Pilot Study Examining the Influence of Potassium Bicarbonate Supplementation on Nitrogen Balance and Whole-Body Ammonia and Urea Turnover Following Short-Term Energy Restriction in Older Men

Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Study Design
2.3. Anthropometrics
2.4. Study Diet
2.5. KHCO3 Supplement
2.6. Net Acid Excretion
2.7. Nitrogen Balance
2.8. Whole-Body Ammonia and Urea Turnover
2.9. Statistical Analysis
3. Results
3.1. Participants, Body Mass, Pill Compliance, and Dietary Intake
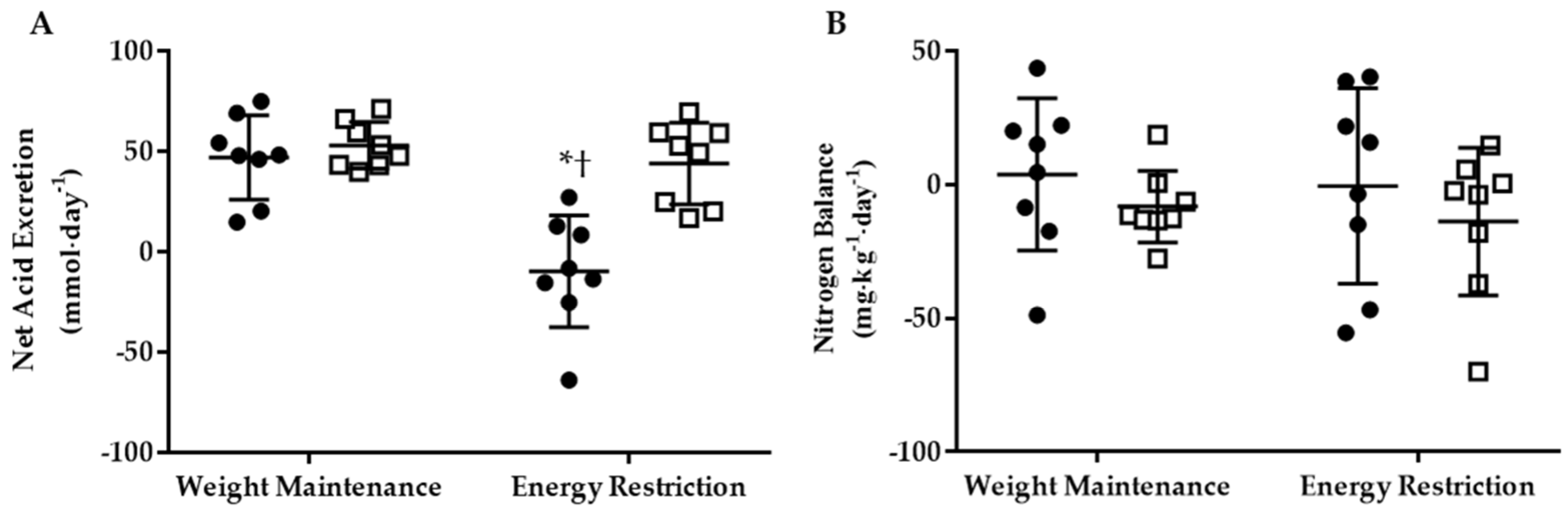
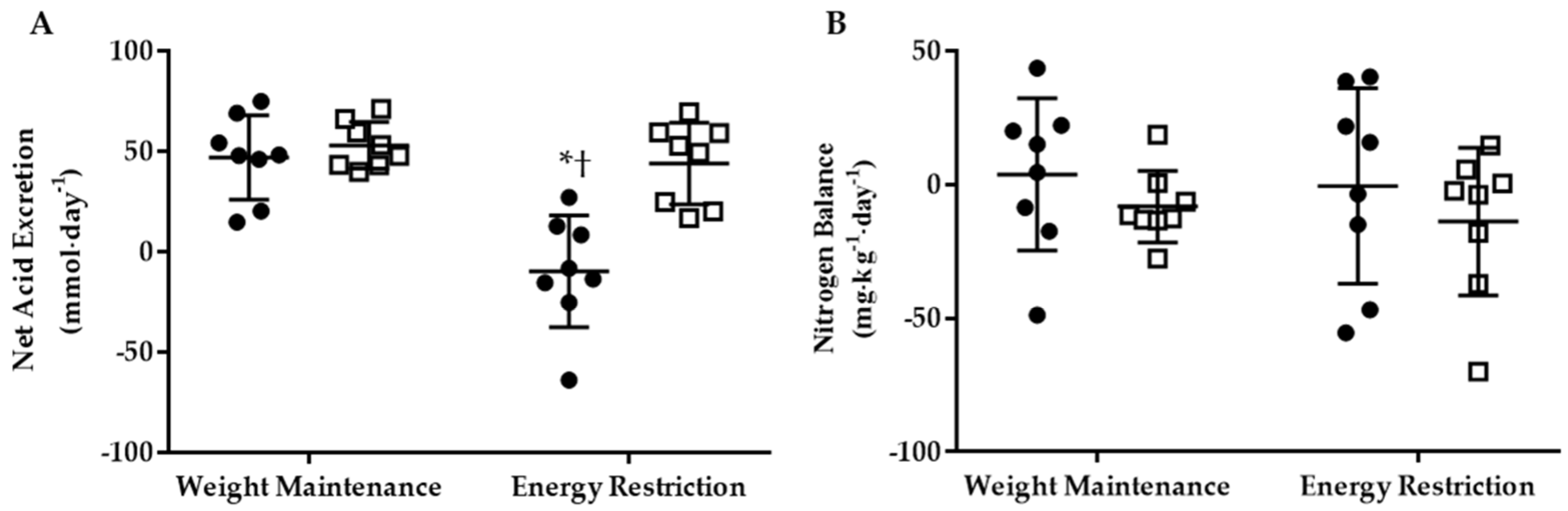
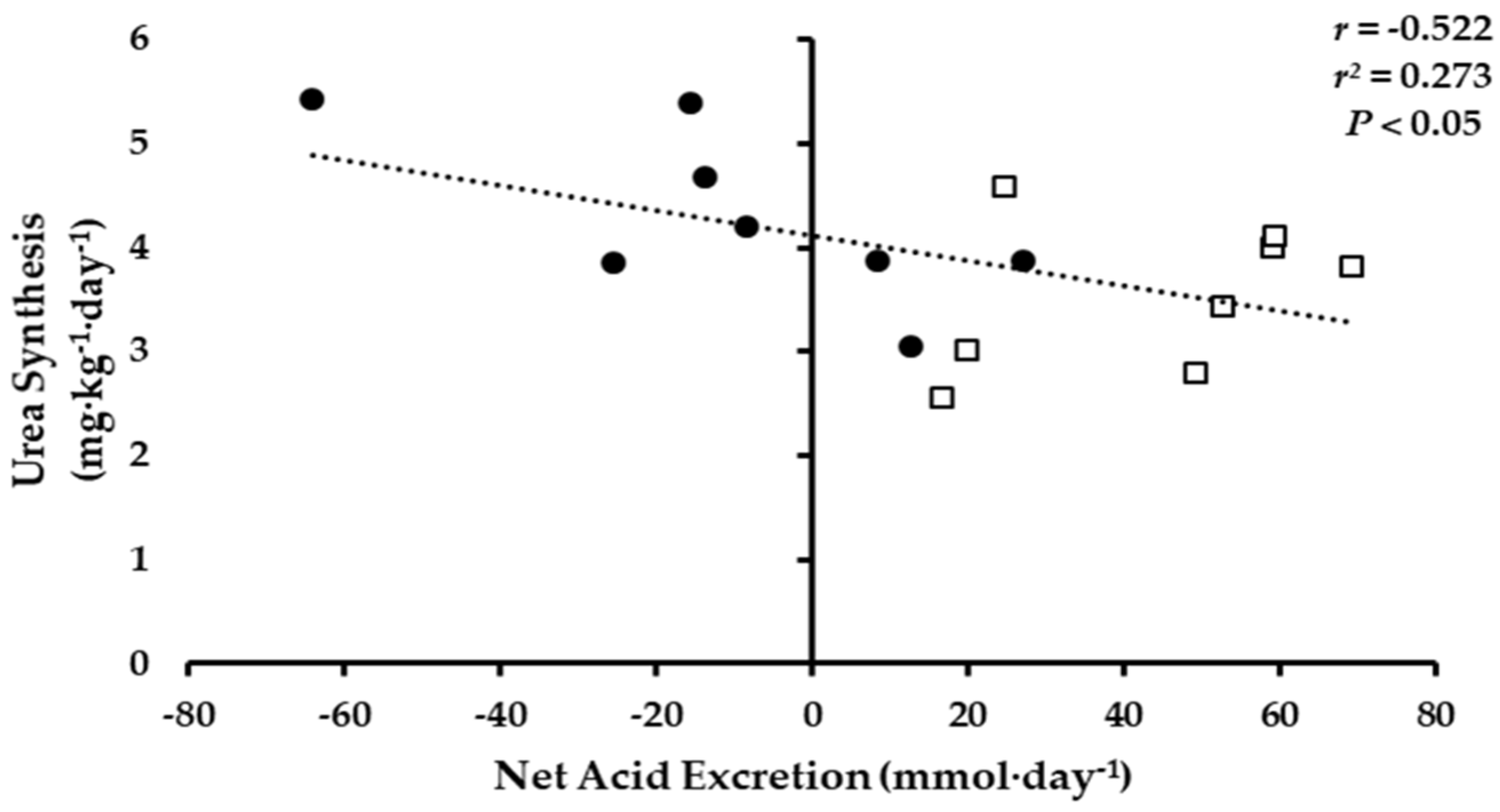
3.2. Net Acid Excretion and Nitrogen Balance
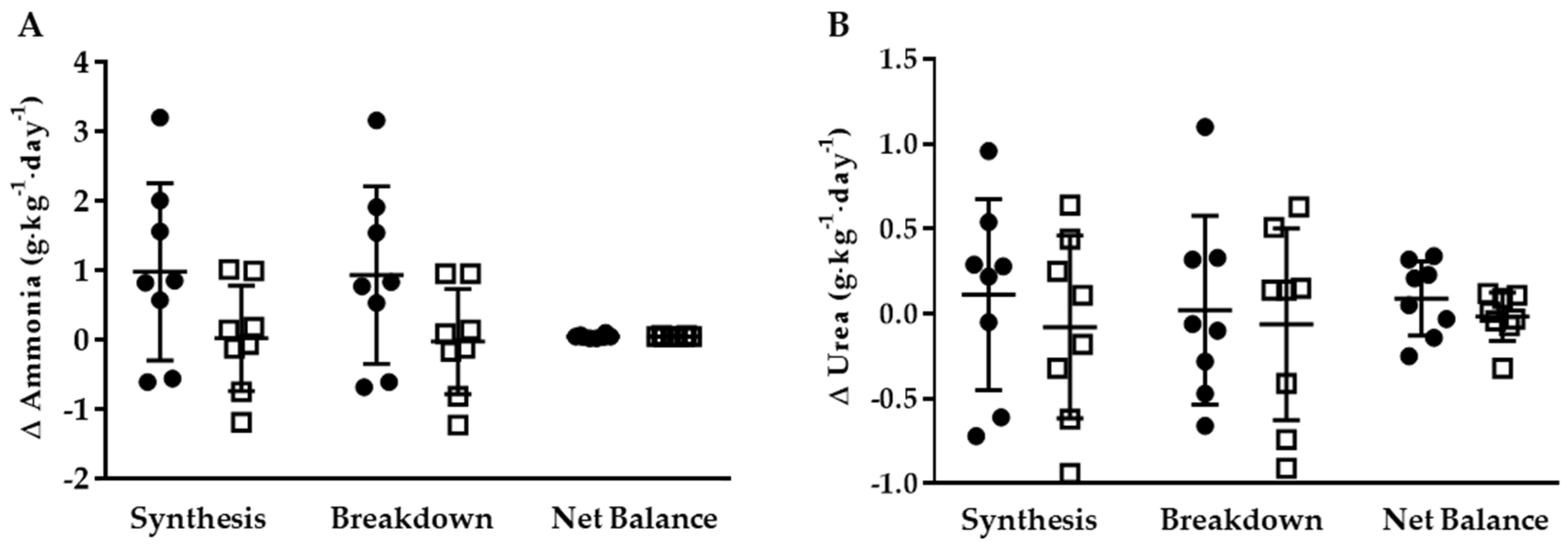
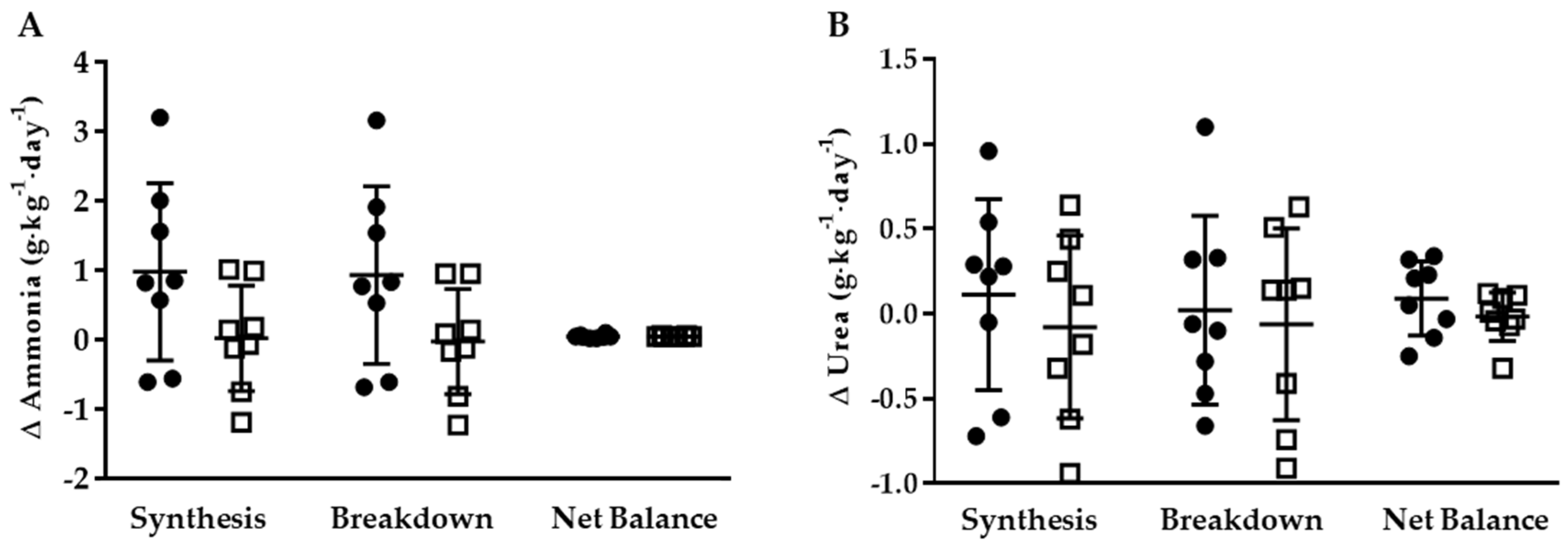
3.3. Whole-Body Ammonia and Urea Turnover
3.3.1. KHCO3 vs. PLA
3.3.2. Energy Restriction
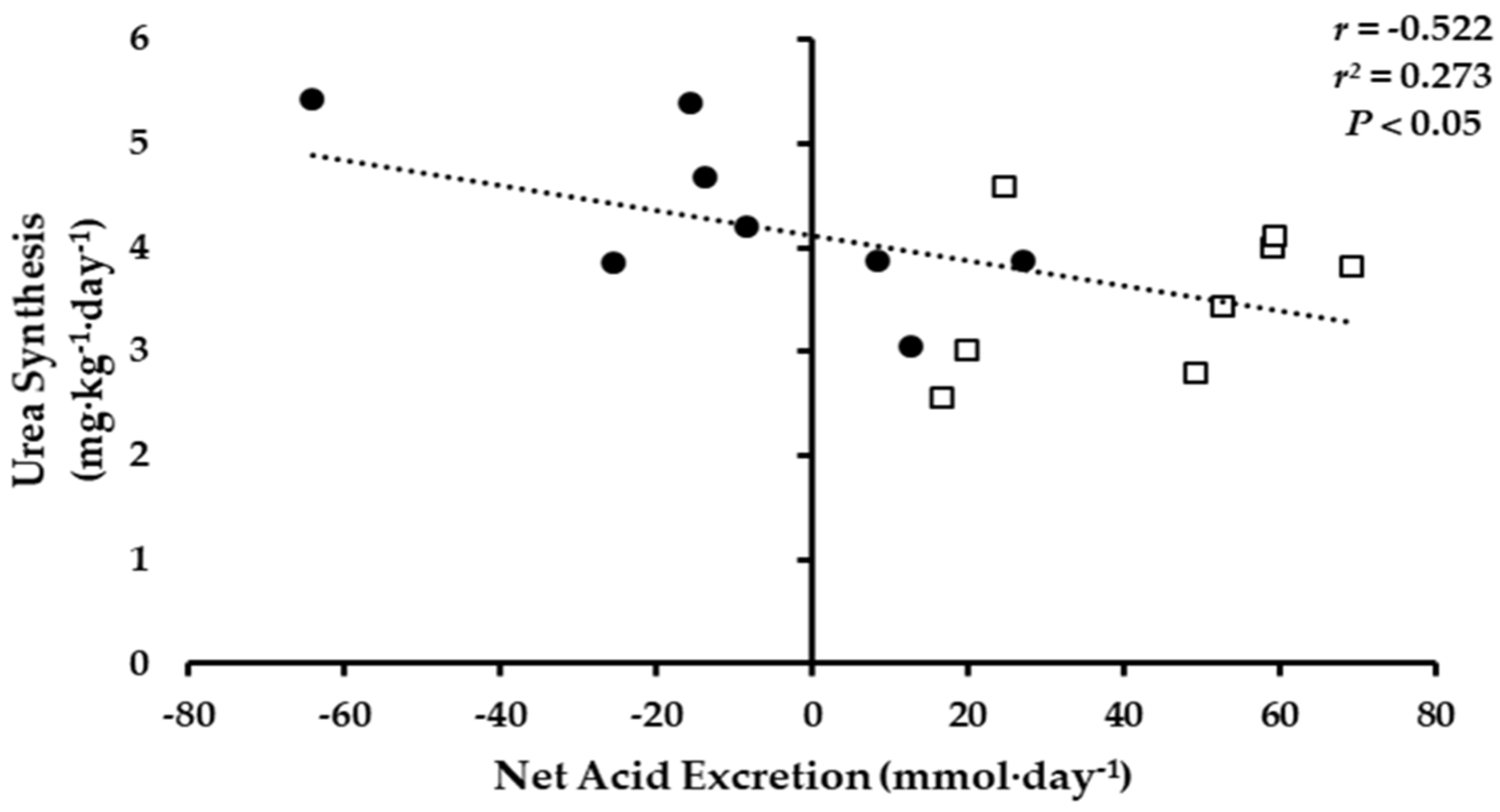
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Finkelstein, E.A.; Trogdon, J.G.; Cohen, J.W.; Dietz, W. Annual medical spending attributable to obesity: Payer-and service-specific estimates. Health Aff. 2009, 28, w822–w831. [Google Scholar] [CrossRef] [PubMed]
- Flegal, K.M.; Carroll, M.D.; Ogden, C.L.; Curtin, L.R. Prevalence and trends in obesity among us adults, 1999–2008. JAMA 2010, 303, 235–241. [Google Scholar] [CrossRef] [PubMed]
- Cawley, J.; Meyerhoefer, C. The medical care costs of obesity: An instrumental variables approach. J. Health Econ. 2012, 31, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, G. Effect of degree of weight loss on health benefits. Obes. Res. 1995, 3 (Suppl. 2), 211s–216s. [Google Scholar] [CrossRef] [PubMed]
- Pasanisi, F.; Contaldo, F.; de Simone, G.; Mancini, M. Benefits of sustained moderate weight loss in obesity. Nutr. Metab. Cardiovasc. Dis. 2001, 11, 401–406. [Google Scholar] [PubMed]
- Starr, K.N.P.; McDonald, S.R.; Bales, C.W. Obesity and physical frailty in older adults: A scoping review of lifestyle intervention trials. J. Am. Med. Dir. Assoc. 2014, 15, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Weinheimer, E.M.; Sands, L.P.; Campbell, W.W. A systematic review of the separate and combined effects of energy restriction and exercise on fat-free mass in middle-aged and older adults: Implications for sarcopenic obesity. Nutr. Rev. 2010, 68, 375–388. [Google Scholar] [CrossRef] [PubMed]
- Fielding, R.A.; Vellas, B.; Evans, W.J.; Bhasin, S.; Morley, J.E.; Newman, A.B.; van Kan, G.A.; Andrieu, S.; Bauer, J.; Breuille, D.; et al. Sarcopenia: An undiagnosed condition in older adults. Current consensus definition: Prevalence, etiology, and consequences. International working group on sarcopenia. J. Am. Med. Dir. Assoc. 2011, 12, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Waters, D.L.; Ward, A.L.; Villareal, D.T. Weight loss in obese adults 65 years and older: A review of the controversy. Exp. Gerontol. 2013, 48, 1054–1061. [Google Scholar] [CrossRef] [PubMed]
- Frassetto, L.; Sebastian, A. Age and systemic acid-base equilibrium: Analysis of published data. J. Gerontol. A Biol. Sci. Med. Sci. 1996, 51, B91–B99. [Google Scholar] [CrossRef] [PubMed]
- Caso, G.; Garlick, P.J. Control of muscle protein kinetics by acid-base balance. Curr. Opin. Clin. Nutr. Metab. Care 2005, 8, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Dawson-Hughes, B.; Harris, S.S.; Ceglia, L. Alkaline diets favor lean tissue mass in older adults. Am. J. Clin. Nutr. 2008, 87, 662–665. [Google Scholar] [CrossRef] [PubMed]
- Ballmer, P.E.; McNurlan, M.A.; Hulter, H.N.; Anderson, S.E.; Garlick, P.J.; Krapf, R. Chronic metabolic acidosis decreases albumin synthesis and induces negative nitrogen balance in humans. J. Clin. Investig. 1995, 95, 39–45. [Google Scholar] [CrossRef] [PubMed]
- Frassetto, L.; Morris, R.C., Jr.; Sebastian, A. Potassium bicarbonate reduces urinary nitrogen excretion in postmenopausal women. J. Clin. Endocrinol. Metab. 1997, 82, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Dawson-Hughes, B.; Harris, S.S.; Palermo, N.J.; Castaneda-Sceppa, C.; Rasmussen, H.M.; Dallal, G.E. Treatment with potassium bicarbonate lowers calcium excretion and bone resorption in older men and women. J. Clin. Endocrinol. Metab. 2009, 94, 96–102. [Google Scholar] [CrossRef] [PubMed]
- Ceglia, L.; Harris, S.S.; Abrams, S.A.; Rasmussen, H.M.; Dallal, G.E.; Dawson-Hughes, B. Potassium bicarbonate attenuates the urinary nitrogen excretion that accompanies an increase in dietary protein and may promote calcium absorption. J. Clin. Endocrinol. Metab. 2009, 94, 645–653. [Google Scholar] [CrossRef] [PubMed]
- Ceglia, L.; Dawson-Hughes, B. Increasing alkali supplementation decreases urinary nitrogen excretion when adjusted for same day nitrogen intake. Osteoporos. Int. 2017, 28, 3355–3359. [Google Scholar] [CrossRef] [PubMed]
- Pasiakos, S.M.; Cao, J.J.; Margolis, L.M.; Sauter, E.R.; Whigham, L.D.; McClung, J.P.; Rood, J.C.; Carbone, J.W.; Combs, G.F., Jr.; Young, A.J. Effects of high-protein diets on fat-free mass and muscle protein synthesis following weight loss: A randomized controlled trial. FASEB J. 2013, 27, 3837–3847. [Google Scholar] [CrossRef] [PubMed]
- Bauer, J.; Biolo, G.; Cederholm, T.; Cesari, M.; Cruz-Jentoft, A.J.; Morley, J.E.; Phillips, S.; Sieber, C.; Stehle, P.; Teta, D.; et al. Evidence-based recommendations for optimal dietary protein intake in older people: A position paper from the prot-age study group. J. Am. Med. Dir. Assoc. 2013, 14, 542–559. [Google Scholar] [CrossRef] [PubMed]
- Remer, T.; Manz, F. Potential renal acid load of foods and its influence on urine ph. J. Am. Diet. Assoc. 1995, 95, 791–797. [Google Scholar] [CrossRef]
- Jorgensen, K. Titrimetric determination of the net excretion of acid/base in urine. Scand. J. Clin. Lab. Investig. 1957, 9, 287–291. [Google Scholar] [CrossRef]
- Chan, J.C. The rapid determination of urinary titratable acid and ammonium and evaluation of freezing as a method of preservation. Clin. Biochem. 1972, 5, 94–98. [Google Scholar] [CrossRef]
- Friedman, J.E.; Lemon, P.W. Effect of chronic endurance exercise on retention of dietary protein. Int. J. Sports Med. 1989, 10, 118–123. [Google Scholar] [CrossRef] [PubMed]
- Duggleby, S.L.; Waterlow, J.C. The end-product method of measuring whole-body protein turnover: A review of published results and a comparison with those obtained by leucine infusion. Br. J. Nutr. 2005, 94, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Guder, W.G.; Haussinger, D.; Gerok, W. Renal and hepatic nitrogen metabolism in systemic acid base regulation. J. Clin. Chem. Clin. Biochem. 1987, 25, 457–466. [Google Scholar] [CrossRef] [PubMed]
- Vazquez, J.A.; Kazi, U.; Madani, N. Protein metabolism during weight reduction with very-low-energy diets: Evaluation of the independent effects of protein and carbohydrate on protein sparing. Am. J. Clin. Nutr. 1995, 62, 93–103. [Google Scholar] [PubMed]
- Weiner, I.D.; Mitch, W.E.; Sands, J.M. Urea and ammonia metabolism and the control of renal nitrogen excretion. Clin. J. Am. Soc. Nephrol. 2015, 10, 1444–1458. [Google Scholar] [CrossRef] [PubMed]
- Hartman, J.W.; Moore, D.R.; Phillips, S.M. Resistance training reduces whole-body protein turnover and improves net protein retention in untrained young males. Appl. Physiol. Nutr. Metab. 2006, 31, 557–564. [Google Scholar] [CrossRef] [PubMed]
- Matthews, D.E.; Conway, J.M.; Young, V.R.; Bier, D.M. Glycine nitrogen metabolism in man. Metabolism 1981, 30, 886–893. [Google Scholar] [CrossRef]
- Wilkinson, D.J. Historical and contemporary stable isotope tracer approaches to studying mammalian protein metabolism. Mass Spectrom. Rev. 2018, 37, 57–80. [Google Scholar] [CrossRef] [PubMed]
- Tareen, N.; Zadshir, A.; Martins, D.; Nagami, G.; Levine, B.; Norris, K.C. Alterations in acid-base homeostasis with aging. J. Natl. Med. Assoc. 2004, 96, 921–925; quiz 925–926. [Google Scholar] [PubMed]
- Kleger, G.R.; Turgay, M.; Imoberdorf, R.; McNurlan, M.A.; Garlick, P.J.; Ballmer, P.E. Acute metabolic acidosis decreases muscle protein synthesis but not albumin synthesis in humans. Am. J. Kidney Dis. 2001, 38, 1199–1207. [Google Scholar] [CrossRef] [PubMed]
- Reaich, D.; Channon, S.M.; Scrimgeour, C.M.; Goodship, T.H. Ammonium chloride-induced acidosis increases protein breakdown and amino acid oxidation in humans. Am. J. Physiol. 1992, 263, E735–E739. [Google Scholar] [CrossRef] [PubMed]
- Ceglia, L.; Rivas, D.A.; Pojednic, R.M.; Price, L.L.; Harris, S.S.; Smith, D.; Fielding, R.A.; Dawson-Hughes, B. Effects of alkali supplementation and vitamin d insufficiency on rat skeletal muscle. Endocrine 2013, 44, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Todd, K.S.; Butterfield, G.E.; Calloway, D.H. Nitrogen balance in men with adequate and deficient energy intake at three levels of work. J. Nutr. 1984, 114, 2107–2118. [Google Scholar] [CrossRef] [PubMed]
- Calloway, D.H. Nitrogen balance of men with marginal intakes of protein and energy. J. Nutr. 1975, 105, 914–923. [Google Scholar] [CrossRef] [PubMed]
- Garlick, P.J.; McNurlan, M.A.; Ballmer, P.E. Influence of dietary protein intake on whole-body protein turnover in humans. Diabetes Care 1991, 14, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Waterlow, J.C. Protein turnover with special reference to man. Q. J. Exp. Physiol. 1984, 69, 409–438. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, R.R.; Chinkes, D.L. Isotope Tracers in Metabolic Research: Principles and Practice of Kinetic Analysis; John Wiley & Sons: Hoboken, NJ, USA, 2005. [Google Scholar]
- Noordzij, M.; Tripepi, G.; Dekker, F.W.; Zoccali, C.; Tanck, M.W.; Jager, K.J. Sample size calculations: Basic principles and common pitfalls. Nephrol. Dial. Transplant. 2010, 25, 1388–1393. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
| Weight Maintenance | Energy Restriction | Delta | ||||
|---|---|---|---|---|---|---|
| KHCO3 | PLA | KHCO3 | PLA | KHCO3 | PLA | |
| Body Mass (g) | 89.9 ± 6.8 | 87.7 ± 10.5 | 85.1 ± 7.4 * | 83.8 ± 10.0 * | −4.8 ± 1.6 | −3.9 ± 0.9 |
| BMI (kg·m−2) | 28.9 ± 1.3 | 28.0 ± 2.5 | 27.3 ± 1.6 * | 26.8 ± 2.4 * | −1.6 ± 0.5 | −1.2 ± 0.3 |
| Weight Maintenance | Energy Restriction | |||
|---|---|---|---|---|
| KHCO3 | PLA | KHCO3 | PLA | |
| Energy (kcal·day−1) | 2642 ± 244 | 2580 ± 296 | 1818 ± 231 * | 1802 ± 189 * |
| Protein (g·day−1) | 90 ± 7 | 89 ± 10 | 89 ± 7 | 88 ± 10 |
| Carbohydrate (g·day−1) | 409 ± 42 | 398 ± 47 | 257 ± 44 * | 257 ± 26 * |
| Fat (g·day−1) | 78 ± 7 | 76 ± 8 | 51 ± 6 * | 50 ± 5 * |
| PRAL (mEq·day−1) 2 | 25 ± 5 | 24 ± 6 | 27 ± 5 * | 26 ± 5 * |
| Weight Maintenance | Energy Restriction | |||
|---|---|---|---|---|
| KHCO3 | PLA | KHCO3 | PLA | |
| Ammonia (g∙kg−1∙day−1) | ||||
| Synthesis | 3.78 ± 1.00 | 3.46 ± 0.91 | 4.76 ± 1.20 | 3.48 ± 1.25 |
| Breakdown | 2.79 ± 1.00 | 2.48 ± 0.91 | 3.72 ± 1.21 | 2.45 ± 1.25 |
| Net Balance | 0.99 ± 0.22 | 0.98 ± 0.09 | 1.04 ± 0.03 * | 1.03 ± 0.12 * |
| Urea (g∙kg−1∙day−1) | ||||
| Synthesis | 4.17 ± 0.98 | 3.61 ± 0.55 | 4.28 ± 0.82 | 3.53 ± 0.71 |
| Breakdown | 3.91 ± 0.90 | 3.25 ± 0.54 | 3.93 ± 0.79 | 3.19 ± 0.75 |
| Net Balance | 0.26 ± 0.19 | 0.36 ± 0.8 | 0.35 ± 0.20 | 0.34 ± 0.14 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Margolis, L.M.; Ceglia, L.; Rivas, D.A.; Dawson-Hughes, B.; Fielding, R.A. Pilot Study Examining the Influence of Potassium Bicarbonate Supplementation on Nitrogen Balance and Whole-Body Ammonia and Urea Turnover Following Short-Term Energy Restriction in Older Men. Nutrients 2018, 10, 624. https://doi.org/10.3390/nu10050624
Margolis LM, Ceglia L, Rivas DA, Dawson-Hughes B, Fielding RA. Pilot Study Examining the Influence of Potassium Bicarbonate Supplementation on Nitrogen Balance and Whole-Body Ammonia and Urea Turnover Following Short-Term Energy Restriction in Older Men. Nutrients. 2018; 10(5):624. https://doi.org/10.3390/nu10050624
Chicago/Turabian StyleMargolis, Lee M., Lisa Ceglia, Donato A. Rivas, Bess Dawson-Hughes, and Roger A. Fielding. 2018. "Pilot Study Examining the Influence of Potassium Bicarbonate Supplementation on Nitrogen Balance and Whole-Body Ammonia and Urea Turnover Following Short-Term Energy Restriction in Older Men" Nutrients 10, no. 5: 624. https://doi.org/10.3390/nu10050624
APA StyleMargolis, L. M., Ceglia, L., Rivas, D. A., Dawson-Hughes, B., & Fielding, R. A. (2018). Pilot Study Examining the Influence of Potassium Bicarbonate Supplementation on Nitrogen Balance and Whole-Body Ammonia and Urea Turnover Following Short-Term Energy Restriction in Older Men. Nutrients, 10(5), 624. https://doi.org/10.3390/nu10050624