Lupinus mutabilis Extract Exerts an Anti-Diabetic Effect by Improving Insulin Release in Type 2 Diabetic Goto-Kakizaki Rats

 ,
,
Abstract
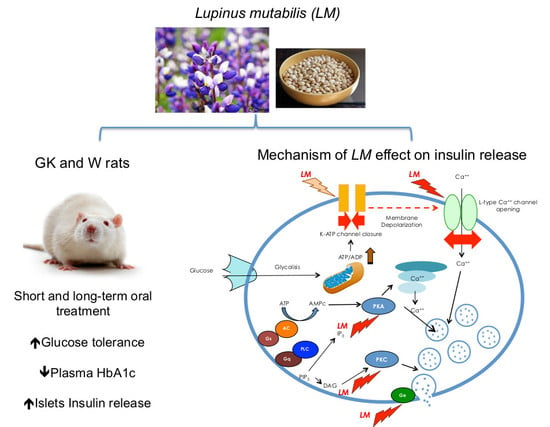
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Plant Material
2.3. Plant Extract Preparation
2.4. Gas Chromatography-Mass Spectrometry Analysis (GC-MS)
2.5. High Performance Liquid Chromatography-High Resolution Mass Spectrometry Analysis (HPLC-HRMS)
2.6. Oral Glucose Tolerance Test (OGTT)
2.7. Pancreatic Islets Isolation
2.8. Cytotoxicity
2.9. Islet Insulin Release
2.10. Islet Perifusion
2.11. Glucose Uptake Evaluation
2.12. Sub-Acute Oral Toxicity
2.13. Long-Term Treatment Evaluation
2.14. Mechanisms of LM-Dependent Insulin Release
2.15. Statistical Analysis
3. Results
3.1. LM Extract Constituents
3.2. LM Improves Glucose Tolerance in GK and W Rats by Increasing Serum Insulin Levels in a Glucose-Independent Manner
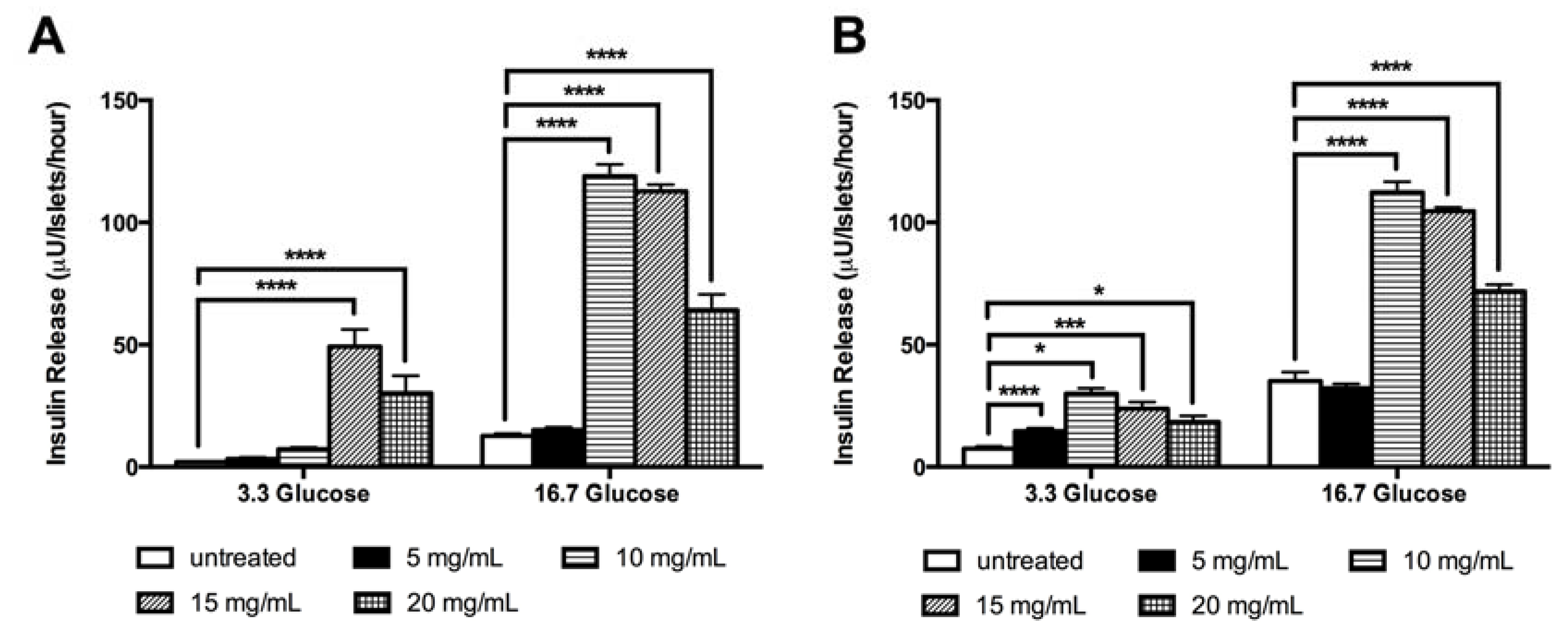
3.3. LM Stimulates The in vitro Insulin Release Independent of Glucose
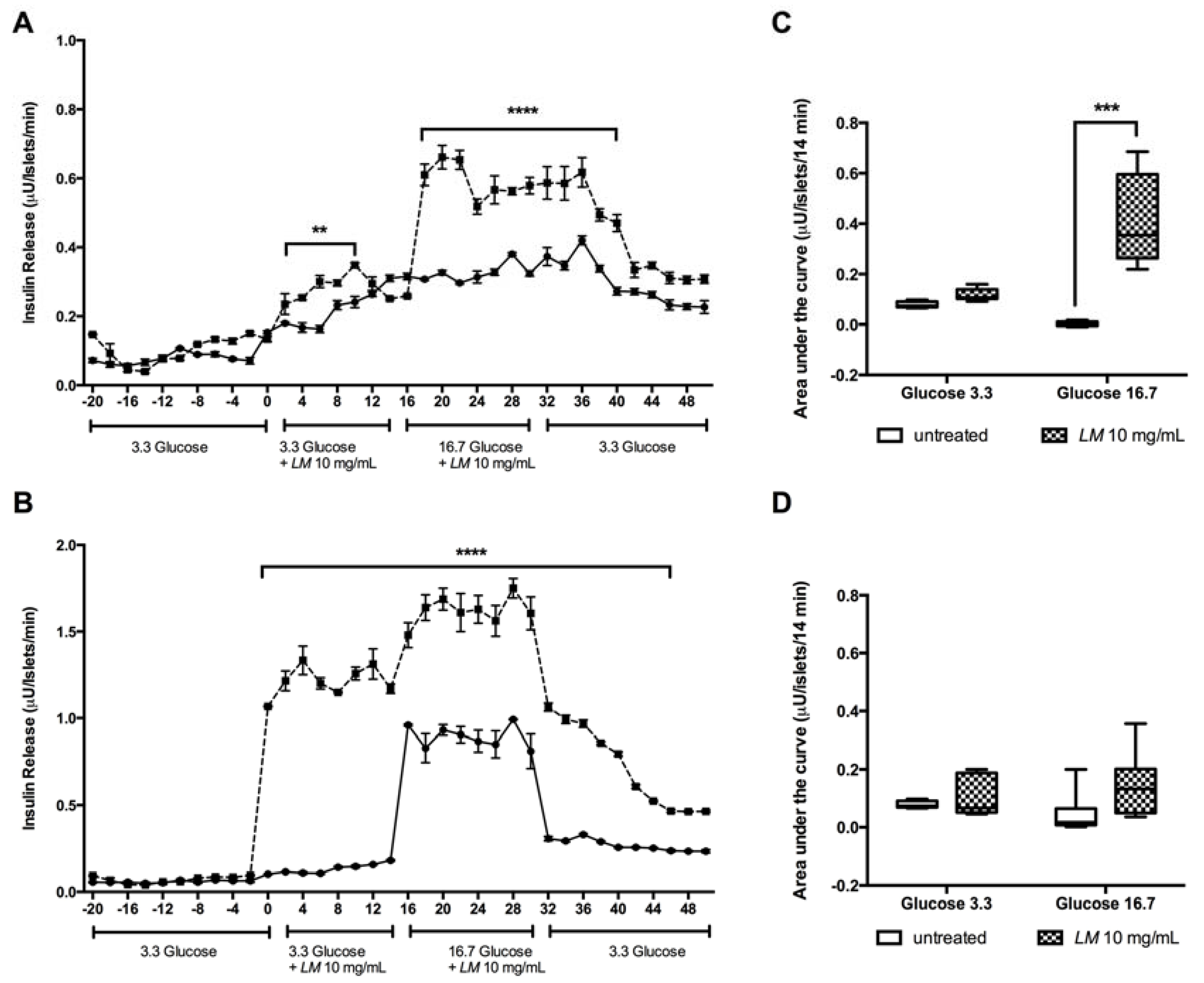
3.4. The LM Effect on The Kinetics of Insulin Release is Glucose Independent.
3.5. LM Does not Stimulates the in vitro Glucose Uptake
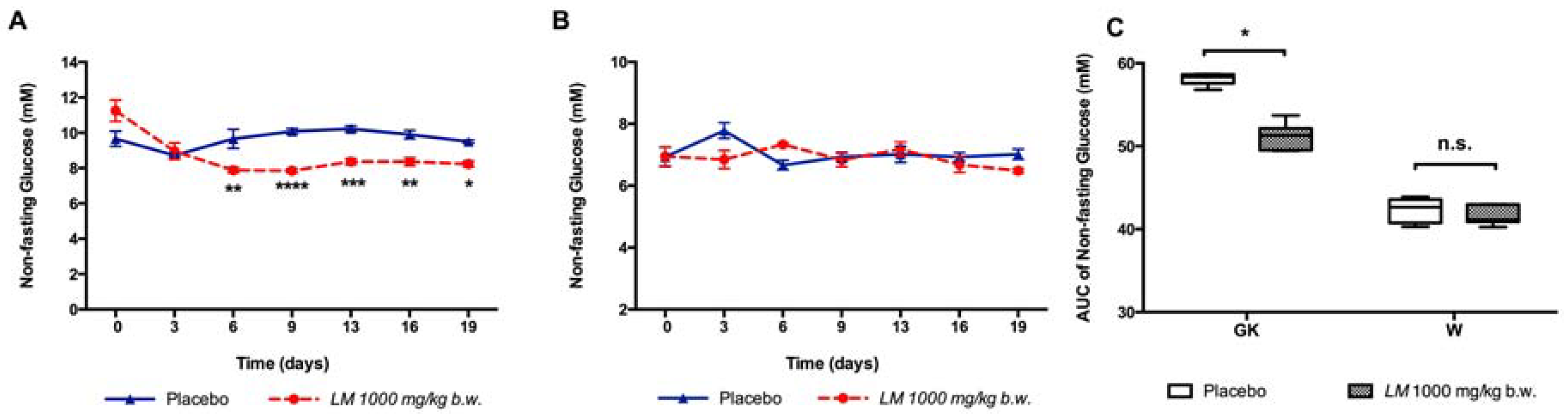
3.6. The Anti-Diabetic Effect of the Oral Long-Term LM Treatment is Mediated by the Increase of Insulin Release
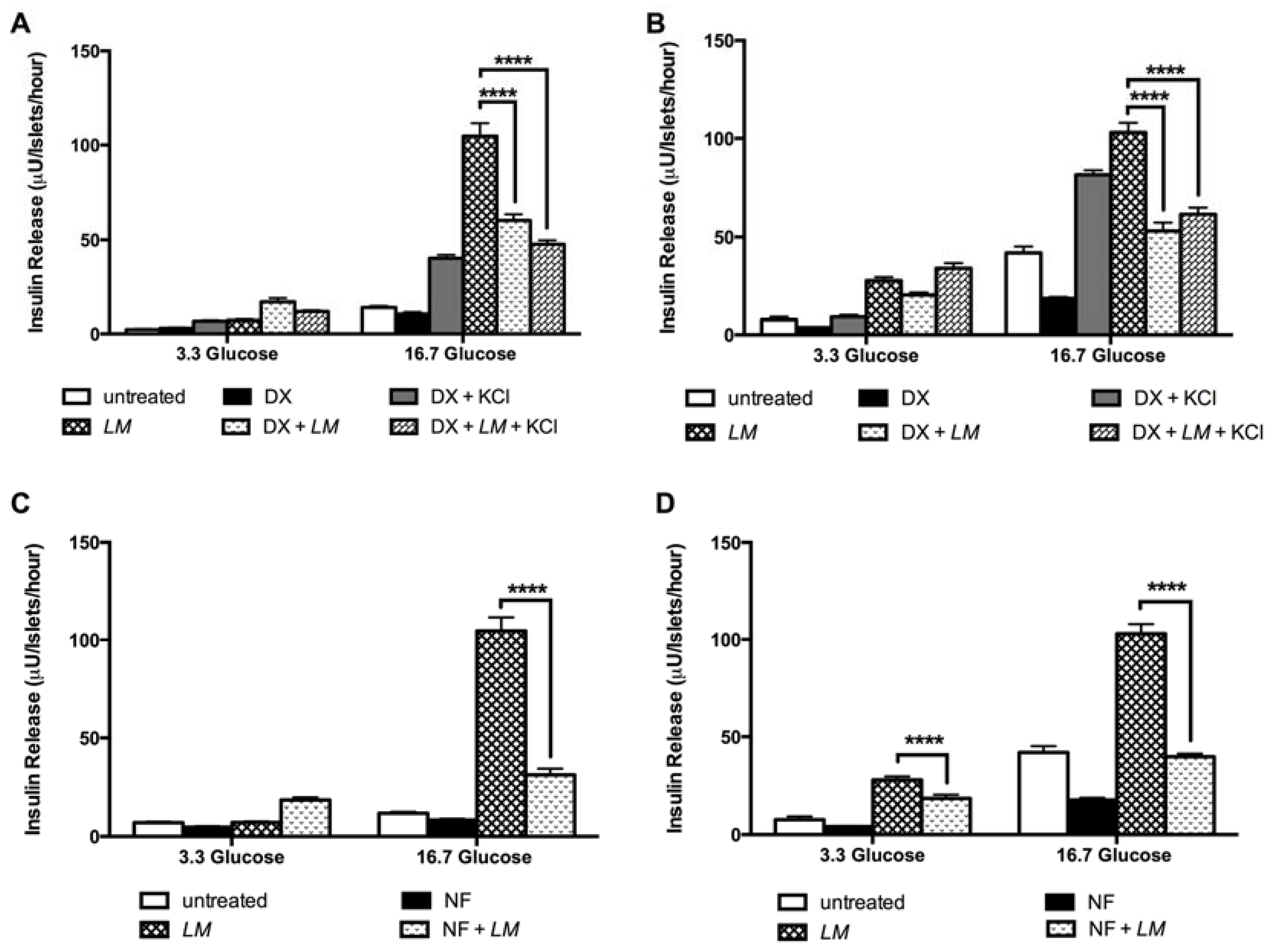
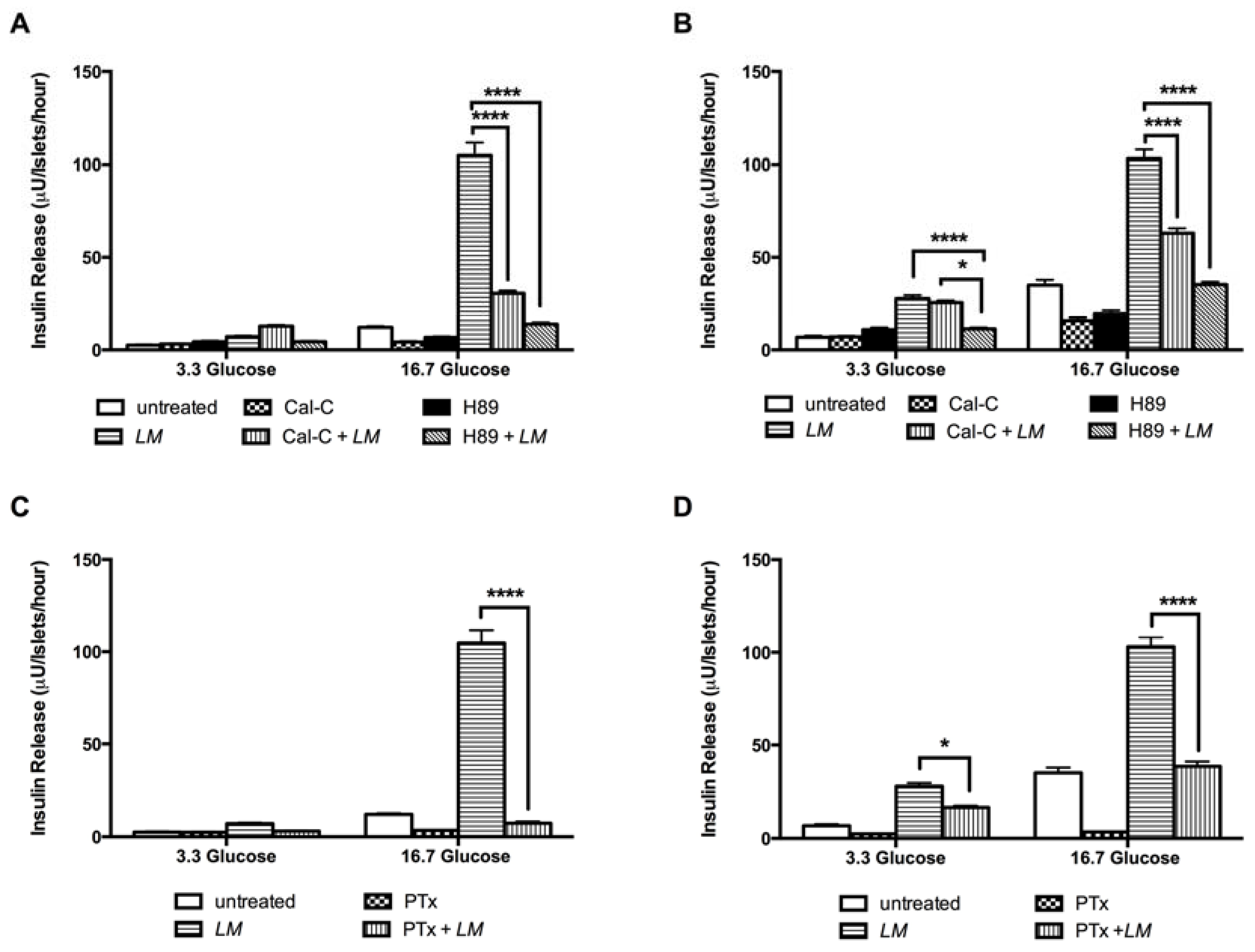
3.7. LM-Dependent Insulin Release is Mediated by L-Type Calcium Channel, PKC and PKA Systems, and Exocytosis by G-Proteins
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ogurtsova, K.; da Rocha Fernandes, J.D.; Huang, Y.; Linnenkamp, U.; Guariguata, L.; Cho, N.H.; Cavan, D.; Shaw, J.E.; Makaroff, L.E. IDF Diabetes Atlas: Global estimates for the prevalence of diabetes for 2015 and 2040. Diabetes Res. Clin. Pract. 2017, 128, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Alberti, K.G.; Zimmet, P.Z. Definition, diagnosis and classification of diabetes mellitus and its complications. Part 1: Diagnosis and classification of diabetes mellitus provisional report of a WHO consultation. Diabet. Med. 1998, 15, 539–553. [Google Scholar] [CrossRef]
- Ostenson, C.G. The pathophysiology of type 2 diabetes mellitus: An overview. Acta Physiol. Scand. 2001, 171, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Kahn, S.E.; Cooper, M.E.; Del Prato, S. Pathophysiology and treatment of type 2 diabetes: Perspectives on the past, present, and future. Lancet 2014, 383, 1068–1083. [Google Scholar] [CrossRef]
- Rios, J.L.; Francini, F.; Schinella, G.R. Natural Products for the Treatment of Type 2 Diabetes Mellitus. Planta. Med. 2015, 81, 975–994. [Google Scholar] [CrossRef] [PubMed]
- Dahlstrom, E.; Sandholm, N. Progress in Defining the Genetic Basis of Diabetic Complications. Curr. Diabetes Rep. 2017, 17, 80. [Google Scholar] [CrossRef] [PubMed]
- Firdous, S.M. Phytochemicals for treatment of diabetes. EXCLI J. 2014, 13, 451–453. [Google Scholar] [PubMed]
- Zhang, B.W.; Sang, Y.B.; Sun, W.L.; Yu, H.S.; Ma, B.P.; Xiu, Z.L.; Dong, Y.S. Combination of flavonoids from Oroxylum indicum seed extracts and acarbose improves the inhibition of postprandial blood glucose: In vivo and in vitro study. Biomed. Pharmacother. 2017, 91, 890–898. [Google Scholar] [CrossRef] [PubMed]
- Kim, C.S.; Sohn, E.J.; Kim, Y.S.; Jung, D.H.; Jang, D.S.; Lee, Y.M.; Kim, D.H.; Kim, J.S. Effects of KIOM-79 on hyperglycemia and diabetic nephropathy in type 2 diabetic Goto-Kakizaki rats. J. Ethnopharmacol. 2007, 111, 240–247. [Google Scholar] [CrossRef] [PubMed]
- Cicero, A.F.; Borghi, C. Evidence of clinically relevant efficacy for dietary supplements and nutraceuticals. Curr. Hypertens. Rep. 2013, 15, 260–267. [Google Scholar] [CrossRef] [PubMed]
- Santini, A.; Novellino, E.; Armini, V.; Ritieni, A. State of the art of Ready-to-Use Therapeutic Food: A tool for nutraceuticals addition to foodstuff. Food Chem. 2013, 140, 843–849. [Google Scholar] [CrossRef] [PubMed]
- Magrone, T.; Jirillo, E. Nutraceuticals and prevention of neurodegeneration. Curr. Pharm. Des. 2012, 18, 2–3. [Google Scholar] [CrossRef] [PubMed]
- Bahadoran, Z.; Mirmiran, P.; Azizi, F. Dietary polyphenols as potential nutraceuticals in management of diabetes: A review. J. Diabetes Metab. Disord. 2013, 12, 43. [Google Scholar] [CrossRef] [PubMed]
- Chirinos-Arias, M.C. Andean Lupin (Lupinus mutabilis Sweet) a plant with nutraceutical and medicinal potential. Rev. Bio Cienc. 2015, 3, 163–172. [Google Scholar]
- Jacobsen, S.-E.M.; Mujica, A. El tarwi (Lupinus mutabilis Sweet.) y sus parientes silvestres, in Botánica Económica de los Andes Centrales. Bot. Econ. Andes Cent. 2006, 28, 458–482. [Google Scholar]
- Fornasini, M.; Castro, J.; Villacrés, E.; Narváez, L.; Villamar, M.P.; Baldeón, M.E. Hypoglycemic effect of Lupinus mutabilis in healthy volunteers and subjects with dysglycemia. Nutr. Hosp. 2012, 27, 425–433. [Google Scholar] [PubMed]
- Baldeon, M.E.; Castro, J.; Villacrés, E.; Narváez, L.; Fornasini, M. Hypoglycemic effect of cooked Lupinus mutabilis and its purified alkaloids in subjects with type-2 diabetes. Nutr. Hosp. 2012, 27, 1261–1266. [Google Scholar] [PubMed]
- Ostenson, C.G.; Efendic, S. Islet gene expression and function in type 2 diabetes; studies in the Goto-Kakizaki rat and humans. Diabetes Obes. Metab. 2007, 9 (Suppl. 2), 180–186. [Google Scholar] [CrossRef] [PubMed]
- Ling, Z.C.; Efendic, S.; Wibom, R.; Abdel-Halim, S.M.; Ostenson, C.G.; Landau, B.R.; Khan, A. Glucose metabolism in Goto-Kakizaki rat islets. Endocrinology 1998, 139, 2670–2675. [Google Scholar] [PubMed]
- Mosen, H.; Salehi, A.; Alm, P.; Henningsson, R.; Jimenez-Feltström, J.; Ostenson, C.G.; Efendic, S.; Lundquist, I. Defective glucose-stimulated insulin release in the diabetic Goto-Kakizaki (GK) rat coincides with reduced activity of the islet carbon monoxide signaling pathway. Endocrinology 2005, 146, 1553–1558. [Google Scholar] [CrossRef] [PubMed]
- Ostenson, C.G.; Khan, A.; Abdel-Halim, S.M.; Guenifi, A.; Suzuki, K.; Goto, Y.; Efendic, S. Abnormal insulin secretion and glucose metabolism in pancreatic islets from the spontaneously diabetic GK rat. Diabetologia 1993, 36, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Zambrana, S.; Lundqvist Lena, C.E.; Veliz, V.; Catrina, S.-B.; Gonzales, E.; Östenson, C.-G. Amaranthus caudatus Stimulates Insulin Secretion in Goto-Kakizaki Rats, a Model of Diabetes Mellitus Type 2. Nutrients 2018, 10, 94. [Google Scholar] [CrossRef] [PubMed]
- Herbert, V.; Herbert, V.; Lau, K.S.; Gottlieb, C.W.; Bleicher, S.J. Coated charcoal immunoassay of insulin. J. Clin. Endocrinol. Metab. 1965, 25, 1375–1384. [Google Scholar] [CrossRef] [PubMed]
- Lokman, E.F.; Gu, H.F.; Wan Mohamud, W.N.; Östenson, C.-G. Evaluation of Antidiabetic Effects of the Traditional Medicinal Plant Gynostemma pentaphyllum and the Possible Mechanisms of Insulin Release. Evid. Based Complement. Altern. Med. 2015, 2015, 120572. [Google Scholar] [CrossRef] [PubMed]
- Hoa, N.K.; Norberg, A.; Sillard, R.; Van Phan, D.; Thuan, N.D.; Dzung, D.T.; Jörnvall, H.; Ostenson, C.-G. The possible mechanisms by which phanoside stimulates insulin secretion from rat islets. J. Endocrinol. 2007, 192, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Mosmann, T. Rapid colorimetric assay for cellular growth and survival: Application to proliferation and cytotoxicity assays. J. Immunol. Methods 1983, 65, 55–63. [Google Scholar] [CrossRef]
- Catrina, S.-B.; Catrina, A.I.; Sirzen, F.; Griffithis, W.; Bergman, T.; Bigerfeldal, D.; Coculescu, M.; Mutt, V. A cytotoxic, apoptotic, low-molecular weight factor from pineal gland. Life Sci. 1999, 65, 1047–1057. [Google Scholar] [CrossRef]
- Drakenberg, K.; Ostenson, C.-G.; Sara, V. Circulating forms and biological activity of intact and truncated insulin-like growth factor I in adult and neonatal rats. Acta Endocrinol. 1990, 123, 43–50. [Google Scholar] [CrossRef] [PubMed]
- Domingues, N.; Pelletier, J.; Ostenson, C.-G.; Castro, M.M. Therapeutic properties of VO(dmpp)2 as assessed by in vitro and in vivo studies in type 2 diabetic GK rats. J. Inorg. Biochem. 2014, 131, 115–122. [Google Scholar] [CrossRef] [PubMed]
- OECD, Test No. 407: Repeated Dose 28-day Oral Toxicity Study in Rodents. OECD Publishing. Available online: https://ntp.niehs.nih.gov/iccvam/suppdocs/feddocs/oecd/oecdtg407-2008 (accessed on 10 May 2013).
- Girija, K.; Lakshman, K.; Udaya, C.; Sabhya, S.G.; Divya, T. Anti-diabetic and anti-cholesterolemic activity of methanol extracts of three species of Amaranthus. Asian Pac. J. Trop. Biomed. 2011, 1, 133–138. [Google Scholar] [CrossRef]
- Pelletier, J.; Domingues, N.; Castro, M.M.; Östenson, C.-G. In vitro effects of bis(1,2-dimethyl-3-hydroxy-4-pyridinonato) oxidovanadium (IV), or VO(dmpp)2, on insulin secretion in pancreatic islets of type 2 diabetic Goto-Kakizaki rats. J. Inorg. Biochem. 2016, 154, 29–34. [Google Scholar] [CrossRef] [PubMed]
- Guemes-Vera, N.; Peña Bautista, R.J.; Jimenez-Martinez, C.; Davila-Ortiz, G.; Calderon-Dominguez, G. Effective detoxification and decoloration of Lupinus mutabilis seed derivatives, and effect of these derivatives on bread quality and acceptance. J. Sci. Food Agric. 2008, 88, 1135–1143. [Google Scholar] [CrossRef]
- Torres Tello, F.; Nagata, A.; Dreifuss Spiegel, W. Methods of eliminating alkaloids from the seeds of Lupinus mutabilis Sweet. Arch Latinoam. Nutr. 1980, 30, 200–209. [Google Scholar] [PubMed]
- Ortiz, C.; Gross, R.; von Baer, E. The lupine, a contribution to nutrition in the Andes. 2. The quality of proteins in Lupinus mutabilis compared to Lupinus albus, Lupinus luteus and Soja max. Z. Ernahrungswiss. 1975, 14, 229–233. [Google Scholar] [CrossRef] [PubMed]
- Schoeneberger, H.; Gross, R.; Cremer, H.D.; Elmadfa, I. Composition and protein quality of Lupinus mutabilis. J. Nutr. 1982, 112, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Ortega Duarte, A.; Martin-Sanchez, F.J.; Gonzalez-Castillo, J.; Ruiz-Artacho, P. Lupinus mutabilis (chocho) water intoxication. Med. Clin. 2013, 140, 43–44. [Google Scholar] [CrossRef] [PubMed]
- Garcia Lopez, P.M.; de la Mora, P.G.; Wysocka, W.; Maiztegui, B.; Alzugaray, M.E.; Del Zotto, H.; Borelli, M.I. Quinolizidine alkaloids isolated from Lupinus species enhance insulin secretion. Eur, J. Pharmacol. 2004, 504, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Kroc, M.; Rybiński, W.; Wilczura, P.; Kamel, K.; Kaczmarek, Z.; Barzyk, P.; Święcicki, W. Quantitative and qualitative analysis of alkaloids composition in the seeds of a white lupin (Lupinus albus L.) collection. Genet. Resour. Crop Evolut. 2017, 64, 1853–1860. [Google Scholar] [CrossRef]
- Wiedemann, M.; Gurrola-Díaz, C.M.; Vargas-Guerrero, B.; Wink, M.; García-López, P.M.; Düfer, M. Lupanine Improves Glucose Homeostasis by Influencing KATP Channels and Insulin Gene Expression. Molecules 2015, 20, 19085–19100. [Google Scholar] [CrossRef] [PubMed]
- Paolisso, G.; Sgambato, S.; Passariello, N.; Pizza, G.; Torella, R.; Tesauro, P.; Varricchio, M.; D’Onofrio, F. Plasma glucose lowering effect of sparteine sulphate infusion in non-insulin-dependent (type 2) diabetic subjects. Eur. J. Clin. Pharmacol. 1988, 34, 227–232. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M.; Newgard, C.B. Mechanisms of disease: Molecular and metabolic mechanisms of insulin resistance and beta-cell failure in type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 193–205. [Google Scholar] [CrossRef] [PubMed]
- Mears, D. Regulation of insulin secretion in islets of Langerhans by Ca2+ channels. J. Membr. Biol. 2004, 200, 57–66. [Google Scholar] [CrossRef] [PubMed]
- Wan, Q.F.; Dong, Y.; Yang, H.; Lou, X.; Ding, J.; Xu, T. Protein Kinase Activation Increases Insulin Secretion by Sensitizing the Secretory Machinery to Ca2+. J. Gener. Physiol. 2004, 124, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Ahren, B. Islet G protein-coupled receptors as potential targets for treatment of type 2 diabetes. Nat. Rev. Drug Discov. 2009, 8, 369–385. [Google Scholar] [CrossRef] [PubMed]
- Bobkiewicz-Kozlowska, T.; Dworacka, M.; Kuczyński, S.; Abramczyk, M.; Kolanoś, R.; Wysocka, W.; Garcia Lopez, P.M.; Winiarska, H. Hypoglycaemic effect of quinolizidine alkaloids—Lupanine and 2-thionosparteine on non-diabetic and streptozotocin-induced diabetic rats. Eur. J. Pharmacol. 2007, 565, 240–244. [Google Scholar] [CrossRef] [PubMed]
- Lokman, F.E.; Gu, H.F.; Wan Mohamud, W.N.; Yusoff, M.M.; Chia, K.L.; Ostenson, C.-G. Antidiabetic Effect of Oral Borapetol B Compound, Isolated from the Plant Tinospora crispa, by Stimulating Insulin Release. Evid. Based Complement. Altern. Med. 2013, 2013, 727602. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Time (min) | 0 | 30 | 60 | 90 | 120 | AUC |
|---|---|---|---|---|---|---|
| Group 1. GK + LM 1000 mg/kg b.w. | ||||||
| Day 0 | 7.8 ± 0.2 | 20.5 ± 0.2 | 19.0 ± 0.3 | 16.4 ± 0.2 # | 12.8 ± 0.4 | 1049.7 ± 26.9 |
| Day 10 | 6.8 ± 0.2 | 19.4 ± 1.1 | 17.5 ± 0.4 | 14,4 ± 0.5 #### | 11.9 ± 0.4 | 1004.8 ± 48.0 |
| Day 20 | 6.4 ± 0.3 | 18.1 ± 0.2 ## | 15.6 ± 0.1 #### | 11.9 ± 0.4 #### | 8.7 ± 0.4 #### | 827.3 ± 24.8 ## |
| Group 2. GK + Placebo | ||||||
| Day 0 | 7.0 ± 0.2 | 21.3 ± 0.5 | 19.2 ± 0.3 | 17.5 ± 0.2 | 13.4± 0.4 | 1209.0 ± 39.0 |
| Day 10 | 7.1 ± 0.6 | 20.4 ± 0.3 | 19.4 ± 0.3 | 17.6 ± 0.4 | 12.5 ± 0.4 | 1162.0 ± 70.0 |
| Day 20 | 6.9 ± 0.2 | 19.9 ± 0.3 | 18.7 ± 0.2 | 16.7 ± 0.2 | 11.8 ± 0.7 | 1110.6 ± 19.3 |
| Group 3. W + LM 1000 mg/kg b.w. | ||||||
| Day 0 | 4.1 ± 0.3 | 11.7 ± 0.4 | 9.8 ± 0.4 | 7.3 ± 0.3 | 6.1 ± 0.4 | 525.5 ± 35.2 |
| Day 10 | 4.1 ± 0.1 | 10.5 ± 0.7 ### | 9.1 ± 0.6 # | 6.9 ± 0.2 | 5.8 ± 0.5 | 453.5 ± 42.0 |
| Day 20 | 4.1 ± 0.1 | 9.3 ± 0.1 #### | 7.7 ± 0.1 #### | 6.4 ± 0.1 #### | 5.4 ± 0.1 ### | 354.3 ± 12.2 # |
| Group 4. W + Placebo | ||||||
| Day 0 | 3.7 ± 0.2 | 12.3 ± 0.1 | 10.1 ± 0.1 | 7.4 ± 0.1 | 6.1 ± 0.2 | 593.2 ± 15.0 |
| Day 10 | 4.3 ± 0.3 | 12.1 ± 0.1 | 10.3 ± 0.1 | 7.5 ± 0.1 | 6.0 ± 0.2 | 534.5 ± 30.4 |
| Day 20 | 4.6 ± 0.3 | 12.1 ± 0.3 | 10.2 ± 0.2 | 7.8 ± 0.2 | 6.3 ± 0.1 | 507.5 ± 40.1 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zambrana, S.; Lundqvist, L.C.E.; Mamani, O.; Catrina, S.-B.; Gonzales, E.; Östenson, C.-G. Lupinus mutabilis Extract Exerts an Anti-Diabetic Effect by Improving Insulin Release in Type 2 Diabetic Goto-Kakizaki Rats. Nutrients 2018, 10, 933. https://doi.org/10.3390/nu10070933
Zambrana S, Lundqvist LCE, Mamani O, Catrina S-B, Gonzales E, Östenson C-G. Lupinus mutabilis Extract Exerts an Anti-Diabetic Effect by Improving Insulin Release in Type 2 Diabetic Goto-Kakizaki Rats. Nutrients. 2018; 10(7):933. https://doi.org/10.3390/nu10070933
Chicago/Turabian StyleZambrana, Silvia, Lena C. E. Lundqvist, Orlando Mamani, Sergiu-Bogdan Catrina, Eduardo Gonzales, and Claes-Göran Östenson. 2018. "Lupinus mutabilis Extract Exerts an Anti-Diabetic Effect by Improving Insulin Release in Type 2 Diabetic Goto-Kakizaki Rats" Nutrients 10, no. 7: 933. https://doi.org/10.3390/nu10070933
APA StyleZambrana, S., Lundqvist, L. C. E., Mamani, O., Catrina, S.-B., Gonzales, E., & Östenson, C.-G. (2018). Lupinus mutabilis Extract Exerts an Anti-Diabetic Effect by Improving Insulin Release in Type 2 Diabetic Goto-Kakizaki Rats. Nutrients, 10(7), 933. https://doi.org/10.3390/nu10070933