The Effect of Vitamin D Supplementation on Hepcidin, Iron Status, and Inflammation in Pregnant Women in the United Kingdom

 , , , ,
, , , ,
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design and Participants
2.2. Anthropometry and Characteristics
2.3. Biochemical Analysis
2.4. Statistical Analysis
3. Results
3.1. Early Pregnancy (Pre-Supplementation)
3.2. Late Pregnancy
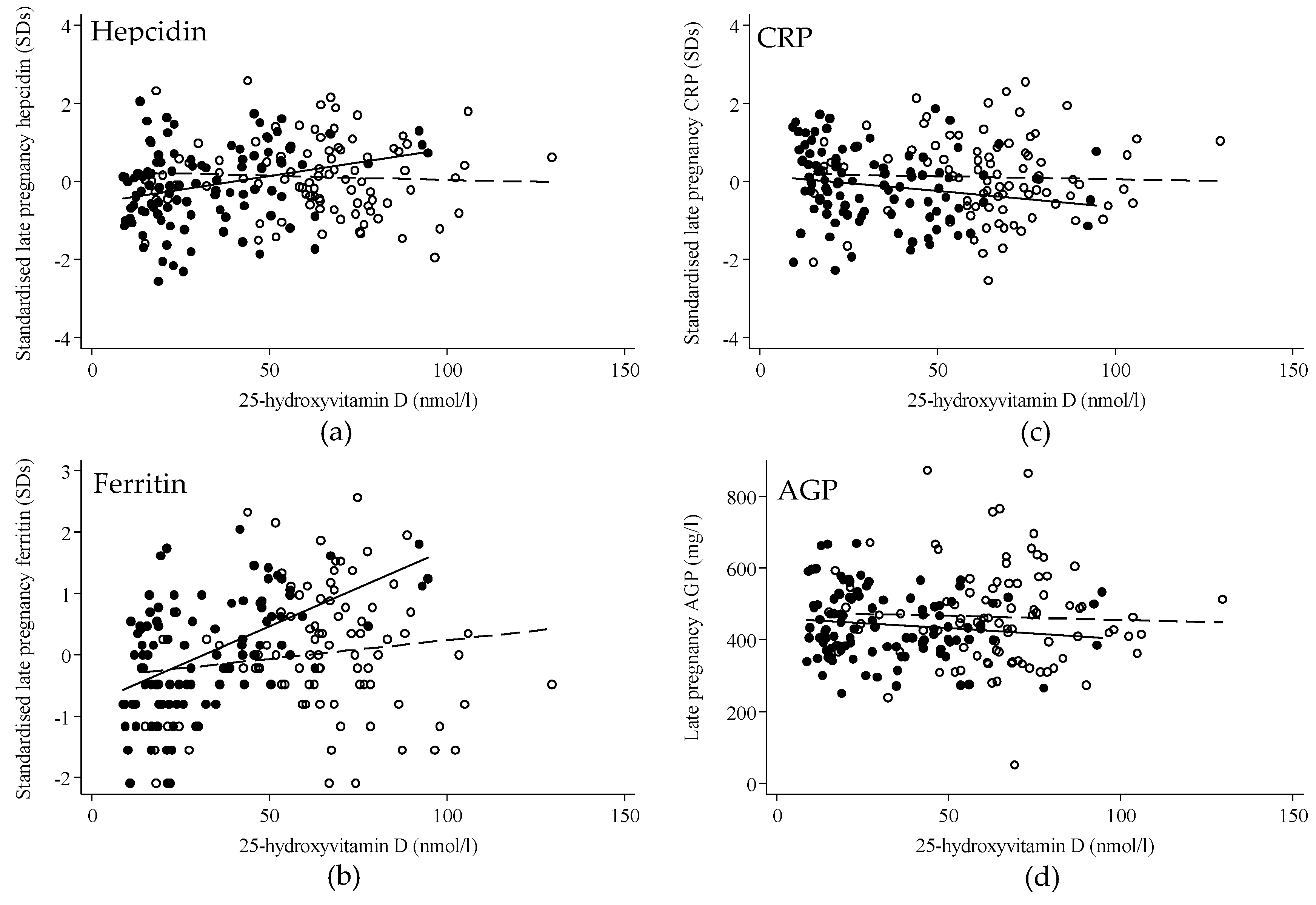
3.3. Regression Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Guillet, R.; O’Brien, K.; Thomas, C.E.; Queenan, R.A.; Cooper, E.M.; Kent, T.R.; Pressman, E.K.; Vermeylen, F.M.; Roberson, M.S.; O’Brien, K.O. Vitamin D status is inversely associated with anemia and serum erythropoietin during pregnancy. Am. J. Clin. Nutr. 2015, 102, 1088–1095. [Google Scholar]
- Nikooyeh, B.; Neyestani, T.R. Poor vitamin D status increases the risk of anemia in school children: National Food and Nutrition Surveillance. Nutrition 2018, 47, 69–74. [Google Scholar] [CrossRef] [PubMed]
- Frelut, M.-L.; Girardet, J.-P.; Bocquet, A.; Briend, A.; Chouraqui, J.-P.; Darmaun, D.; Dupont, C.; Feillet, F.; Hankard, R.; Rozé, J.-C.; et al. Impact of obesity on biomarkers of iron and vitamin D status in children and adolescents: The risk of misinterpretation. Archives de Pédiatrie 2018, 25, 3–5. [Google Scholar] [CrossRef] [PubMed]
- Michalski, E.S.; Nguyen, P.H.; Gonzalez-Casanova, I.; Nguyen, S.V.; Martorell, R.; Tangpricha, V.; Ramakrishnan, U. Serum 25-hydroxyvitamin D but not dietary vitamin D intake is associated with hemoglobin in women of reproductive age in rural northern Vietnam. J. Clin. Transl. Endocrinol. 2017, 8, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Malczewska-Lenczowska, J.; Sitkowski, D.; Surała, O.; Orysiak, J.; Szczepańska, B.; Witek, K. The Association between Iron and Vitamin D Status in Female Elite Athletes. Nutrients 2018, 10, 167. [Google Scholar] [CrossRef] [PubMed]
- Ganz, T.; Nemeth, E. Hepcidin and iron homeostasis. Biochimica et Biophysica Acta (BBA)-Mol. Cell Res. 2012, 1823, 1434–1443. [Google Scholar] [CrossRef] [PubMed]
- Zughaier, S.M.; Alvarez, J.A.; Sloan, J.H.; Konrad, R.J.; Tangpricha, V. The role of vitamin D in regulating the iron-hepcidin-ferroportin axis in monocytes. J. Clin. Transl. Endocrinol. 2014, 1, e19–e25. [Google Scholar] [CrossRef]
- Bacchetta, J.; Zaritsky, J.J.; Sea, J.L.; Chun, R.F.; Lisse, T.S.; Zavala, K.; Nayak, A.; Wesseling-Perry, K.; Westerman, M.; Hollis, B.W.; et al. Suppression of Iron-Regulatory Hepcidin by Vitamin D. J. Am. Soc. Nephrol. 2013, 25, 564–572. [Google Scholar] [CrossRef]
- Smith, E.M.; Alvarez, J.A.; Kearns, M.D.; Hao, L.; Sloan, J.H.; Konrad, R.J.; Ziegler, T.R.; Zughaier, S.M.; Tangpricha, V. High-dose vitamin D 3 reduces circulating hepcidin concentrations: A pilot, randomized, double-blind, placebo-controlled trial in healthy adults. Clin. Nutr. 2017, 36, 980–985. [Google Scholar] [CrossRef]
- Laird, E.; McNulty, H.; Ward, M.; Hoey, L.; McSorley, E.; Wallace, J.M.W.; Carson, E.; Molloy, A.M.; Healy, M.; Casey, M.C.; et al. Vitamin D Deficiency Is Associated with Inflammation in Older Irish Adults. J. Clin. Endocrinol. Metab. 2014, 99, 1807–1815. [Google Scholar] [CrossRef]
- Calton, E.K.; Keane, K.N.; Newsholme, P.; Zhao, Y.; Soares, M.J. The impact of cholecalciferol supplementation on the systemic inflammatory profile: A systematic review and meta-analysis of high-quality randomized controlled trials. Eur. J. Clin. Nutr. 2017, 71, 931–943. [Google Scholar] [CrossRef] [PubMed]
- Vanherwegen, A.-S.; Gysemans, C.; Mathieu, C. Vitamin D endocrinology on the cross-road between immunity and metabolism. Mol. Cell. Endocrinol. 2017, 453, 52–67. [Google Scholar] [CrossRef] [PubMed]
- WHO. The Global Prevalence of Anaemia 2011; WHO: Geneva, Switzerland, 2015. [Google Scholar]
- Stevens, G.A.; Finucane, M.M.; De-Regil, L.M.; Paciorek, C.J.; Flaxman, S.R.; Branca, F.; Peña-Rosas, J.P.; Bhutta, Z.A.; Ezzati, M. Global, regional, and national trends in haemoglobin concentration and prevalence of total and severe anaemia in children and pregnant and non-pregnant women for 1995–2011: A systematic analysis of population-representative data. Lancet Glob. Health 2013, 1, e16–e25. [Google Scholar] [CrossRef]
- Allen, L.H. Anemia and iron deficiency: Effects on pregnancy outcome. Am. J. Clin. Nutr. 2000, 71, 1280S–1284S. [Google Scholar] [CrossRef] [PubMed]
- Haider, B.A.; Olofin, I.; Wang, M.; Spiegelman, D.; Ezzati, M.; Fawzi, W.W. Anaemia, prenatal iron use, and risk of adverse pregnancy outcomes: Systematic review and meta-analysis. BMJ 2013, 346, f3443. [Google Scholar] [CrossRef] [PubMed]
- Bah, A.; Pasricha, S.-R.; Jallow, M.W.; A Sise, E.; Wegmuller, R.; E Armitage, A.; Drakesmith, H.; Moore, S.E.; Prentice, A.M. Serum Hepcidin Concentrations Decline during Pregnancy and May Identify Iron Deficiency: Analysis of a Longitudinal Pregnancy Cohort in The Gambia. J. Nutr. 2017, 147, 1131–1137. [Google Scholar] [CrossRef] [PubMed]
- Watts, D.H.; Krohn, M.A.; Wener, M.H.; Eschenbach, D.A. C-Reactive Protein in Normal Pregnancy. Obstet. Gynecol. 1991, 77, 176–180. [Google Scholar] [CrossRef] [PubMed]
- Koenig, M.D.; Tussing-Humphreys, L.; Day, J.; Cadwell, B.; Nemeth, E. Hepcidin and Iron Homeostasis during Pregnancy. Nutrients 2014, 6, 3062–3083. [Google Scholar] [CrossRef]
- Harvey, N.C.; Javaid, K.; Bishop, N.; Kennedy, S.; Papageorghiou, A.T.; Fraser, R.; Gandhi, S.V.; Schoenmakers, I.; Prentice, A.; Cooper, C.; et al. MAVIDOS Maternal Vitamin D Osteoporosis Study: Study protocol for a randomized controlled trial. The MAVIDOS Study Group. Trials 2012, 13, 13. [Google Scholar] [CrossRef]
- Cooper, C.; Harvey, N.C.; Bishop, N.J.; Kennedy, S.; Papageorghiou, A.T.; Schoenmakers, I.; Fraser, R.; Gandhi, S.V.; Carr, A.; D’Angelo, S.; et al. Maternal gestational vitamin D supplementation and offspring bone health (MAVIDOS): A multicentre, double-blind, randomised placebo-controlled trial. Lancet Diabetes Endocrinol. 2016, 4, 393–402. [Google Scholar] [CrossRef]
- Webb, A.; Kazantzidis, A.; Kift, R.; Farrar, M.; Wilkinson, J.; Rhodes, L. Colour Counts: Sunlight and Skin Type as Drivers of Vitamin D Deficiency at UK Latitudes. Nutrients 2018, 10, 457. [Google Scholar] [CrossRef] [PubMed]
- Armitage, P.; Berry, G.; Matthews, J.N.S. Statistical Methods in Medical Research; Wiley: Hoboken, NJ, USA, 2002. [Google Scholar]
- Namaste, S.M.; Rohner, F.; Huang, J.; Bhushan, N.L.; Flores-Ayala, R.; Kupka, R.; Mei, Z.; Rawat, R.; Williams, A.M.; Raiten, D.J.; et al. Adjusting ferritin concentrations for inflammation: Biomarkers Reflecting Inflammation and Nutritional Determinants of Anemia (BRINDA) project. Am. J. Clin. Nutr. 2017, 106, 359–371. [Google Scholar]
- WHO. Serum Ferritin Concentrations for the Assessment of Iron Status and Iron Deficiency in Populations. Vitamin and Mineral Nutrition Information System 2011 [cited 2017 24/10/2017]. Available online: http://www.who.int/vmnis/indicators/serum_ferritin/en (accessed on 16 January 2019).
- Roth, D.E.; Abrams, S.A.; Aloia, J.; Bergeron, G.; Bourassa, M.W.; Brown, K.H.; Calvo, M.S.; Cashman, K.D.; Combs, G.; De-Regil, L.M.; et al. Global prevalence and disease burden of vitamin D deficiency: A roadmap for action in low- and middle-income countries. Ann. N. Y. Acad. Sci. 2018, 1430, 44–79. [Google Scholar] [CrossRef] [PubMed]
- Mwangi, M.N.; Roth, J.M.; Smit, M.R.; Trijsburg, L.; Mwangi, A.M.; Demir, A.Y.; Wielders, J.P.; Mens, P.F.; Verweij, J.J.; Cox, S.E.; et al. Effect of Daily Antenatal Iron Supplementation on Plasmodium Infection in Kenyan Women: A Randomized Clinical Trial. JAMA 2015, 314, 1009–1020. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; O’Brien, K. Pregnancy and iron homeostasis: An update. Nutr. Rev. 2013, 71, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.S.; Assar, S.; Prentice, A.; Schoenmakers, I. Vitamin D expenditure is not altered in pregnancy and lactation despite changes in vitamin D metabolite concentrations. Sci. Rep. 2016, 6, 26795. [Google Scholar] [CrossRef] [PubMed]
- Schoenmakers, I.; Jones, K.S. Chapter 37—Pharmacology and Pharmacokinetics, in Vitamin D, 4th ed.; Feldman, D., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 635–661. [Google Scholar]
- Byg, K.-E.; Milman, N.; Hansen, S.; Agger, A.O. Serum Ferritin is a Reliable, Non-invasive Test for Iron Status in Pregnancy: Comparison of Ferritin with Other Iron Status Markers in a Longitudinal Study on Healthy Pregnant Women; Erythropoiesis. Hematology 2000, 5, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Daru, J.; Colman, K.; Stanworth, S.J.; De La Salle, B.; Wood, E.M.; Pasricha, S.-R. Serum ferritin as an indicator of iron status: What do we need to know? Am. J. Clin. Nutr. 2017, 106, 1634S–1639S. [Google Scholar] [CrossRef]


{kind=link}
| Measures in Early Pregnancy | All | Placebo | Vitamin D3 |
|---|---|---|---|
| n = (195) | n = (102) | n = (93) | |
| White ethnicity, n (%) | 182 (94%) | 95 (93%) | 87 (95%) |
| Previous children, median (IQR) | 1 (0, 1) | 1 (0, 1) | 1 (0, 1) |
| Nulliparous, n (%) | 91 (47%) | 49 (49%) | 42 (46%) |
| Age (years), mean (SD) | 30.4 (5.2) | 30.8 (5.2) | 29.9 (5.2) |
| Gestation (weeks), mean (SD) | 15.7 (1.0) | 15.8 (1.0) | 15.6 (1.0) |
| Weight (kg), median (IQR) | 69.6 (61.5, 80.0) | 70.5 (62.4, 79.6) | 68.0 (60.7, 81.0) |
| Height (m), mean (SD) | 1.66 (0.07) | 1.66 (0.07) | 1.66 (0.07) |
| BMI (kg/m2), median (IQR) | 24.7 (22.1, 28.7) | 24.9 (22.0, 28.8) | 24.6 (22.1, 28.6) |
| Supplement use, n (%) | 178 (92%) | 92 (90%) | 86 (93%) |
| Iron supplement use, n (% of those using supplements) | 106 (60%) | 49 (53%) | 57 (66%) |
| CRP (mg/L), median (IQR) | 5.4 (3.1, 8.3) | 4.2 (2.9, 7.8) | 6.2 (3.6, 11.5) |
| CRP > 5 mg/L, n (%) | 103 (53%) | 48 (47%) | 55 (60%) |
| AGP (mg/L), mean (SD) | 506 (115) | 500 (109) | 512 (122) |
| AGP > 500 mg/L, n (%) | 96 (49%) | 50 (49%) | 46 (49%) |
| Hepcidin (μg/L), median (IQR) | 7.3 (3.0, 16.7) | 7.4 (2.5, 16.5) | 6.9 (3.3, 16.8) |
| Ferritin (μg/L), median (IQR) | 38 (25, 60) | 43 (26, 62) | 35 (25, 55) |
| Ferritin < 15 μg/L, n (%) | 19 (9.7%) | 11 (10.8%) | 8 (8.6%) |
| 25-hydroxyvitamin D (nmol/L), mean (SD) | 44.1 (16.0) | 42.5 (15.8) | 45.7 (16.2) |
| 25-hydroxyvitamin D < 25 nmol/L, n (%) | 23 (12%) | 15 (15%) | 8 (9%) |
| 25-hydroxyvitamin D < 50 nmol/L, n (%) | 128 (66%) | 70 (69%) | 58 (62%) |
| Measures in Late Pregnancy | All | Placebo | Vitamin D3 | p-Value |
|---|---|---|---|---|
| n = (195) | n = (102) | n = (93) | ||
| Age (years), mean (SD) | 30.6 (5.3) | 31.2 (5.2) d | 30.0 (5.3) d | 0.13 |
| Gestation (weeks), mean (SD) | 34.7 (0.8) | 34.6 (0.6) d | 34.8 (0.9) d | 0.06 |
| Weight (kg), median (IQR) | 78.8 (71.1, 90.4) | 78.5 (73.2, 90.0) d | 79.2 (69.9, 92.8) d | 0.69 |
| BMI (kg/m2), median (IQR) | 28.4 (25.5, 32.9) | 28.6 (25.5, 32.5) d | 28.3 (25.4, 32.9) d | 0.71 |
| Supplement use, n (%) | 119 (62%) | 61 (61%) d | 58 (63%) d | 0.77 |
| Iron supplement use, n (% of those using supplements) | 111 (93%) | 54 (89%) a | 57 (98%) b | 0.03 |
| CRP (mg/L), median (IQR) | 3.9 (2.5, 7.3) | 3.8 (2.3, 6.4) c | 4.1 (2.6, 8.1) c | 0.16 |
| CRP > 5 mg/L, n (%) | 69 (38%) | 31 (32%) b | 38 (43%) b | 0.13 |
| AGP (mg/L), mean (SD) | 452 (115) | 442 (93) d | 463 (135) b | 0.20 |
| AGP > 500 mg/L, n (%) | 57 (29%) | 28 (27%) c | 29 (32%) b | 0.53 |
| Hepcidin (μg/L), median (IQR) | 0.97 (0.79, 1.99) | 0.93 (0.74, 1.57) d | 0.99 (0.84, 2.30) d | 0.19 |
| Ferritin (μg/L), median (IQR) | 10 (7, 18) | 10 (7, 17) d | 10 (8, 18) d | 0.94 |
| Ferritin < 15 μg/L, n (%) | 129 (67%) | 65 (64%) d | 64 (70%) d | 0.44 |
| 25-hydroxyvitamin D (nmol/L), median (IQR) | 47.1 (21.4, 66.8) | 25.2 (16.9, 45.8) d | 64.6 (52.0, 75.7) d | 0.0001 |
| 25-hydroxyvitamin D < 25 nmol/L, n (%) | 61 (31%) | 51 (50%) d | 10 (11%) | 0.001 |
| 25-hydroxyvitamin D < 50 nmol/L, n (%) | 107 (55%) | 85 (83%) b | 22 (24%) d | 0.001 |
| Change in Characteristic | All | Placebo | Vitamin D3 | p-Value |
|---|---|---|---|---|
| n = (195) | n = (102) | n = (93) | ||
| Maternal pregnancy weight (kg), mean (SD) | 9.8 (3.6) | 9.7 (3.5) | 9.8 (3.8) | 0.65 |
| Maternal pregnancy BMI (kg/m2), mean (SD) | 3.6 (1.3) | 3.6 (1.2) | 3.5 (1.4) | 0.44 |
| CRP (mg/L), mean (SD) | −1.6 (10.3) | −1.8 (9.4) | −1.4 (11.2) | 0.12 |
| AGP (mg/L), mean (SD) | −54 (126) | −58 (101) | −50 (149) | 0.76 |
| Hepcidin (μg/L), median (IQR) | −5.1 (−14.1, −1.3) | −5.6 (−14.9, −1.1) | −4.3 (−11.0, −1.6) | 0.10 |
| Ferritin (μg/L), mean (SD) | −31.9 (58.5) | −36.5 (41.2) | −26.9 (72.9) | 0.67 |
| 25-hydroxyvitamin D (nmol/L), mean (SD) | 2.6 (27.1) | −10.7 (17.9) | 17.1 (28.0) | <0.0001 |
| All Women (n = 195) | Beta (95% CI) | p-Value | n | Interaction p-Value * |
|---|---|---|---|---|
| Early pregnancy: hepcidin (SDs) | 0.02 (−0.07, 0.11) | 0.67 | 195 | 0.99 |
| Late pregnancy: hepcidin (SDs) | 0.06 (0.01, 0.11) | 0.03 | 195 | 0.02 |
| Early pregnancy: ferritin (SDs) | 0.06 (−0.02, 0.15) | 0.16 | 195 | 0.85 |
| Late pregnancy: ferritin (SDs) | 0.09 (0.04, 0.15) | <0.001 | 193 | 0.003 |
| Early pregnancy: CRP (SDs) | −0.10 (−0.19, −0.02) | 0.02 | 193 | 0.23 |
| Late pregnancy: CRP (SDs) | −0.01 (−0.06, 0.05) | 0.85 | 184 | 0.32 |
| Early pregnancy: AGP (mg/L) | −5.3 (−15.4, 4.9) | 0.31 | 195 | 0.06 |
| Late pregnancy: AGP (mg/L) | −0.1 (6.4, 6.2) | 0.97 | 194 | 0.66 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braithwaite, V.S.; Crozier, S.R.; D’Angelo, S.; Prentice, A.; Cooper, C.; Harvey, N.C.; Jones, K.S.; the MAVIDOS Trial Group. The Effect of Vitamin D Supplementation on Hepcidin, Iron Status, and Inflammation in Pregnant Women in the United Kingdom. Nutrients 2019, 11, 190. https://doi.org/10.3390/nu11010190
Braithwaite VS, Crozier SR, D’Angelo S, Prentice A, Cooper C, Harvey NC, Jones KS, the MAVIDOS Trial Group. The Effect of Vitamin D Supplementation on Hepcidin, Iron Status, and Inflammation in Pregnant Women in the United Kingdom. Nutrients. 2019; 11(1):190. https://doi.org/10.3390/nu11010190
Chicago/Turabian StyleBraithwaite, Vickie S., Sarah R. Crozier, Stefania D’Angelo, Ann Prentice, Cyrus Cooper, Nicholas C. Harvey, Kerry S. Jones, and the MAVIDOS Trial Group. 2019. "The Effect of Vitamin D Supplementation on Hepcidin, Iron Status, and Inflammation in Pregnant Women in the United Kingdom" Nutrients 11, no. 1: 190. https://doi.org/10.3390/nu11010190
APA StyleBraithwaite, V. S., Crozier, S. R., D’Angelo, S., Prentice, A., Cooper, C., Harvey, N. C., Jones, K. S., & the MAVIDOS Trial Group. (2019). The Effect of Vitamin D Supplementation on Hepcidin, Iron Status, and Inflammation in Pregnant Women in the United Kingdom. Nutrients, 11(1), 190. https://doi.org/10.3390/nu11010190