Ultraviolet Irradiation of Skin Alters the Faecal Microbiome Independently of Vitamin D in Mice

 ,
,
Abstract
1. Introduction
2. Materials and Methods
2.1. Mice and Diets
2.2. UV Radiation
2.3. Faecal Microbiota Analysis
2.4. Real-Time PCR
2.5. Colitis Model
2.6. Murine Colonoscopy
2.7. Histological Assessment of Colitis
2.8. Measurement of Serum Metabolites
2.9. Statistical Analyses
3. Results
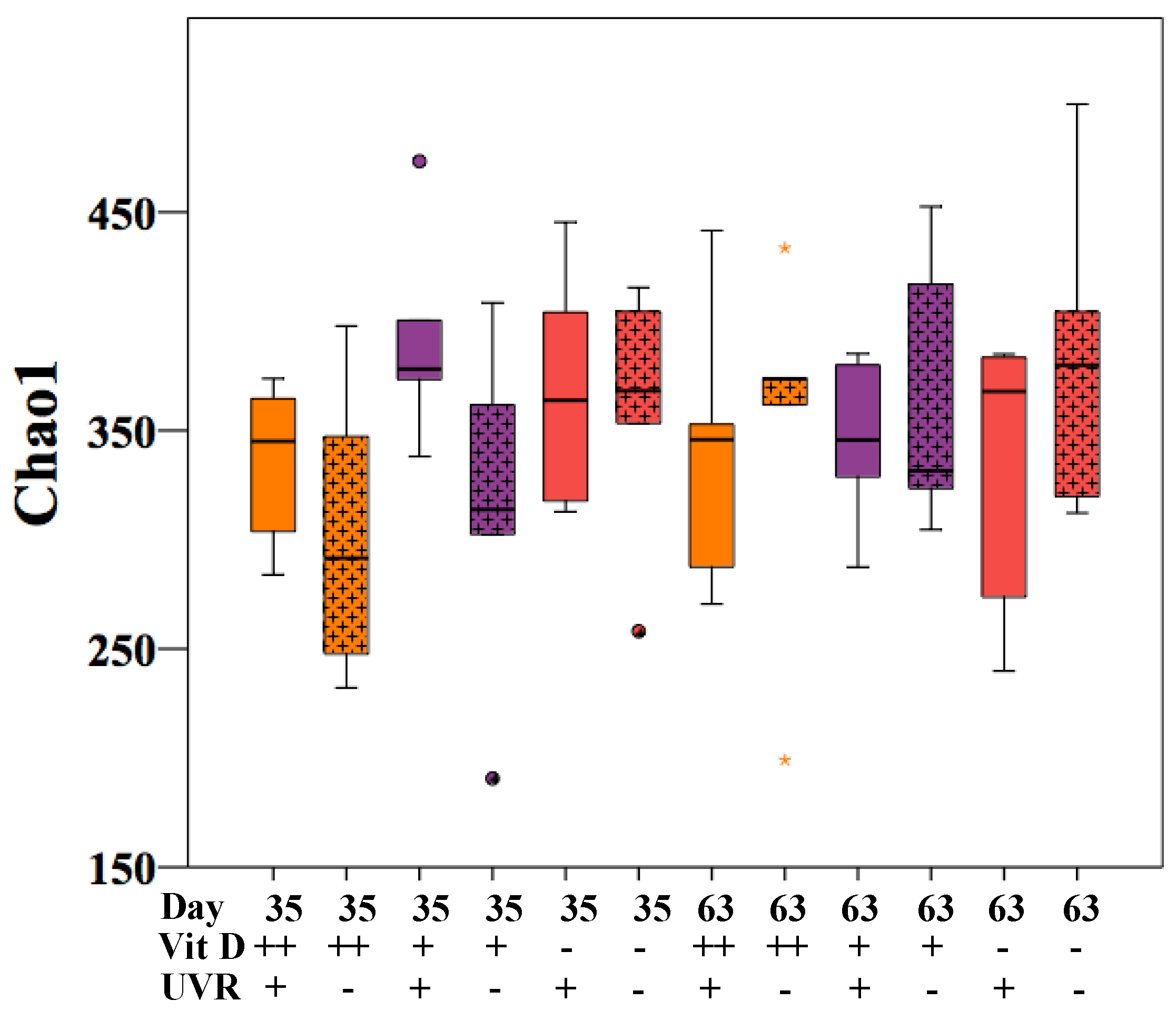
3.1. Microbiome Changes in UV-Irradiated Mice
3.2. Effect of UV Irradiation of Skin on Serum Vitamin D and Cytokine Levels
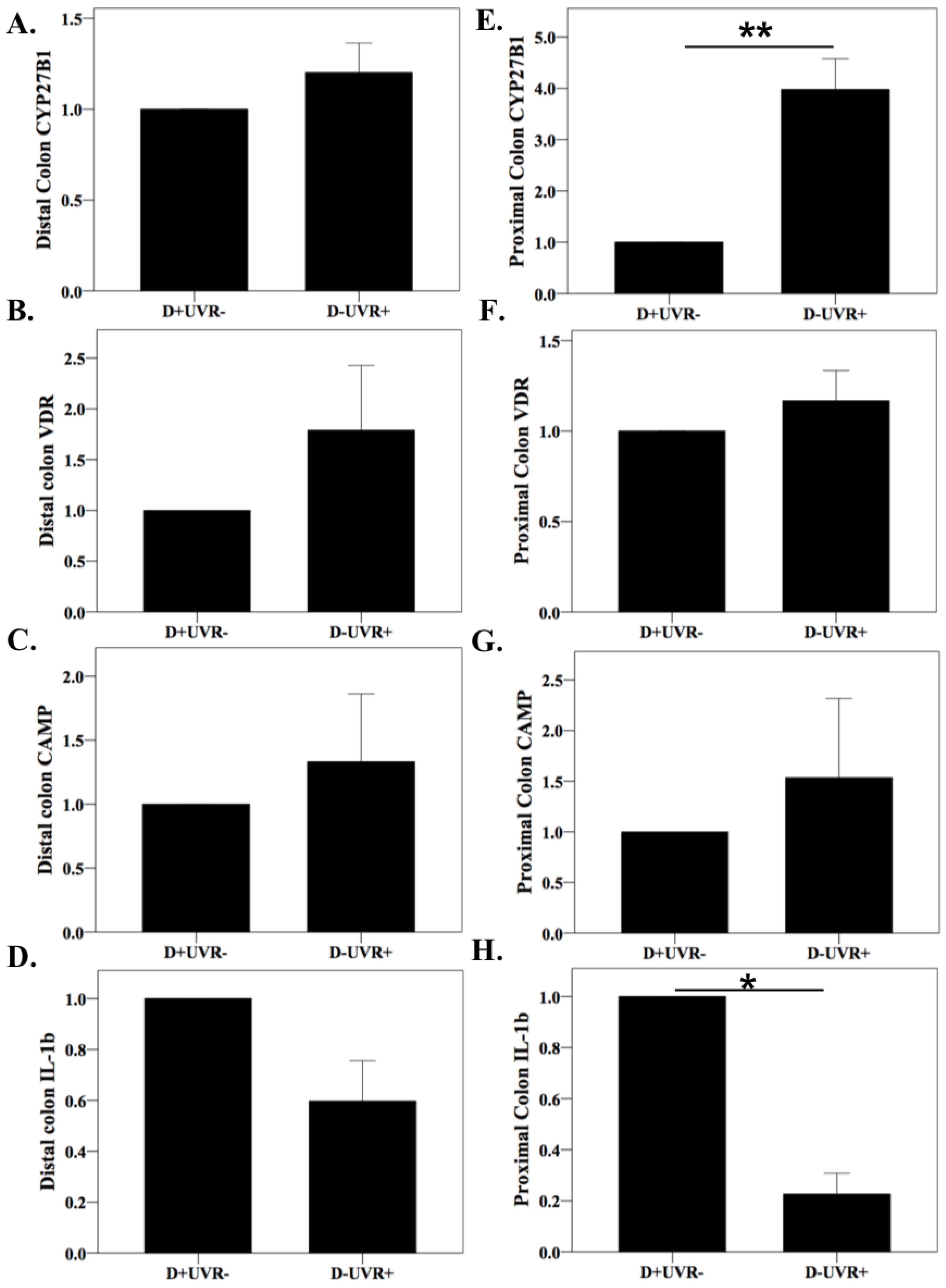
3.3. Effect of UV Irradiation on Colonic and Kidney Vitamin D Pathway Gene Expression
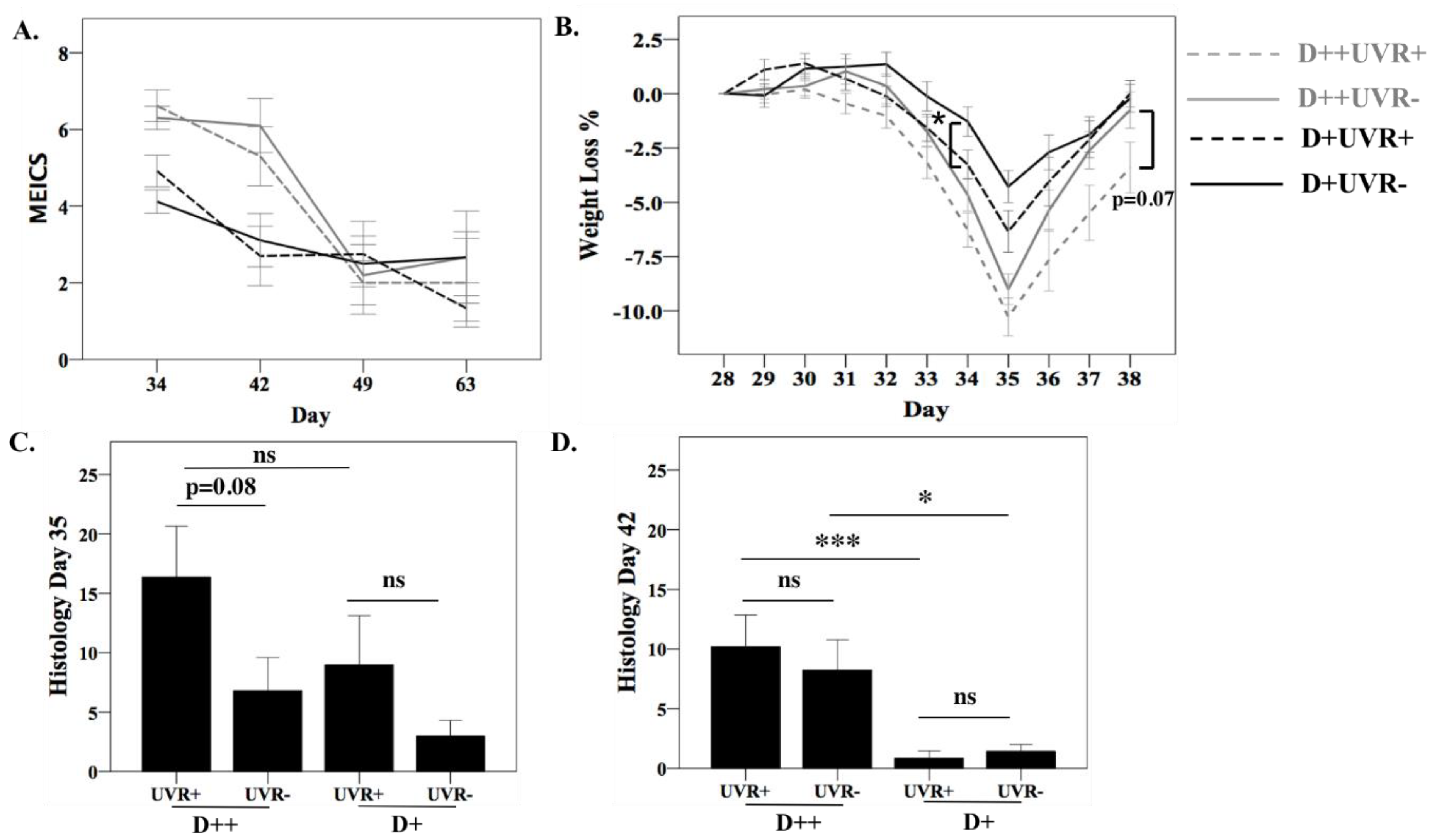
3.4. The Effect of UV Irradiation of Skin on DSS Colitis
3.5. Microbiome in Faecal Samples from DSS Mice
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Shivananda, S.; Lennard-Jones, J.; Logan, R.; Fear, N.; Price, A.; Carpenter, L.; Blankenstein, M.V. Incidence of inflammatory bowel disease across europe: Is there a difference between north and south? Results of the european collaborative study on inflammatory bowel disease (EC–IBD). Gut 1996, 39, 690–697. [Google Scholar] [CrossRef] [PubMed]
- Gower-Rousseau, C.; Salomez, J.L.; Dupas, J.L.; Marti, R.; Nuttens, M.C.; Votte, A.; Lemahieu, M.; Lemaire, B.; Colombel, J.F.; Cortot, A. Incidence of inflammatory bowel disease in northern france (1988–1990). Gut 1994, 35, 1433–1438. [Google Scholar] [CrossRef] [PubMed]
- Sonnenberg, A. Demographic characteristics of hospitalized IBD patients. Dig. Dis. Sci. 2009, 54, 2449–2455. [Google Scholar] [CrossRef] [PubMed]
- Nerich, V.; Monnet, E.; Etienne, A.; Louafi, S.; Ramée, C.; Rican, S.; Weill, A.; Vallier, N.; Vanbockstael, V.; Auleley, G.-R.; et al. Geographical variations of inflammatory bowel disease in france: A study based on national health insurance data. Inflamm. Bowel Dis. 2006, 12, 218–226. [Google Scholar] [CrossRef] [PubMed]
- Nerich, V.; Jantchou, P.; Boutron-Ruault, M.-C.; Monnet, E.; Weill, A.; Vanbockstael, V.; Auleley, G.-R.; Balaire, C.; Dubost, P.; Rican, S.; et al. Low exposure to sunlight is a risk factor for Crohn’s disease. Aliment. Pharmacol. Ther. 2011, 33, 940–945. [Google Scholar] [CrossRef] [PubMed]
- Assa, A.; Vong, L.; Pinnell, L.J.; Avitzur, N.; Johnson-Henry, K.C.; Sherman, P.M. Vitamin d deficiency promotes epithelial barrier dysfunction and intestinal inflammation. J. Infect. Dis. 2014, 210, 1296–1305. [Google Scholar] [CrossRef] [PubMed]
- Ghaly, S.; Kaakoush, N.O.; Lloyd, F.; McGonigle, T.; Mok, D.; Baird, A.; Klopcic, B.; Gordon, L.; Gorman, S.; Forest, C.; et al. High dose vitamin D supplementation alters faecal microbiome and predisposes mice to more severe colitis. Sci. Rep. 2018, 8, 11511. [Google Scholar] [PubMed]
- Hart, P.H.; Gorman, S.; Finlay-Jones, J.J. Modulation of the immune system by UV radiation: More than just the effects of vitamin d? Nat. Rev. Immunol. 2011, 11, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Ullrich, S.E.; Byrne, S.N. The immunologic revolution: Photoimmunology. J. Investig. Dermatol. 2012, 132, 896–905. [Google Scholar] [CrossRef] [PubMed]
- Wirtz, S.; Neurath, M.F. Mouse models of inflammatory bowel disease. Adv. Drug Deliv. Rev. 2007, 59, 1073–1083. [Google Scholar] [CrossRef] [PubMed]
- Gorman, S.; Scott, N.M.; Tan, D.H.; Weeden, C.E.; Tuckey, R.C.; Bisley, J.L.; Grimbaldeston, M.A.; Hart, P.H. Acute erythemal ultraviolet radiation causes systemic immunosuppression in the absence of increased 25-hydroxyvitamin D3 levels in male mice. PLoS ONE 2012, 7, e46006. [Google Scholar] [CrossRef] [PubMed]
- Geldenhuys, S.; Hart, P.H.; Endersby, R.; Jacoby, P.; Feelisch, M.; Weller, R.B.; Matthews, V.; Gorman, S. Ultraviolet radiation suppresses obesity and symptoms of metabolic syndrome independently of vitamin D in mice fed a high-fat diet. Diabetes 2014, 63, 3759–3769. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Kobert, K.; Flouri, T.; Stamatakis, A. Pear: A fast and accurate illumina paired-end read merger. Bioinformatics 2014, 30, 614–620. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.; Jee, S.H.; Kim, M.; Yoo, H.J.; Kang, M.; Kim, J.; Lee, J.H. Serum vitamin A-related metabolite levels are associated with incidence of type 2 diabetes. Dia. Metabol. 2017, 43, 287–291. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Pena, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. Search and clustering orders of magnitude faster than blast. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C.; Haas, B.J.; Clemente, J.C.; Quince, C.; Knight, R. Uchime improves sensitivity and speed of chimera detection. Bioinformatics 2011, 27, 2194–2200. [Google Scholar] [CrossRef] [PubMed]
- DeSantis, T.Z.; Hugenholtz, P.; Larsen, N.; Rojas, M.; Brodie, E.L.; Keller, K.; Huber, T.; Dalevi, D.; Hu, P.; Andersen, G.L. Greengenes, a chimera-checked 16s rRNA gene database and workbench compatible with ARB. Appl. Environ. Microbiol. 2006, 72, 5069–5072. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome. Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Gorman, S.; Judge, M.A.; Hart, P.H. Topical 1,25-dihydroxyvitamin D3 subverts the priming ability of draining lymph node dendritic cells. Immunology 2010, 131, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Ng, Y.-L.; Klopcic, B.; Lloyd, F.; Forrest, C.; Greene, W.; Lawrance, I.C. Secreted protein acidic and rich in cysteine (SPARC) exacerbates colonic inflammatory symptoms in dextran sodium sulphate-induced murine colitis. PLoS ONE 2013, 8, e77575. [Google Scholar] [CrossRef] [PubMed]
- Becker, C.; Fantini, M.C.; Wirtz, S.; Nikolaev, A.; Kiesslich, R.; Lehr, H.A.; Galle, P.R.; Neurath, M.F. In vivo imaging of colitis and colon cancer development in mice using high resolution chromoendoscopy. Gut 2005, 54, 950–954. [Google Scholar] [CrossRef] [PubMed]
- Dieleman, L.A.; Palmen, M.J.; Akol, H.; Bloemena, E.; Peña, A.S.; Meuwissen, S.G.; Van Rees, E.P. Chronic experimental colitis induced by dextran sulphate sodium (DSS) is characterized by th1 and th2 cytokines. Clin. Exp. Immunol. 1998, 114, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Clarke, M.W.; Tuckey, R.C.; Gorman, S.; Holt, B.; Hart, P.H. Optimized 25-hydroxyvitamin d analysis using liquid-liquid extraction with 2D separation with LC/MS/MS detection, provides superior precision compared to conventional assays. Metabolomics 2013, 9, 1031–1040. [Google Scholar] [CrossRef]
- Bashir, M.; Prietl, B.; Tauschmann, M.; Mautner, S.I.; Kump, P.K.; Treiber, G.; Wurm, P.; Gorkiewicz, G.; Högenauer, C.; Pieber, T.R. Effects of high doses of vitamin D3 on mucosa-associated gut microbiome vary between regions of the human gastrointestinal tract. Eur. J. Nutr. 2015, 55, 1479–1489. [Google Scholar] [CrossRef] [PubMed]
- Png, C.W.; Linden, S.K.; Gilshenan, K.S.; Zoetendal, E.G.; McSweeney, C.S.; Sly, L.I.; McGuckin, M.A.; Florin, T.H. Mucolytic bacteria with increased prevalence in IBD mucosa augment in vitro utilization of mucin by other bacteria. Am. J. Gastroenterol. 2010, 105, 2420–2428. [Google Scholar] [CrossRef] [PubMed]
- Loy, A.; Pfann, C.; Steinberger, M.; Hanson, B.; Herp, S.; Brugiroux, S.; Gomes Neto, J.C.; Boekschoten, M.V.; Schwab, C.; Urich, T.; et al. Lifestyle and horizontal gene transfer-mediated evolution of mucispirillum schaedleri, a core member of the murine gut microbiota. Msystems 2017, 2, e00171. [Google Scholar] [CrossRef] [PubMed]
- Berry, D.; Schwab, C.; Milinovich, G.; Reichert, J.; Ben Mahfoudh, K.; Decker, T.; Engel, M.; Hai, B.; Hainzl, E.; Heider, S.; et al. Phylotype-level 16s rRNA analysis reveals new bacterial indicators of health state in acute murine colitis. ISME J. 2012, 6, 2091–2106. [Google Scholar] [CrossRef] [PubMed]
- Van der Sluis, M.; De Koning, B.A.; De Bruijn, A.C.; Velcich, A.; Meijerink, J.P.; Van Goudoever, J.B.; Buller, H.A.; Dekker, J.; Van Seuningen, I.; Renes, I.B.; et al. Muc2-deficient mice spontaneously develop colitis, indicating that MUC2 is critical for colonic protection. Gastroenterology 2006, 131, 117–129. [Google Scholar] [CrossRef] [PubMed]
- Heazlewood, C.K.; Cook, M.C.; Eri, R.; Price, G.R.; Tauro, S.B.; Taupin, D.; Thornton, D.J.; Png, C.W.; Crockford, T.L.; Cornall, R.J.; et al. Aberrant mucin assembly in mice causes endoplasmic reticulum stress and spontaneous inflammation resembling ulcerative colitis. PLoS Med. 2008, 5, e54. [Google Scholar] [CrossRef] [PubMed]
- Shaw, K.A.; Bertha, M.; Hofmekler, T.; Chopra, P.; Vatanen, T.; Srivatsa, A.; Prince, J.; Kumar, A.; Sauer, C.; Zwick, M.E.; et al. Dysbiosis, inflammation, and response to treatment: A longitudinal study of pediatric subjects with newly diagnosed inflammatory bowel disease. Genome Med. 2016, 8, 75. [Google Scholar] [CrossRef] [PubMed]
- Earley, H.; Lennon, G.; Balfe, A.; Kilcoyne, M.; Clyne, M.; Joshi, L.; Carrington, S.; Martin, S.T.; Coffey, J.C.; Winter, D.C.; et al. A preliminary study examining the binding capacity of akkermansia muciniphila and desulfovibrio spp., to colonic mucin in health and ulcerative colitis. PLoS ONE 2015, 10, e0135280. [Google Scholar] [CrossRef] [PubMed]
- Schwarz, T. Mechanisms of UV–induced immunosuppression. Keio J. Med. 2005, 54, 165–171. [Google Scholar] [CrossRef] [PubMed]
- Gorman, S.; McGlade, J.P.; Lambert, M.J.; Strickland, D.H.; Thomas, J.A.; Hart, P.H. UV exposure and protection against allergic airways disease. Photochem. Photobiol. Sci. 2010, 9, 571–577. [Google Scholar] [CrossRef] [PubMed]
- Catana, C.S.; Berindan Neagoe, I.; Cozma, V.; Magdas, C.; Tabaran, F.; Dumitrascu, D.L. Contribution of the IL-17/IL-23 axis to the pathogenesis of inflammatory bowel disease. World J. Gastroenterol. 2015, 21, 5823–5830. [Google Scholar] [CrossRef] [PubMed]
- Wedebye Schmidt, E.G.; Larsen, H.L.; Kristensen, N.N.; Poulsen, S.S.; Lynge Pedersen, A.M.; Claesson, M.H.; Pedersen, A.E. Th17 cell induction and effects of IL-17a and IL-17f blockade in experimental colitis. Inflamm. Bowel Dis. 2013, 19, 1567–1576. [Google Scholar] [CrossRef] [PubMed]
- Hueber, W.; Sands, B.E.; Lewitzky, S.; Vandemeulebroecke, M.; Reinisch, W.; Higgins, P.D.; Wehkamp, J.; Feagan, B.G.; Yao, M.D.; Karczewski, M.; et al. Secukinumab, a human anti-IL-17a monoclonal antibody, for moderate to severe crohn’s disease: Unexpected results of a randomised, double-blind placebo-controlled trial. Gut 2012, 61, 1693–1700. [Google Scholar] [CrossRef] [PubMed]
- Bissonnette, R.; Luger, T.; Thaci, D.; Toth, D.; Lacombe, A.; Xia, S.; Mazur, R.; Patekar, M.; Charef, P.; Milutinovic, M.; et al. Secukinumab demonstrates high sustained efficacy and a favorable safety profile in patients with moderate to severe psoriasis through 5 years of treatment (sculpture extension study). J. Eur. Acad. Dermatol. Venereol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Nguyen, L.; Chun, R.F.; Lagishetty, V.; Ren, S.; Wu, S.; Hollis, B.; DeLuca, H.F.; Adams, J.S.; Hewison, M. Altered endocrine and autocrine metabolism of vitamin D in a mouse model of gastrointestinal inflammation. Endocrinology 2008, 149, 4799–4808. [Google Scholar] [CrossRef] [PubMed]
- Ghaly, S.; Lawrance, I. The role of vitamin D in gastrointestinal inflammation. Expert Rev. Gastroenterol. Hepatol. 2014, 8, 909–923. [Google Scholar] [CrossRef] [PubMed]
- Santarlasci, V.; Cosmi, L.; Maggi, L.; Liotta, F.; Annunziato, F. IL-1 and T helper immune responses. Front. Immunol. 2013, 4, 182. [Google Scholar] [CrossRef] [PubMed]
- Hiratsuka, T.; Inomata, M.; Goto, S.; Oyama, Y.; Nakano, T.; Chen, C.-L.; Shiraishi, N.; Noguchi, T.; Kitano, S. Phototherapy with artificial light suppresses dextran sulfate sodium–induced colitis in a mouse model. J. Gastroenterol. Hepatol. 2014, 29, 749–756. [Google Scholar] [CrossRef] [PubMed]
- Breuer, J.; Schwab, N.; Schneider-Hohendorf, T.; Marziniak, M.; Mohan, H.; Bhatia, U.; Gross, C.C.; Clausen, B.E.; Weishaupt, C.; Luger, T.A.; et al. Ultraviolet B light attenuates the systemic immune response in central nervous system autoimmunity. Ann. Neurol. 2014, 75, 739–758. [Google Scholar] [CrossRef] [PubMed]
- Lagishetty, V.; Misharin, A.V.; Liu, N.Q.; Lisse, T.S.; Chun, R.F.; Ouyang, Y.; McLachlan, S.M.; Adams, J.S.; Hewison, M. Vitamin D deficiency in mice impairs colonic antibacterial activity and predisposes to colitis. Endocrinology 2010, 151, 2423–2432. [Google Scholar] [CrossRef] [PubMed]
- Ooi, J.H.; Li, Y.; Rogers, C.J.; Cantorna, M.T. Vitamin D regulates the gut microbiome and protects mice from dextran sodium sulfate-induced colitis. J. Nutr. 2013, 143, 1679–1686. [Google Scholar] [CrossRef] [PubMed]
- Schaffler, H.; Herlemann, D.P.; Klinitzke, P.; Berlin, P.; Kreikemeyer, B.; Jaster, R.; Lamprecht, G. Vitamin D administration leads to a shift of the intestinal bacterial composition in Crohn’s disease patients, but not in healthy controls. J. Dig. Dis. 2018, 19, 225–234. [Google Scholar] [CrossRef] [PubMed]
- Garg, M.; Hendy, P.; Ding, J.N.; Shaw, S.; Hold, G.; Hart, A. The effect of vitamin D on intestinal inflammation and faecal microbiota in patients with ulcerative colitis. J. Crohns Colitis 2018, 1, 10. [Google Scholar] [CrossRef] [PubMed]
- Borton, M.A.; Sabag-Daigle, A.; Wu, J.; Solden, L.M.; O’Banion, B.S.; Daly, R.A.; Wolfe, R.A.; Gonzalez, J.F.; Wysocki, V.H.; Ahmer, B.M.M.; et al. Chemical and pathogen-induced inflammation disrupt the murine intestinal microbiome. Microbiome 2017, 5, 47. [Google Scholar] [CrossRef] [PubMed]
- Vinolo, M.A.; Rodrigues, H.G.; Nachbar, R.T.; Curi, R. Regulation of inflammation by short chain fatty acids. Nutrients 2011, 3, 858–876. [Google Scholar] [CrossRef] [PubMed]
- Sharifi, N.; Amani, R.; Hajiani, E.; Cheraghian, B. Women may respond different from men to vitamin D supplementation regarding cardiometabolic biomarkers. Exp. Biol. Med. (Maywood) 2016, 241, 830–838. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Day/Treatment | Group 1 | Group 2 | t-Value | p-Value |
|---|---|---|---|---|
| Day 35 | D++ UVR+ | D++ UVR− | 1.24 | 0.15 |
| D+ UVR+ | D+ UVR− | 0.79 | 0.84 | |
| D− UVR+ | D− UVR− | 0.89 | 0.67 | |
| D− UVR+ | D+ UVR− | 0.86 | 0.67 | |
| Day 63 | D++ UVR+ | D++ UVR− | 1.09 | 0.30 |
| D+ UVR+ | D+ UVR− | 1.03 | 0.39 | |
| D− UVR+ | D− UVR− | 0.79 | 0.84 | |
| D− UVR+ | D+ UVR− | 0.82 | 0.79 |
| D++UVR+ | D++UVR− | p | D+UVR+ | D+UVR− | p | D−UVR+ | D−UVR− | p | |
|---|---|---|---|---|---|---|---|---|---|
| IL-1β (pg/mL) | 35.0 ± 70.0 | 97.3 ± 194.6 | ns | 0 | 226.0 ± 226.0 | ns | 0 | 86.9 ± 78.0 | ns |
| TNF-α (pg/mL) | 335.4 ± 120.6 | 411.3 ± 274.2 | ns | 301.6 ± 99.5 | 392.2 ± 161.2 | ns | 230.2 ± 19.1 | 356.4 ± 107.6 | ns |
| IL-10 (pg/mL) | 53.8 ± 20.1 | 66.9 ± 31.9 | ns | 51.9 ± 3.8 | 98.1 ± 60.8 | ns | 40.4 ± 3.3 | 57.2 ± 12.2 | ns |
| IL-6 (pg/mL) | 7.9 ± 3.6 | 7.8 ± 7.2 | ns | 6.7 ± 1.0 | 18.5 ± 14.5 | ns | 6.1 ± 1.6 | 7.2 ± 3.1 | ns |
| IL-17 (pg/mL) | 146.2 ± 38.5 | 177.6 ± 53.2 | ns | 116.7 ± 30.1 | 207.9 ± 72.7 | 0.05 | 121.5 ± 12.0 | 159.7 ± 24.8 | ns |
| Group 1 | Group 2 | t | p-Value |
|---|---|---|---|
| D++ UVR+ | D++ UVR− | 0.92 | 0.61 |
| D+ UVR+ | D+ UVR− | 1.17 | 0.26 |
| D− UVR+ | D− UVR− | 1.42 | 0.034 |
| D− UVR+ | D+ UVR− | 1.52 | 0.021 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghaly, S.; Kaakoush, N.O.; Lloyd, F.; Gordon, L.; Forest, C.; Lawrance, I.C.; Hart, P.H. Ultraviolet Irradiation of Skin Alters the Faecal Microbiome Independently of Vitamin D in Mice. Nutrients 2018, 10, 1069. https://doi.org/10.3390/nu10081069
Ghaly S, Kaakoush NO, Lloyd F, Gordon L, Forest C, Lawrance IC, Hart PH. Ultraviolet Irradiation of Skin Alters the Faecal Microbiome Independently of Vitamin D in Mice. Nutrients. 2018; 10(8):1069. https://doi.org/10.3390/nu10081069
Chicago/Turabian StyleGhaly, Simon, Nadeem O. Kaakoush, Frances Lloyd, Lavinia Gordon, Cynthia Forest, Ian C. Lawrance, and Prue H. Hart. 2018. "Ultraviolet Irradiation of Skin Alters the Faecal Microbiome Independently of Vitamin D in Mice" Nutrients 10, no. 8: 1069. https://doi.org/10.3390/nu10081069
APA StyleGhaly, S., Kaakoush, N. O., Lloyd, F., Gordon, L., Forest, C., Lawrance, I. C., & Hart, P. H. (2018). Ultraviolet Irradiation of Skin Alters the Faecal Microbiome Independently of Vitamin D in Mice. Nutrients, 10(8), 1069. https://doi.org/10.3390/nu10081069