Donado-Pestana et al., 2018 [19] | High fat diet fed male C57BL/6J mice treated with cagaita (Eugenia dysenterica DC.) extracts at 7 and 14 mg gallic acid equivalent (GAE)/kg body weight for 8 weeks | None | The extract protected against dyslipidemia, fasting hyperglycemia, and attenuated both hepatic gluconeogenesis and inflammation as observed by the expression of tumor necrosis factor alpha (TNF-α) and transcriptional factor NF-κB |
| Author, Year. | Experimental Model, Dose Used, And Intervention Period | Comparative/Combination Therapy | Experimental Outcome and Proposed Mechanism |
|---|---|---|---|
| Roberts, 2006 [141] | Obese human subjects receiving capsules containing 200 mg of gallic acid and 50 mg of a Chinese herbal decoction, three times a day for 24 weeks | None | Did not cause weight loss or a decrease in food intake in humans, principally due to the inability to achieve adequate serum levels |
| Greenway et al., 2006 [142] | Overweight women receiving number ten supplement (6 and mg/day), containing gallic acid, for 8 weeks | None | The supplement did not affect weight change; however had varied effect in food intake |
| Heber et al., 2007 [143] | Overweight human subjects received one or two pomegranate ellagitannin-enriched polyphenol extract capsules per day providing 710 mg (435 mg of gallic acid equivalents, GAEs) or 1420 mg (870 mg of GAEs) of extracts, respectively | None | Improved antioxidant activity through a significant reduction in thiobarbituric acid reactive substances |
| Skrzypczak-Jankun and Jankun, 2010 [144] | Plasma from human subjects treated with theaflavin digallate at 18 µM for 30 min | PAI-1 inhibitor PAI039 and epigallocatechin-3-gallate were used at 15 μM for 30 min | Inactivated plasminogen activator inhibitor type one (PAI-1) |
| Kubota et al., 2011 [145] | Pre-obese Japanese human subjects treated with water-soluble black Chinese (Pu-Erh) tea extract rich in gallic acid at 333 mg for 12 weeks | None | Exhibited significant effects in reducing the mean waist circumference, body mass index, and visceral fat values |
| Hernández et al., 2015 [146] | Obese patients undergoing biliopancreatic diversion received treatment with 2 courses of oral bismuth subgallate at 200 mg every 8 h for 12weeks, with a 4-week rest period | None | Improved the quality of life score of patients |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dludla, P.V.; Nkambule, B.B.; Jack, B.; Mkandla, Z.; Mutize, T.; Silvestri, S.; Orlando, P.; Tiano, L.; Louw, J.; Mazibuko-Mbeje, S.E. Inflammation and Oxidative Stress in an Obese State and the Protective Effects of Gallic Acid. Nutrients 2019, 11, 23. https://doi.org/10.3390/nu11010023
Dludla PV, Nkambule BB, Jack B, Mkandla Z, Mutize T, Silvestri S, Orlando P, Tiano L, Louw J, Mazibuko-Mbeje SE. Inflammation and Oxidative Stress in an Obese State and the Protective Effects of Gallic Acid. Nutrients. 2019; 11(1):23. https://doi.org/10.3390/nu11010023
Chicago/Turabian StyleDludla, Phiwayinkosi V., Bongani B. Nkambule, Babalwa Jack, Zibusiso Mkandla, Tinashe Mutize, Sonia Silvestri, Patrick Orlando, Luca Tiano, Johan Louw, and Sithandiwe E. Mazibuko-Mbeje. 2019. "Inflammation and Oxidative Stress in an Obese State and the Protective Effects of Gallic Acid" Nutrients 11, no. 1: 23. https://doi.org/10.3390/nu11010023
APA StyleDludla, P. V., Nkambule, B. B., Jack, B., Mkandla, Z., Mutize, T., Silvestri, S., Orlando, P., Tiano, L., Louw, J., & Mazibuko-Mbeje, S. E. (2019). Inflammation and Oxidative Stress in an Obese State and the Protective Effects of Gallic Acid. Nutrients, 11(1), 23. https://doi.org/10.3390/nu11010023