Aqueous Extract of Pepino (Solanum muriactum Ait) Leaves Ameliorate Lipid Accumulation and Oxidative Stress in Alcoholic Fatty Liver Disease



Abstract
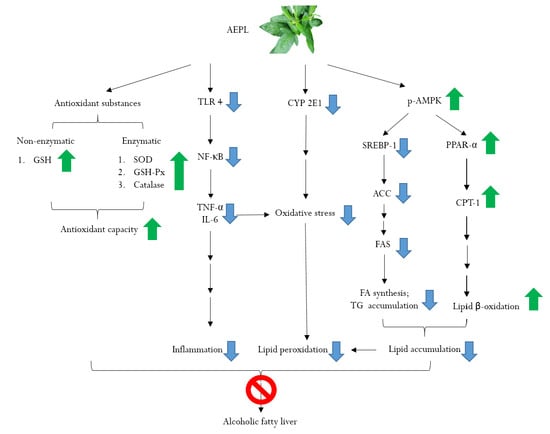
:
1. Introduction
2. Materials and Methods
2.1. Preparation of AEPL and Alcohol-Containing Liquid Diet
2.2. Animals and Experimental Design
2.3. Serum Biochemical Parameters
2.4. Measurement of Thiobarbituric Acid Relative Substances (TBARS) and Antioxidant Status in Liver
2.5. Trolox Equivalent Antioxidant Capacity (TEAC) Assay
2.6. Measurement of Liver TG Levels
2.7. Measurement of Liver Tumor NecrosisFfactor (TNF)-α and Interleukin (IL)-6 Levels
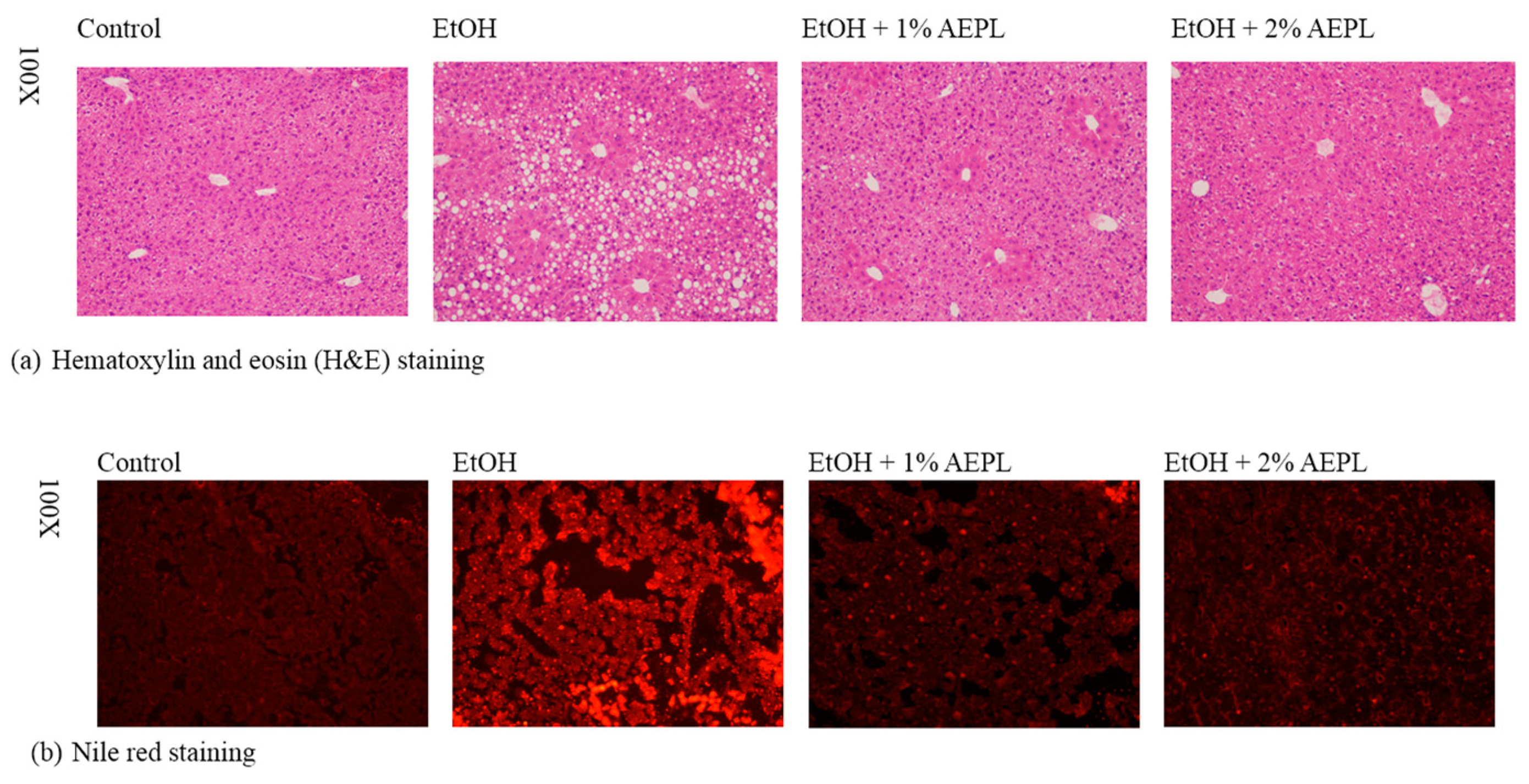
2.8. Histological Evaluation
2.9. Protein Preparation and Western Blot Analysis
2.10. Statistical Analysis
3. Results
3.1. AEPL Helped Maintain Mice Body Weight
3.2. AEPL Diminished Serum AST, ALT, TC, TG, LDL, and VLDL, and Hepatic TG Level
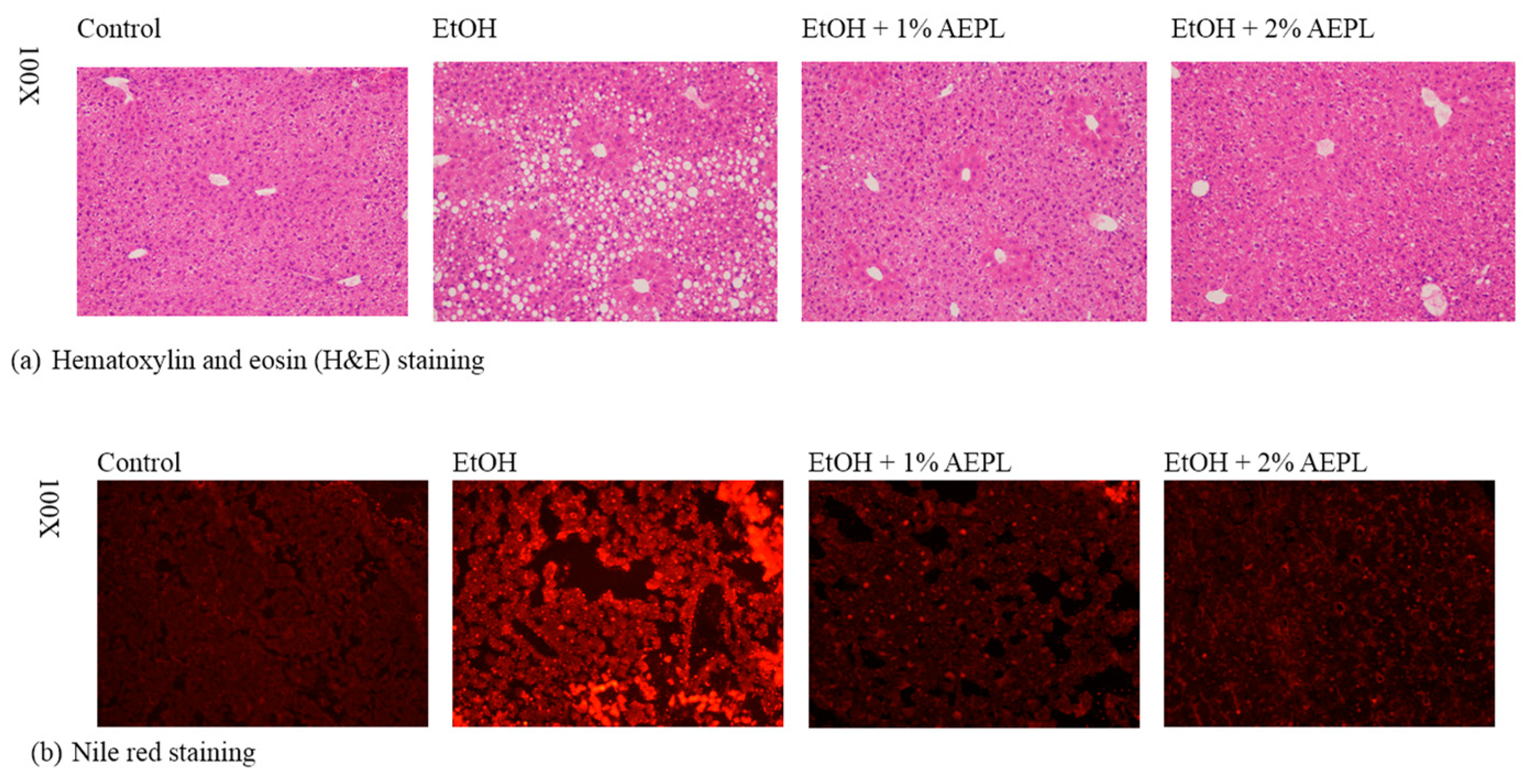
3.3. AEPL Attenuated Alcohol-Induced Liver Lipid Accumulation Via Histological Assessment
3.4. AEPL Improved the Lipid Metabolism by Lipid Synthesis and Oxidation
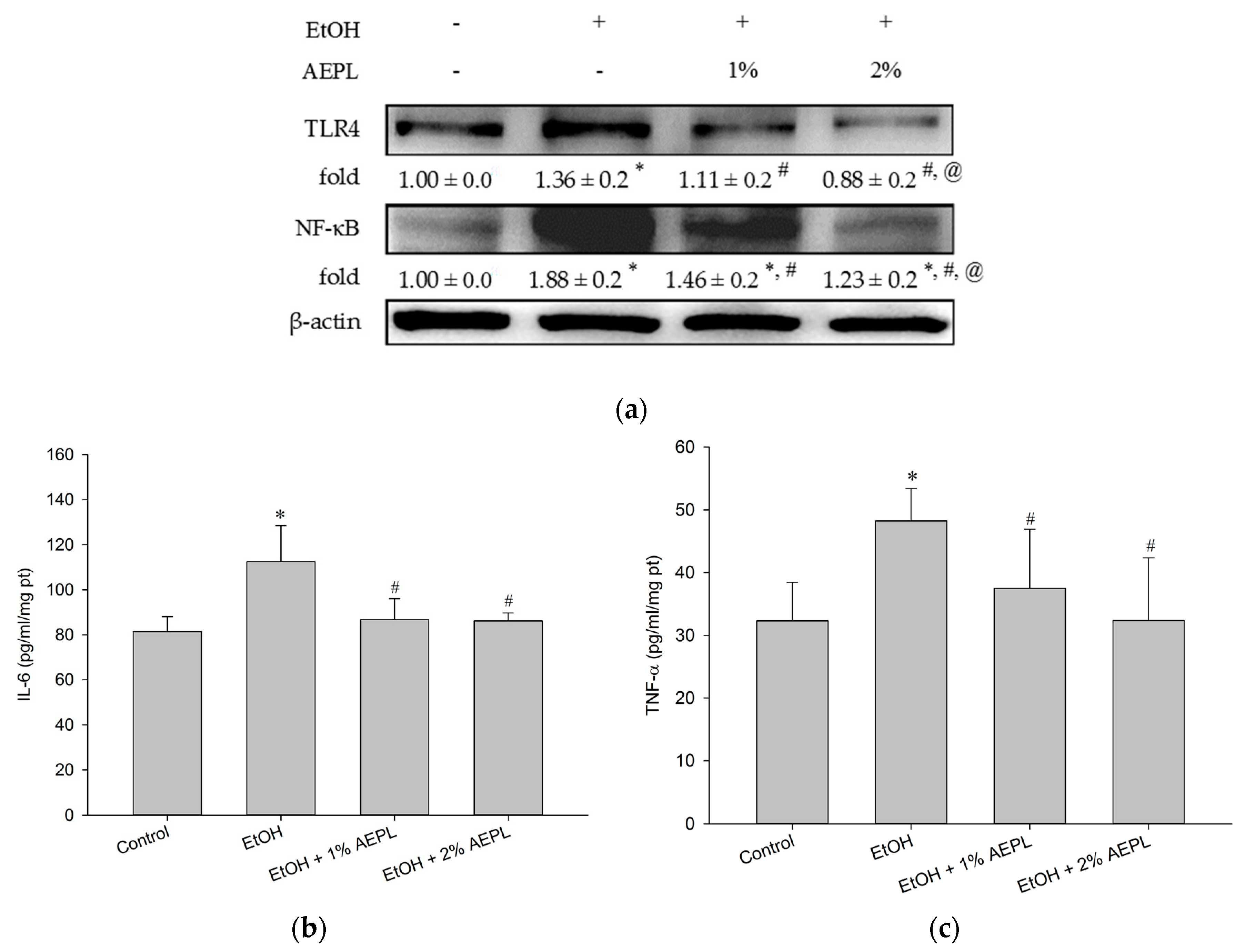
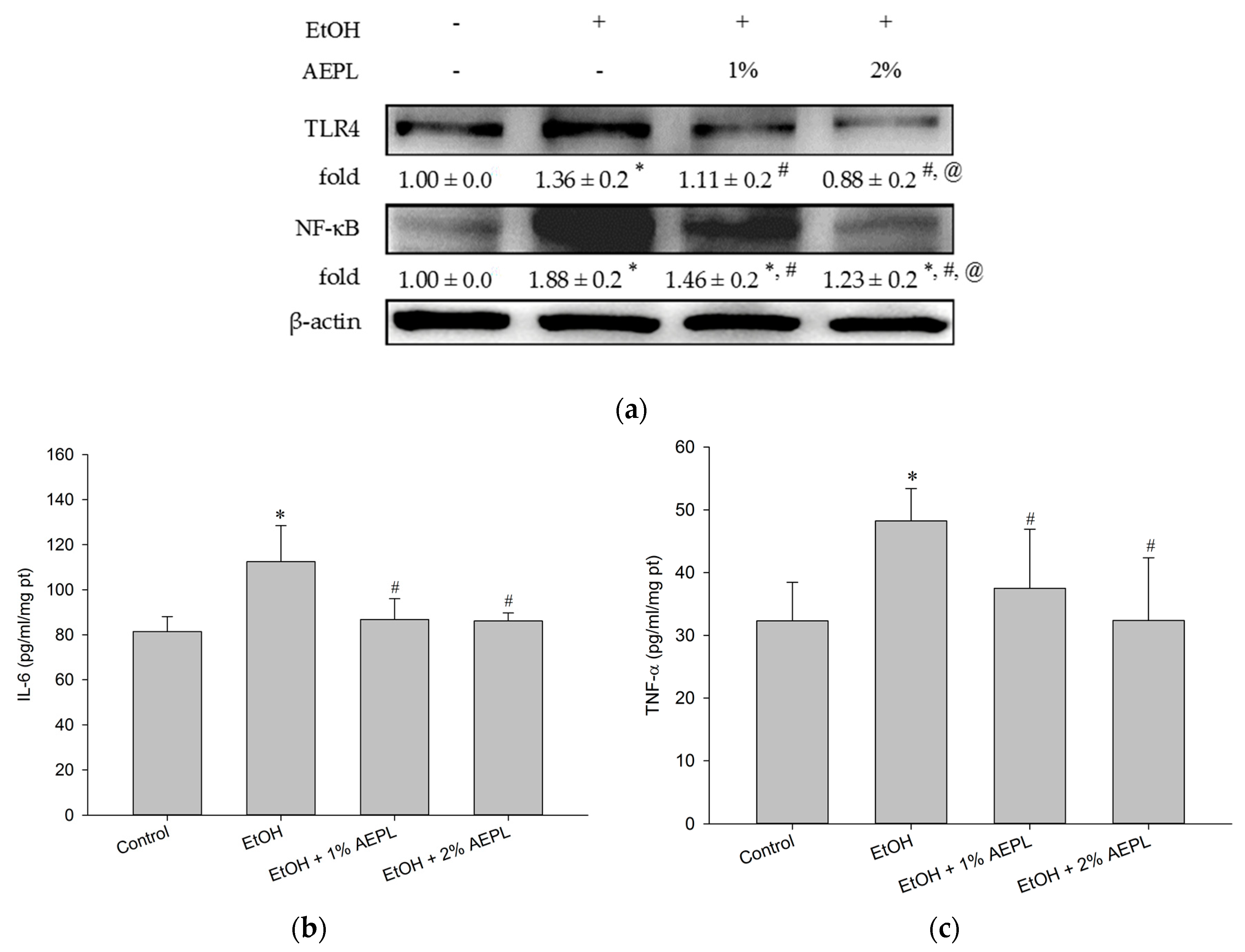
3.5. AEPL Could Suppress the Expression of Liver Cytochrome P 2E1
3.6. AEPL Reduced Alcohol-Induced Inflammatory Cytokines Levels
3.7. AEPL Increased Antioxidant Substances and Decreased Lipid Peroxidation
4. Discussion
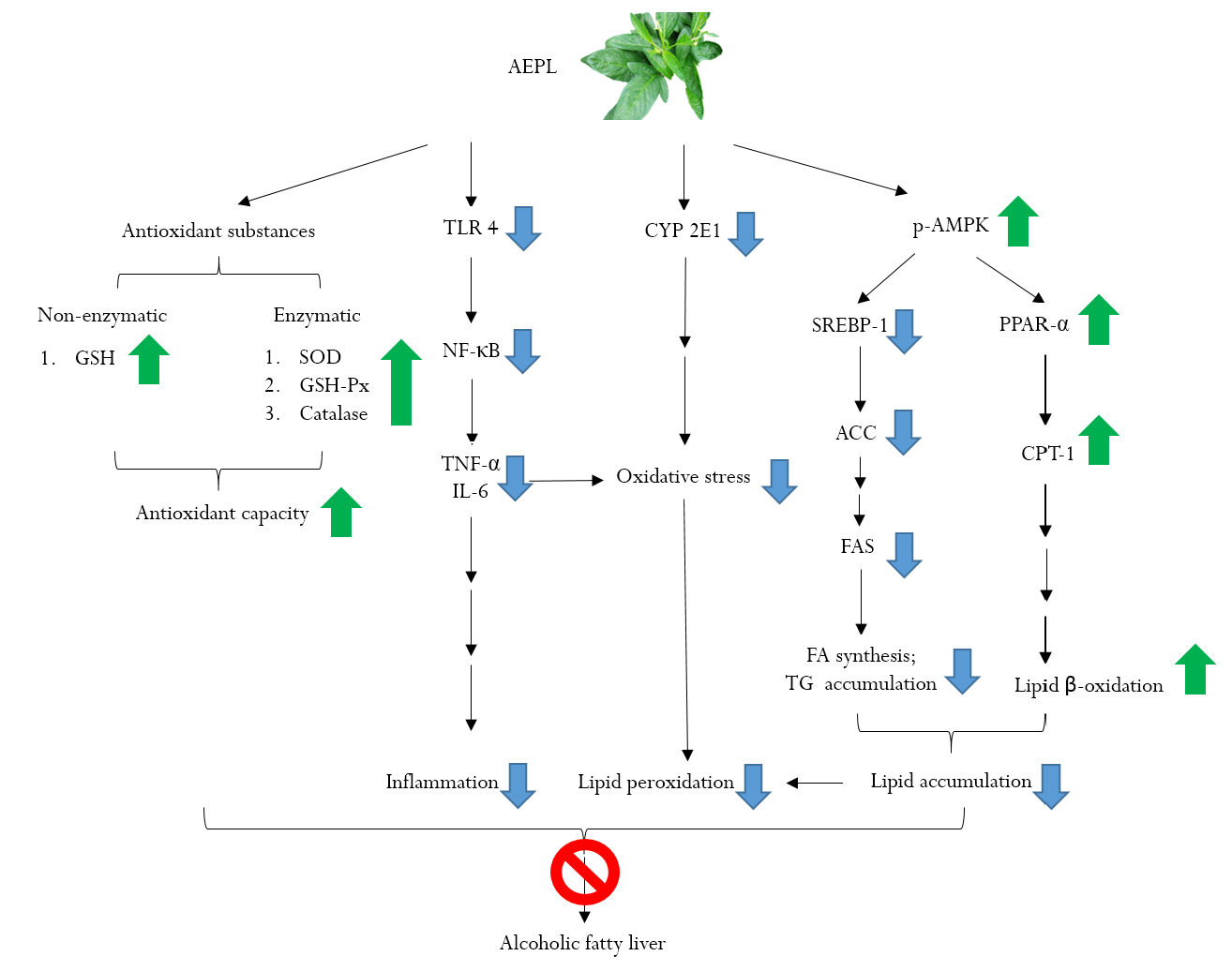
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Gao, B.; Bataller, R. Alcoholic liver disease: Pathogenesis and new therapeutic targets. Gastroenterology 2011, 141, 1572–1585. [Google Scholar] [CrossRef] [PubMed]
- Liu, J. Ethanol and liver: Recent insights into the mechanisms of ethanol-induced fatty liver. World J. Gastroenterol. 2014, 20, 14672–14685. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, S.; Mihaylova, M.M.; Zheng, B.; Hou, X.; Jiang, B.; Park, O.; Luo, Z.; Lefai, E.; Shyy, J.Y.; et al. AMPK phosphorylates and inhibits SREBP activity to attenuate hepatic steatosis and atherosclerosis in diet-induced insulin-resistant mice. Cell Metab. 2011, 13, 376–388. [Google Scholar] [CrossRef] [PubMed]
- Rakhshandehroo, M.; Knoch, B.; Muller, M.; Kersten, S. Peroxisome proliferator-activated receptor alpha target genes. PPAR Res. 2010, 61, 393–416. [Google Scholar] [CrossRef] [PubMed]
- Birben, E.; Sahiner, U.M.; Sackesen, C.; Erzurum, S.; Kalayci, O. Oxidative stress and antioxidant defense. World Allergy Organ J. 2012, 5, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.; Ande, A.; Kumar, A.; Kumar, S. Regulation of cytochrome P450 2e1 expression by ethanol: Role of oxidative stress-mediated pkc/jnk/sp1 pathway. Cell Death Dis. 2013, 4, e554. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Cederbaum, A.I. CYP2E1 and oxidative liver injury by alcohol. Free Radic. Biol. Med. 2008, 44, 723–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sid, B.; Verrax, J.; Calderon, P.B. Role of oxidative stress in the pathogenesis of alcohol-induced liver disease. Free Radic. Res. 2013, 47, 894–904. [Google Scholar] [CrossRef] [PubMed]
- Ayala, A.; Munoz, M.F.; Arguelles, S. Lipid peroxidation: Production, metabolism, and signaling mechanisms of malondialdehyde and 4-hydroxy-2-nonenal. Oxid. Med. Cell Longev. 2014, 2014, 360438. [Google Scholar] [CrossRef] [PubMed]
- Han, K.H.; Hashimoto, N.; Fukushima, M. Relationships among alcoholic liver disease, antioxidants, and antioxidant enzymes. World J. Gastroenterol. 2016, 22, 37–49. [Google Scholar] [CrossRef] [PubMed]
- Zhou, T.; Zhang, Y.J.; Xu, D.P.; Wang, F.; Zhou, Y.; Zheng, J.; Li, Y.; Zhang, J.J.; Li, H.B. Protective effects of lemon juice on alcohol-induced liver injury in mice. Biomed. Res. Int. 2017, 2017, 7463571. [Google Scholar] [CrossRef] [PubMed]
- Park, H.S.; Jung, H.Y.; Park, E.Y.; Kim, J.; Lee, W.J.; Bae, Y.S. Cutting edge: Direct interaction of TLR4 with NAD(P)H oxidase 4 isozyme is essential for lipopolysaccharide-induced production of reactive oxygen species and activation of NF-kB. J. Immunol. 2004, 173, 3589–3593. [Google Scholar] [CrossRef] [PubMed]
- Chung, D.J.; Yang, M.Y.; Li, Y.R.; Chen, W.J.; Hung, C.Y.; Wang, C.J. Ganoderma lucidum repress injury of ethanol-induced steatohepatitis via anti-inflammation, anti-oxidation and reducing hepatic lipid in C57BL/6J mice. J. Funct. Foods 2017, 33, 314. [Google Scholar] [CrossRef]
- Tang, C.C.; Huang, H.P.; Lee, Y.J.; Tang, Y.H.; Wang, C.J. Hepatoprotective effect of mulberry water extracts on ethanol-induced liver injury via anti-inflammation and inhibition of lipogenesis in C57BL/6J mice. Food Chem. Toxicol. 2013, 62, 786–796. [Google Scholar] [CrossRef] [PubMed]
- He, P.; Wu, Y.; Shun, J.; Liang, Y.; Cheng, M.; Wang, Y. Baicalin ameliorates liver injury induced by chronic plus binge ethanol feeding by modulating oxidative stress and inflammation via CYP2E1 and NRF2 in mice. Oxid. Med. Cell Longev. 2017, 2017, 4820414. [Google Scholar] [CrossRef] [PubMed]
- Ren, W.; Tang, D.G. Extract of solanum muricatum (Pepino/CSG) inhibits tumor growth by inducing apoptosis. Anticancer Res. 1999, 19, 403–408. [Google Scholar] [PubMed]
- Hsu, C.C.; Guo, Y.R.; Wang, Z.H.; Yin, M.C. Protective effects of an aqueous extract from pepino (solanum muricatum Ait.) in diabetic mice. J. Sci. Food Agric. 2011, 91, 1517–1522. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.T.; Chyau, C.C.; Hsu, C.C.; Kuo, S.M.; Chuang, C.W.; Lin, H.H.; Chen, J.H. Pepino polyphenolic extract improved oxidative, inflammatory and glycative stress in the sciatic nerves of diabetic mice. Food Funct. 2016, 7, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Yagi, K. A simple fluorometric assay for lipoperoxide in blood plasma. Biochem. Med. 1976, 15, 212–216. [Google Scholar] [CrossRef]
- Eyer, P.; Podhradsky, D. Evaluation of the micromethod for determination of glutathione using enzymatic cycling and ellman’s reagent. Anal. Biochem. 1986, 153, 57–66. [Google Scholar] [CrossRef]
- Knight, S.A.; Sunde, R.A. The effect of progressive selenium deficiency on anti-glutathione peroxidase antibody reactive protein in rat liver. J. Nutr. 1987, 117, 732–738. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Oberley, L.W.; Li, Y. A simple method for clinical assay of superoxide dismutase. Clin. Chem. 1988, 34, 497–500. [Google Scholar] [PubMed]
- Aebi, H.; Wyss, S.R.; Scherz, B.; Skvaril, F. Heterogeneity of erythrocyte catalase ii. Isolation and characterization of normal and variant erythrocyte catalase and their subunits. Eur. J. Biochem. 1974, 48, 137–145. [Google Scholar] [CrossRef] [PubMed]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Fowler, S.D.; Greenspan, P. Application of Nile red, a fluorescent hydrophobic probe, for the detection of neutral lipid deposits in tissue sections: Comparison with oil red O. J. Histochem. Cytochem. 1985, 33, 833–836. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; López-Casillas, F.; Bai, D.H.; Luo, X.; Pape, M.E. Role of reversible phosphorylation of acetyl-COA carboxylase in long-chain fatty acid synthesis. FASEB J. 1989, 3, 2250–2256. [Google Scholar] [CrossRef] [PubMed]
- Brandon-Warner, E.; Schrum, L.W.; Schmidt, C.M.; McKillop, I.H. Rodent models of alcoholic liver disease: Of mice and men. Alcohol 2012, 46, 715–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pirola, R.C.; Lieber, C.S. Hypothesis: Energy wastage in alcoholism and drug abuse: Possible role of hepatic microsomal enzymes. AJCN 1976, 29, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Anji, A.; Kumari, M. Supplementing the liquid alcohol diet with chow enhances alcohol intake in C57 BL/6 mice. Drug Alcohol Depend. 2008, 97, 86–93. [Google Scholar] [CrossRef] [PubMed]
- Ji, C.; Chan, C.; Kaplowitz, N. Predominant role of sterol response element binding proteins (SREBP) lipogenic pathways in hepatic steatosis in the murine intragastric ethanol feeding model. J. Hepatol. 2006, 45, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Investig. 2002, 109, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Zeng, T.; Zhang, C.L.; Song, F.Y.; Zhao, X.L.; Xie, K.Q. CMZ Reversed Chronic Ethanol-Induced Disturbance of PPAR-α Possibly by Suppressing Oxidative Stress and PGC-1α Acetylation, and Activating the MAPK and GSK3β Pathway. PLoS ONE 2014, 9, e98658. [Google Scholar] [CrossRef] [PubMed]
- Crabb, D.W.; Galli, A.; Fischer, M.; You, M. Molecular mechanisms of alcoholic fatty liver: Role of peroxisome proliferator-activated receptor alpha. Alcohol 2004, 34, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Fischer, M.; You, M.; Matsumoto, M.; Crabb, D.W. Peroxisome proliferator-activated receptor α (PPARα) agonist treatment reverses PPARα dysfunction and abnormalities in hepatic lipid metabolism in ethanol-fed mice. J. Biol. Chem. 2003, 278, 27997–28004. [Google Scholar] [CrossRef] [PubMed]
- Klop, B.; Do Rego, A.T.; Cabezas, M.C. Alcohol and plasma triglycerides. Curr. Opin. Lipidol. 2013, 24, 321–326. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, E.S.E.R.; Foster, D.; McGee Harper, M.; Seidman, C.E.; Smith, J.D.; Breslow, J.L.; Brinton, E.A. Alcohol consumption raises HDL cholesterol levels by increasing the transport rate of apolipoproteins a-i and a-ii. Circulation 2000, 102, 2347–2352. [Google Scholar] [CrossRef]
- Fan, F.; Cao, Q.; Wang, C.; Ma, X.; Shen, C.; Liu, X.W.; Bu, L.P.; Zou, Y.Z.; Hu, K.; Sun, A.J.; et al. Impact of chronic low to moderate alcohol consumption on blood lipid and heart energy profile in acetaldehyde dehydrogenase 2-deficient mice. Acta. Pharmacol. Sin. 2014, 35, 1015–1022. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ambade, A.; Mandrekar, P. Oxidative stress and inflammation: Essential partners in alcoholic liver disease. Int. J. Hepatol. 2012, 2012, 853175. [Google Scholar] [CrossRef] [PubMed]
- Keshavarzian, A.; Farhadi, A.; Forsyth, C.B.; Rangan, J.; Jakate, S.; Shaikh, M.; Banan, A.; Fields, J.Z. Evidence that chronic alcohol exposure promotes intestinal oxidative stress, intestinal hyperpermeability and endotoxemia prior to development of alcoholic steatohepatitis in rats. J. Hepatol. 2009, 50, 538–547. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, G.L.; Klein, R.D.; Aminlari, A.; Zhang, H.Y.; Steinstraesser, L.; Alarcon, W.H.; Remick, D.G.; Wang, S.C. Kupffer cell activation by lipopolysaccharide in rats: Role for lipopolysaccharide binding protein and toll-like receptor 4. Hepatology 2000, 31, 932–936. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaratani, H.; Tsujimoto, T.; Douhara, A.; Takaya, H.; Moriya, K.; Namisaki, T.; Noguchi, R.; Yoshiji, H.; Fujimoto, M.; Fukui, H. The effect of inflammatory cytokines in alcoholic liver disease. Mediat. Inflamm. 2013, 2013, 495156. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, B.S.; Rodrigues, G.B.; Rocha, S.W.; Ribeiro, E.L.; Gomes, F.O.; E Silva, A.K.; Peixoto, C.A. Inhibition of NF-kB activation by diethylcarbamazine prevents alcohol-induced liver injury in C57BL/6 mice. Tissue Cell 2014, 46, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Bohm, F.; Kohler, U.A.; Speicher, T.; Werner, S. Regulation of liver regeneration by growth factors and cytokines. EMBO Mol. Med. 2010, 2, 294–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Isayama, F.; Froh, M.; Yin, M.; Conzelmann, L.O.; Milton, R.J.; McKim, S.E.; Wheeler, M.D. TNF α-induced ras activation due to ethanol promotes hepatocyte proliferation independently of liver injury in the mouse. Hepatology 2004, 39, 721–731. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Tachibana, S.; Wang, H.; Hisada, M.; Williams, G.M.; Gao, B.; Sun, Z. Interleukin-6 is an important mediator for mitochondrial DNA repair after alcoholic liver injury in mice. Hepatology 2010, 52, 2137–2147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Pandey, A.K. Chemistry and biological activities of flavonoids: An overview. Sci. World J. 2013, 2013, 162750. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Jia, Q.; Wang, Y.; Zhang, Y.; Xia, M. The anthocyanin cyanidin-3-O-β-glucoside, a flavonoid, increases hepatic glutathione synthesis and protects hepatocytes against reactive oxygen species during hyperglycemia: Involvement of a cAMP-PKA-dependent signaling pathway. Free Radic. Biol. Med. 2012, 52, 314–327. [Google Scholar] [CrossRef] [PubMed]
- Ludwig, A.; Lorenz, M.; Grimbo, N.; Steinle, F.; Meiners, S.; Bartsch, C.; Stangl, K.; Baumann, G.; Stangl, V. The tea flavonoid epigallocatechin-3-gallate reduces cytokine-induced VCAM-1 expression and monocyte adhesion to endothelial cells. Biochem. Biophys. Res. Commun. 2004, 316, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Pezeshki, A.; Safi, S.; Feizi, A.; Askari, G.; Karami, F. The effect of green tea extract supplementation on liver enzymes in patients with nonalcoholic fatty liver disease. Int. J. Prev. Med. 2016, 7, 28. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diets Composition | Control Liquid Diet (%) | Alcohol-Containing Liquid Diet (%) |
|---|---|---|
| Maltodextrin | 9.00 | 0.00 |
| Ethanol | 0.00 | 5.10 |
| Carbohydrate | 5.35 | 5.35 |
| Protein | 4.06 | 4.06 |
| Fat | 3.90 | 3.90 |
| Fiber | 0.69 | 0.69 |
| Water | 77.00 | 80.90 |
| Body Weight (g) | ||||
|---|---|---|---|---|
| Group Week | Control | EtOH | EtOH + 1% AEPL | EtOH + 2% AEPL |
| 1st week | 22.5 ± 0.6 a | 22.5 ± 0.8 a | 22.0 ± 0.7 a | 22.0 ± 0.7 a |
| 2nd week | 22.5 ± 0.5 a | 22.5 ± 0.7 a | 22.5 ± 0.8 a | 22.5 ± 0.7 a |
| 3rd week | 23.0 ± 0.6 a | 21.0 ± 0.8 b | 22.5 ± 0.6 a | 22.0 ± 0.9 a |
| 4th week | 23.5 ± 0.9 a | 20.5 ± 0.5 b | 22.0 ± 0.8 a | 22.5 ± 0.9 a |
| 5th week | 25.0 ± 0.7 a | 20.0 ± 0.7 c | 21.5 ± 0.9 b | 23.5 ± 0.8 a |
| Liver weight (g/100 g body weight) | 5.3 ± 0.9 a | 5.5 ± 0.5 ab | 5.4 ± 0.5 ab | 5.6 ± 0.2 ab |
| Groups | Control | EtOH | EtOH + 1% AEPL | EtOH + 2% AEPL |
|---|---|---|---|---|
| AST (U/L) | 169.7 ± 61.2 b | 285.8 ± 21.5 a | 225.0 ± 53.4 ab | 205.0 ± 65.5 b |
| ALT (U/L) | 40.8 ± 6.2 b | 65.3 ± 6.1 a | 51.8 ± 9.4 b | 51.7 ± 8.1 b |
| TC (mg/dL) | 105.7 ± 5.8 b | 134.1 ± 17.7 a | 121.0 ± 9.5 a | 104.6 ± 20.0 b |
| TG (mg/dL) | 67.6 ± 9.4 b | 94.1 ± 16.7 a | 74.8 ± 13.4 b | 73.3 ± 12.1 b |
| HDL (mg/dL) | 49.5 ± 5.6 b | 74.5 ± 11.9 a | 78.0 ± 12.2 a | 69.7 ± 14.5 a |
| LDL (mg/dL) | 40.0 ± 4.6 b | 57.0 ± 7.0 a | 41.7 ± 5.8 b | 45.3 ± 0.6 b |
| VLDL (mg/dL) | 14.8 ± 2.2 b | 19.3 ± 3.0 a | 15.6 ± 2.5 b | 13.3 ± 1.2 b |
| Hepatic TG (µg/mg protein) | 306.3 ± 15.2 b | 388.9 ± 33.7 a | 360.9 ± 35.4 a | 326.4 ± 42.6 b |
| Antioxidant Substances | Control | EtOH | EtOH + 1% AEPL | EtOH + 2% AEPL |
|---|---|---|---|---|
| SOD (U/mg protein) | 1.6 ± 0.3 a | 0.8 ± 0.3 b | 1.3 ± 0.4 a | 1.2 ± 0.4 a |
| Catalase (U/mg protein) | 36.2 ± 7.8 ab | 28.5 ± 4.0 c | 36.7 ± 5.8 b | 45.9 ± 6.2 a |
| GPx (nmol NADPH/min/mg protein) | 32.0 ± 4.4 a | 18.2 ± 4.8 c | 22.3 ± 6.8 bc | 30.1 ± 2.1 ab |
| GSH (nmol/mg protein) | 15.3 ± 2.3 b | 17.7 ± 2.6 b | 17.1 ± 1.2 ab | 20.1 ± 3.1 a |
| TEAC (mg/dL) | 0.18 ± 0.0 a | 0.16 ± 0.0 b | 0.18 ± 0.0 a | 0.19 ± 0.0 a |
| MDA (nmole/mg protein) | 3.2 ± 0.1 b | 8.2 ± 1.0 a | 2.0 ± 1.0 b | 2.0 ± 0.6 b |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hsu, J.-Y.; Lin, H.-H.; Hsu, C.-C.; Chen, B.-C.; Chen, J.-H. Aqueous Extract of Pepino (Solanum muriactum Ait) Leaves Ameliorate Lipid Accumulation and Oxidative Stress in Alcoholic Fatty Liver Disease. Nutrients 2018, 10, 931. https://doi.org/10.3390/nu10070931
Hsu J-Y, Lin H-H, Hsu C-C, Chen B-C, Chen J-H. Aqueous Extract of Pepino (Solanum muriactum Ait) Leaves Ameliorate Lipid Accumulation and Oxidative Stress in Alcoholic Fatty Liver Disease. Nutrients. 2018; 10(7):931. https://doi.org/10.3390/nu10070931
Chicago/Turabian StyleHsu, Jen-Ying, Hui-Hsuan Lin, Cheng-Chin Hsu, Bing-Chen Chen, and Jing-Hsien Chen. 2018. "Aqueous Extract of Pepino (Solanum muriactum Ait) Leaves Ameliorate Lipid Accumulation and Oxidative Stress in Alcoholic Fatty Liver Disease" Nutrients 10, no. 7: 931. https://doi.org/10.3390/nu10070931
APA StyleHsu, J.-Y., Lin, H.-H., Hsu, C.-C., Chen, B.-C., & Chen, J.-H. (2018). Aqueous Extract of Pepino (Solanum muriactum Ait) Leaves Ameliorate Lipid Accumulation and Oxidative Stress in Alcoholic Fatty Liver Disease. Nutrients, 10(7), 931. https://doi.org/10.3390/nu10070931