Anthropometric and Dietary Factors as Predictors of DNA Damage in Obese Women




Abstract
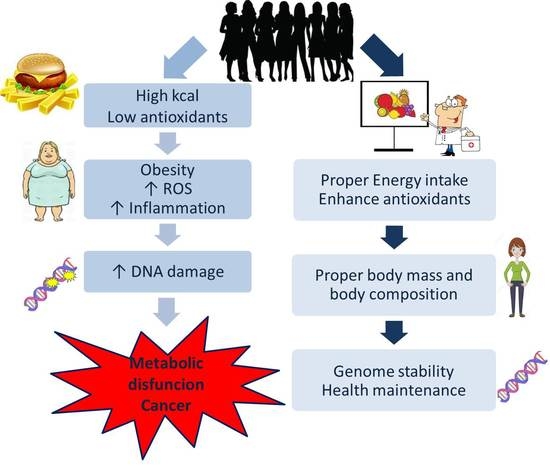
1. Introduction
2. Materials and Methods
2.1. Study Participants
2.2. Anthropometric Measurements
2.3. Dietary Intake Assessment
2.4. Blood Analyses
2.5. Comet Assay
2.6. Statistical Analysis
3. Results
4. Discussion
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| CRP | C-reactive protein |
| WHR | waist-hip-ratio |
| ROS | reactive oxygen species |
| SFAs | saturated fatty acids |
| NF-kB | Nuclear factor kappa B |
| 8-oxodG | 8-Oxo-2′-deoxyguanosine |
| 8-oxoGua | 8-Oxoguanine |
References
- Chatterjee, N.; Walker, G.C. Mechanisms of DNA damage, repair, and mutagenesis. Environ. Mol. Mutagen. 2017, 58, 235–263. [Google Scholar] [CrossRef] [PubMed]
- Schmutte, C.; Fishel, R. Genomic instability: First step to carcinogenesis. Anticancer Res. 1999, 19, 4665–4696. [Google Scholar] [PubMed]
- Collins, A.R. The comet assay for DNA damage and repair: Principles, applications, and limitations. Mol. Biotechnol. 2004, 26, 249–261. [Google Scholar] [CrossRef]
- Bryant, P.E. DNA damage, repair and chromosomal damage. Int. J. Radiat. Biol. 1997, 71, 675–680. [Google Scholar] [CrossRef] [PubMed]
- Kasai, H.; Kawai, K. Oxidative DNA damage: Mechanisms and significance in health and disease. Antioxid. Redox Signal. 2006, 8, 981–983. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Otin, C.; Blasco, M.A.; Partridge, L.; Serrano, M.; Kroemer, G. The hallmarks of aging. Cell 2013, 153, 1194–1217. [Google Scholar] [CrossRef] [PubMed]
- Cerda, C.; Sanchez, C.; Climent, B.; Vazquez, A.; Iradi, A.; El Amrani, F.; Bediaga, A.; Saez, G.T. Oxidative stress and DNA damage in obesity-related tumorigenesis. Adv. Exp. Med. Biol. 2014, 824, 5–17. [Google Scholar] [PubMed]
- Navarro, E.; Funtikova, A.N.; Fito, M.; Schroder, H. Can metabolically healthy obesity be explained. Mol. Nutr. Food Res. 2014. [Google Scholar] [CrossRef]
- Mandviwala, T.; Khalid, U.; Deswal, A. Obesity and Cardiovascular Disease: A Risk Factor or a Risk Marker? Curr. Atheroscler. Rep. 2016, 18, 21. [Google Scholar] [CrossRef] [PubMed]
- Shimizu, I.; Yoshida, Y.; Suda, M.; Minamino, T. DNA damage response and metabolic disease. Cell Metab. 2014, 20, 967–977. [Google Scholar] [CrossRef] [PubMed]
- Puglisi, M.J.; Fernandez, M.L. Modulation of C-reactive protein, tumor necrosis factor-alpha, and adiponectin by diet, exercise, and weight loss. J. Nutr. 2008, 138, 2293–2296. [Google Scholar] [CrossRef] [PubMed]
- Nemzer, B.; Chang, T.; Xie, Z.; Pietrzkowski, Z.; Reyes, T.; Ou, B. Decrease of free radical concentrations in humans following consumption of a high antioxidant capacity natural product. Food Sci. Nutr. 2014, 2, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Riso, P.; Pinder, A.; Santangelo, A.; Porrini, M. Does tomato consumption effectively increase the resistance of lymphocyte DNA to oxidative damage? Am. J. Clin. Nutr. 1999, 69, 712–718. [Google Scholar] [CrossRef] [PubMed]
- Esmaillzadeh, A.; Kimiagar, M.; Mehrabi, Y.; Azadbakht, L.; Hu, F.B.; Willett, W.C. Fruit and vegetable intakes, C-reactive protein, and the metabolic syndrome. Am. J. Clin. Nutr. 2006, 84, 1489–1497. [Google Scholar] [CrossRef] [PubMed]
- Boeing, H.; Dietrich, T.; Hoffmann, K.; Pischon, T.; Ferrari, P.; Lahmann, P.H.; Boutron-Ruault, M.C.; Clavel-Chapelon, F.; Allen, N.; Key, T.; et al. Intake of fruits and vegetables and risk of cancer of the upper aero-digestive tract: The prospective EPIC-study. Cancer Causes Control. 2006, 17, 957–969. [Google Scholar] [CrossRef] [PubMed]
- Mullner, E.; Brath, H.; Pleifer, S.; Schiermayr, C.; Baierl, A.; Wallner, M.; Fastian, T.; Millner, Y.; Paller, K.; Henriksen, T.; et al. Vegetables and PUFA-rich plant oil reduce DNA strand breaks in individuals with type 2 diabetes. Mol. Nutr. Food Res. 2013, 57, 328–338. [Google Scholar] [CrossRef] [PubMed]
- Kimura, Y.; Sato, M.; Kurotani, K.; Nanri, A.; Kawai, K.; Kasai, H.; Imaizumi, K.; Mizoue, T. PUFAs in serum cholesterol ester and oxidative DNA damage in Japanese men and women. Am. J. Clin. Nutr. 2012, 95, 1209–1214. [Google Scholar] [CrossRef] [PubMed]
- Cosgrove, J.P.; Church, D.F.; Pryor, W.A. The kinetics of the autoxidation of polyunsaturated fatty acids. Lipids 1987, 22, 299–304. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Wu, G.Z.; Goh, K.J.; Lee, Y.M.; Ng, C.C.; You, A.B.; Wang, J.; Jia, D.; Hao, A.; Yu, Q.; et al. Saturated fatty acids modulate cell response to DNA damage: Implication for their role in tumorigenesis. PLoS ONE 2008, 3, e2329. [Google Scholar] [CrossRef] [PubMed]
- Jablonowska-Lietz, B.; Wrzosek, M.; Wlodarczyk, M.; Nowicka, G. New indexes of body fat distribution, visceral adiposity index, body adiposity index, waist-to-height ratio, and metabolic disturbances in the obese. Kardiol. Pol. 2017, 75, 1185–1191. [Google Scholar] [PubMed]
- World Health Organization. Obesity: Preventing and Managing the Global Epidemic: Report of a WHO Consultation; WHO Technical Report 894; World Health Organization: Geneva, Switzerland, 1999. [Google Scholar]
- Kunachowicz, H.N.I.; Przygoda, B.; Iwanow, K. Tables of Composition and Nutritional Value of Food; PZWL: Warsaw, Poland, 2005. (In Polish) [Google Scholar]
- Jarosz, M.B.-J.B. Standards of Human Nutrition; PZWL: Warsaw, Poland, 2012. (In Polish) [Google Scholar]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low-density lipoprotein cholesterol in plasma, without use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [PubMed]
- Matthews, D.R.; Hosker, J.P.; Rudenski, A.S.; Naylor, B.A.; Treacher, D.F.; Turner, R.C. Homeostasis model assessment: Insulin resistance and beta-cell function from fasting plasma glucose and insulin concentrations in man. Diabetologia 1985, 28, 412–419. [Google Scholar] [CrossRef] [PubMed]
- Singh, N.P.; McCoy, M.T.; Tice, R.R.; Schneider, E.L. A simple technique for quantitation of low levels of DNA damage in individual cells. Exp. Cell Res. 1988, 175, 184–191. [Google Scholar] [CrossRef]
- Wlodarczyk, M.; Nowicka, G. XPD gene rs13181 polymorphism and DNA damage in human lymphocytes. Biochem. Genet. 2012, 50, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Dusinska, M.; Collins, A.R. The comet assay in human biomonitoring: Gene-environment interactions. Mutagenesis 2008, 23, 191–205. [Google Scholar] [CrossRef] [PubMed]
- Hoelzl, C.; Knasmuller, S.; Misik, M.; Collins, A.; Dusinska, M.; Nersesyan, A. Use of single cell gel electrophoresis assays for the detection of DNA-protective effects of dietary factors in humans: Recent results and trends. Mutat. Res. 2009, 681, 68–79. [Google Scholar] [CrossRef] [PubMed]
- Valverde, M.; Rojas, E. Environmental and occupational biomonitoring using the Comet assay. Mutat. Res. 2009, 681, 93–109. [Google Scholar] [CrossRef] [PubMed]
- McKenna, D.J.; McKeown, S.R.; Kelvey-Martin, V.J. Potential use of the comet assay in the clinical management of cancer. Mutagenesis 2008, 23, 183–190. [Google Scholar] [CrossRef] [PubMed]
- Wiencke, J.K.; Kelsey, K.T.; Varkonyi, A.; Semey, K.; Wain, J.C.; Mark, E.; Christiani, D.C. Correlation of DNA adducts in blood mononuclear cells with tobacco carcinogen-induced damage in human lung. Cancer Res. 1995, 55, 4910–4914. [Google Scholar] [PubMed]
- Lenton, K.J.; Therriault, H.; Fulop, T.; Payette, H.; Wagner, J.R. Glutathione and ascorbate are negatively correlated with oxidative DNA damage in human lymphocytes. Carcinogenesis 1999, 20, 607–613. [Google Scholar] [CrossRef] [PubMed]
- Weeden, C.E.; Asselin-Labat, M.L. Mechanisms of DNA damage repair in adult stem cells and implications for cancer formation. Biochim. Biophys. Acta 2018, 1864, 89–101. [Google Scholar] [CrossRef] [PubMed]
- Sestakova, Z.; Kalavska, K.; Hurbanova, L.; Jurkovicova, D.; Gursky, J.; Chovanec, M.; Svetlovska, D.; Miskovska, V.; Obertova, J.; Palacka, P.; et al. The prognostic value of DNA damage level in peripheral blood lymphocytes of chemotherapy-naïve patients with germ cell cancer. Oncotarget 2016, 15, 75996–76005. [Google Scholar]
- Mahmoudi, M.; Gorenne, I.; Mercer, J.; Figg, N.; Littlewood, T.; Bennett, M. Statins use a novel Nijmegen breakage syndrome-1-dependent pathway to accelerate DNA repair in vascular smooth muscle cells. Circ. Res. 2008, 103, 717–725. [Google Scholar] [CrossRef] [PubMed]
- Gray, K.; Kumar, S.; Figg, N.; Harrison, J.; Baker, L.; Mercer, J.; Littlewood, T.; Bennett, M. Effects of DNA damage in smooth muscle cells in atherosclerosis. Circ. Res. 2015, 27, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Cervelli, T.; Borghini, A.; Galli, A.; Andreassi, M.G. DNA damage and repair in atherosclerosis: Current insights and future perspectives. Int. J. Mol. Sci. 2012, 11, 16929–16944. [Google Scholar] [CrossRef] [PubMed]
- Gladyshev, V.N. The free radical theory of aging is dead. Long live the damage theory! Antioxid. Redox Signal. 2014, 20, 727–731. [Google Scholar] [CrossRef] [PubMed]
- Gorbunova, V.; Seluanov, A. DNA double strand break repair, aging and the chromatin connection. Mutat. Res. 2016, 788, 2–6. [Google Scholar] [CrossRef] [PubMed]
- Manna, P.; Jain, S.K. Obesity, Oxidative Stress, Adipose Tissue Dysfunction, and the Associated Health Risks: Causes and Therapeutic Strategies. Metab. Syndr. Relat. Disord. 2015, 13, 423–444. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.C.; Chan, J.C. Evidence for DNA damage as a biological link between diabetes and cancer. Chin. Med. J. 2015, 128, 1543–1548. [Google Scholar] [PubMed]
- Al-Aubaidy, H.A.; Jelinek, H.F. Oxidative DNA damage and obesity in type 2 diabetes mellitus. Eur. J. Endocrinol. 2011, 164, 899–904. [Google Scholar] [CrossRef] [PubMed]
- Okamoto, T.; Kohno, M.; Ito, K.; Takada, K.; Katsura, M.; Morodomi, Y.; Toyokawa, G.; Shoji, F.; Maehara, Y. Clinical Significance of DNA Damage Response Factors and Chromosomal Instability in Primary Lung Adenocarcinoma. Anticancer Res. 2017, 37, 1729–1735. [Google Scholar] [PubMed]
- Herbig, U.; Ferreira, M.; Condel, L.; Carey, D.; Sedivy, J.M. Cellular senescence in aging primates. Science 2006, 311, 1257. [Google Scholar] [CrossRef] [PubMed]
- Freund, A.; Orjalo, A.V.; Desprez, P.Y.; Campisi, J. Inflammatory networks during cellular senescence: Causes and consequences. Trends Mol. Med. 2010, 16, 238–246. [Google Scholar] [CrossRef] [PubMed]
- Rodier, F.; Coppe, J.P.; Patil, C.K.; Hoeijmakers, W.A.; Munoz, D.P.; Raza, S.R.; Freund, A.; Campeau, E.; Davalos, A.R.; Campisi, J. Persistent DNA damage signalling triggers senescence-associated inflammatory cytokine secretion. Nat. Cell Biol. 2009, 11, 973–979. [Google Scholar] [CrossRef] [PubMed]
- Tchkonia, T.; Zhu, Y.; van Deursen, J.; Campisi, J.; Kirkland, J.L. Cellular senescence and the senescent secretory phenotype: Therapeutic opportunities. J. Clin. Investig. 2013, 123, 966–972. [Google Scholar] [CrossRef] [PubMed]
- Hotamisligil, G.S. Inflammation and metabolic disorders. Nature 2006, 444, 860–867. [Google Scholar] [CrossRef] [PubMed]
- Stewart, S.A.; Weinberg, R.A. Telomeres: Cancer to human aging. Annu. Rev. Cell Dev. Biol. 2006, 22, 531–557. [Google Scholar] [CrossRef] [PubMed]
- Inoue, N.; Yahagi, N.; Yamamoto, T.; Ishikawa, M.; Watanabe, K.; Matsuzaka, T.; Nakagawa, Y.; Takeuchi, Y.; Kobayashi, K.; Takahashi, A.; et al. Cyclin-dependent kinase inhibitor, p21WAF1/CIP1, is involved in adipocyte differentiation and hypertrophy, linking to obesity, and insulin resistance. J. Biol. Chem. 2008, 283, 21220–21229. [Google Scholar] [CrossRef] [PubMed]
- Tavana, O.; Zhu, C. Too many breaks (brakes): Pancreatic beta-cell senescence leads to diabetes. Cell Cycle 2011, 10, 2471–2484. [Google Scholar] [CrossRef] [PubMed]
- Silva Figueiredo, P.; Carla Inada, A.; Marcelino, G.; Maiara Lopes Cardozo, C.; de Cassia Freitas, K.; de Cassia Avellaneda Guimaraes, R.; Pereira de Castro, A.; Aragao do Nascimento, V.; Aiko Hiane, P. Fatty Acids Consumption: The Role Metabolic Aspects Involved in Obesity and Its Associated Disorders. Nutrients 2017, 9, 1158. [Google Scholar] [CrossRef] [PubMed]
- Ladeira, C.; Carolino, E.; Gomes, M.C.; Brito, M. Role of Macronutrients and Micronutrients in DNA Damage: Results from a Food Frequency Questionnaire. Nutr. Metab. Insights 2017, 10, 1178638816684666. [Google Scholar] [CrossRef] [PubMed]
- EFSA Panel on Dietetic Products, Nutrition and Allergies (NDA). Scientific Opinion on Dietary Reference Values for energy. EFSA J. 2013, 11, 3005. [Google Scholar]
- Drygas, W.; Niklas, A.A.; Piwońska, A.; Piotrowski, W.; Flotyńska, A.; Kwaśniewska, M.; Nadrowski, P.; Puch-Walczak, A.; Szafraniec, K.; Bielecki, W.; et al. Multi-centre National Population Health Examination Survey (WOBASZ II study): Assumptions, methods, and implementation. Kardiol. Pol. 2016, 74, 681–690. [Google Scholar] [CrossRef] [PubMed]
- Santhakumar, A.B.; Bulmer, A.C.; Singh, I. A review of the mechanisms and effectiveness of dietary polyphenols in reducing oxidative stress and thrombotic risk. J. Hum. Nutr. Diet. 2014, 27, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Fang, Y.Z.; Yang, S.; Wu, G. Free radicals, antioxidants, and nutrition. Nutrition 2002, 18, 872–879. [Google Scholar] [CrossRef]
- Ulrich-Merzenich, G.; Zeitler, H.; Vetter, H.; Kraft, K. Synergy research: Vitamins and secondary plant components in the maintenance of the redox-homeostasis and in cell signaling. Phytomedicine 2009, 16, 2–16. [Google Scholar] [CrossRef] [PubMed]
- Mitjavila, M.T.; Fandos, M.; Salas-Salvado, J.; Covas, M.I.; Borrego, S.; Estruch, R.; Lamuela-Raventos, R.; Corella, D.; Martinez-Gonzalez, M.A.; Sanchez, J.M.; et al. The Mediterranean diet improves the systemic lipid and DNA oxidative damage in metabolic syndrome individuals. A randomized, controlled, trial. Clin. Nutr. 2013, 32, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Rinne, T.; Mutschler, E.; Wimmer-Greinecker, G.; Moritz, A.; Olbrich, H.G. Vitamins C and E protect isolated cardiomyocytes against oxidative damage. Int. J. Cardiol. 2000, 75, 275–281. [Google Scholar] [CrossRef]
- Foksinski, M.; Gackowski, D.; Rozalski, R.; Siomek, A.; Guz, J.; Szpila, A.; Dziaman, T.; Olinski, R. Effects of basal level of antioxidants on oxidative DNA damage in humans. Eur. J. Nutr. 2007, 46, 174–180. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Diet, Nutrition and the Prevention of Chronic Diseases; WHO Technical Report Series 916, 77; World Health Organization: Geneva, Switzerland, 2003. [Google Scholar]
- Pelliccia, F.; Marazzi, G.; Greco, C.; Franzoni, F.; Speziale, G.; Gaudio, C. Current evidence and future perspectives on n-3 PUFAs. Int. J. Cardiol. 2013, 170, 37. [Google Scholar] [CrossRef] [PubMed]
- Reddy, B.S. Dietary fat and cancer: Specific action or caloric effect. J. Nutr. 1986, 116, 1132–1135. [Google Scholar] [CrossRef] [PubMed]
- Bishop, K.S.; Erdrich, S.; Karunasinghe, N.; Han, D.Y.; Zhu, S.; Jesuthasan, A.; Ferguson, L.R. An investigation into the association between DNA damage and dietary fatty acid in men with prostate cancer. Nutrients 2015, 7, 405–422. [Google Scholar] [CrossRef] [PubMed]
- Robinson, L.E.; Buchholz, A.C.; Mazurak, V.C. Inflammation, obesity, and fatty acid metabolism: Influence of n-3 polyunsaturated fatty acids on factors contributing to metabolic syndrome. Appl. Physiol. Nutr. Metab. 2007, 32, 1008–1024. [Google Scholar] [CrossRef] [PubMed]
- Mitterberger, M.C.; Mattesich, M.; Zwerschke, W. Bariatric surgery and diet-induced long-term caloric restriction protect subcutaneous adipose-derived stromal/progenitor cells and prolong their life span in formerly obese humans. Exp. Gerontol. 2014, 56, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Himbert, C.; Thompson, H.; Ulrich, C.M. Effects of Intentional Weight Loss on Markers of Oxidative Stress, DNA Repair and Telomere Length—A Systematic Review. Obes. Facts 2017, 10, 648–665. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Variable | Controls (n = 26) | Obese (n = 88) |
|---|---|---|
| Age (years) | 39.96 ± 7.07 | 38.15 ± 5.55 |
| Height (cm) | 164.58 ± 7.35 | 163.96 ± 6.22 |
| Body weight (kg) | 72.25 ± 13.83 | 87.98 ± 9.66 *** |
| BMI (kg/m2) | 26.33 ± 3.32 | 33.05 ± 2.17 *** |
| WHR | 0.837 ± 0.07 | 0.875 ± 0.06 * |
| Systolic blood pressure (mmHg) | 127.71 ± 19.00 | 125.56 ± 17.22 |
| Diastolic blood pressure (mmHg) | 82.29 ± 9.67 | 82.19 ± 8.19 |
| Fat mass (%) | 32.92 ± 5.38 | 37.78 ± 3.21 *** |
| Fat mass (kg) | 25.12 ± 8.24 | 34.59 ± 6.62 *** |
| Free fat mass (kg) | 47.13 ± 9.75 | 52.22 ± 4.96 *** |
| Total Cholesterol (mg/dL) | 190.42 ± 30.97 | 209.83 ± 36.52 * |
| HDL cholesterol (mg/dL) | 60.23 ± 10.34 | 58.86 ± 15.26 |
| LDL cholesterol (mg/dL) | 110.85 ± 29.14 | 128.17 ± 32.92 * |
| Triglycerides (mg/dL) | 93.93 ± 29.14 | 113.23 ± 48.40 |
| Total Cholesterol/HDL cholesterol | 3.14 ± 0.66 | 3.88 ± 1.30 * |
| Triglycerides/HDL cholesterol | 2.07 ± 1.35 | 2.13 ± 1.20 |
| Glucose (mg/dL) | 84.20 ± 8.58 | 88.12 ± 8.49 * |
| Insulin (mU/mL) | 8.91 ± 2.59 | 11.63 ± 5.20 |
| HOMA | 1.92 ± 0.60 | 2.53 ± 1.19 |
| Apolipoprotein AI (mg/dL) | 163.30 ± 18.34 | 159.55 ± 30.09 |
| CRP-hs (mg/L) | 1.57 ± 0.96 | 3.94 ± 3.51 *** |
| DNA damage (%) | 2.37 ± 0.64 | 5.13 ± 2.46 *** |
| Variable | Controls (n = 26) | Obese (n = 88) | ||
|---|---|---|---|---|
| Mean ± SD | Median (IQR) | Mean ± SD | Median (IQR) | |
| Total Energy (kcal/day) | 1555 ± 362 | 1531 (1371;1676) | 1922 ± 487 ** | 1929 (1599;2226) |
| Protein (% Energy) | 17 ± 4 | 17 (14;18) | 16 ± 4 | 16 (14;19) |
| Carbohydrate (% Energy) | 54 ± 7 | 53 (50;57) | 47 ± 9 ** | 48 (45;52) |
| Total Fat (% Energy) | 32 ± 5 | 33 (31;35) | 36 ± 7 * | 37 (34;40) |
| Saturated Fat (% Energy) | 11 ± 5 | 11 (9;12) | 13 ± 3 ** | 13 (12;15) |
| Monounsaturated Fat (% Energy) | 15 ± 4 | 16 (13;17) | 14 ± 4 | 14 (12;17) |
| Polyunsaturated Fat (% Energy) | 6 ± 2 | 6 (5;7) | 5 ± 1 * | 5 (4;6) |
| Retinol (μg/day) | 398 ± 294 | 330 (162;509) | 393 ± 260 | 336 (216;468) |
| β-carotene (μg/day) | 3715 ± 2090 | 3566 (1891;5863) | 3428 ± 1719 | 3146 (2041;4654) |
| Vitamin E (mg/day) | 12 ± 6 | 12 (8;18) | 10 ± 5 * | 9 (6;13) |
| Vitamin C (mg/day) | 120 ± 52 | 129 (87;157) | 98 ± 57 | 82 (57;139) |
| Variable | All Subjects (n = 114) | Controls (n = 26) | Obese (n = 88) |
|---|---|---|---|
| R | R | R | |
| Age (years) | 0.018 | 0.420 * | 0.091 |
| Body weight (kg) | 0.511 * | 0.635 * | 0.251 * |
| BMI (kg/m2) | 0.596 * | 0.551 * | 0.192 |
| WHR | 0.169 | 0.500 * | −0.032 |
| Fat mass (%) | 0.162 * | 0.598 * | 0.251 * |
| Fat mass (kg) | 0.538 * | 0.670 * | 0.256 |
| Total Cholesterol (mg/dL) | 0.319 * | 0.297 | 0.183 |
| HDL cholesterol (mg/dL) | −0.035 | −0.003 | 0.026 |
| LDL cholesterol (mg/dL) | 0.302 * | 0.248 | 0.155 |
| Triglycerides (mg/dL) | 0.275 * | 0.493 * | 0.202 |
| Glucose (mg/dL) | 0.371 * | −0.057 | 0.358 * |
| Insulin (mU/mL) | 0.128 | −0.515 | 0.045 |
| HOMA | 0.154 | −0.491 | 0.090 |
| Apolipoprotein AI (mg/dL) | −0.068 | −0.173 | 0.035 |
| CRP-hs (mg/L) | 0.457 * | 0.242 | 0.301 * |
| Total Energy (kcal/day) | 0.446 * | 0.188 | 0.316 * |
| Carbohydrate (% Energy) | −0.351 * | −0.342 | −0.099 |
| Total Fat (% Energy) | 0.411 * | 0.231 | 0.269 * |
| Saturated Fat (% Energy) | 0.480 * | 0.249 | 0.335 * |
| Monounsaturated Fat (% Energy) | −0.001 | 0.104 | 0.081 |
| Polyunsaturated Fat (% Energy) | −0.142 | −0.081 | 0.039 |
| Retinol (μg/day) | −0.182 | −0.492 * | −0.257 * |
| β-carotene (μg/day) | −0.148 | −0.325 | −0.193 |
| Vitamin E (mg/day) | −0.316 * | −0.228 | −0.264 * |
| Vitamin C (mg/day) | −0.305 * | −0.069 | −0.324 * |
| Model | B (SE) | P | β | R | R2 | F | P |
|---|---|---|---|---|---|---|---|
| Model 1 | 0.489 | 0.239 | 17.422 | 0.000 | |||
| Age (years) | 0.025 (0.035) | 0.475 | 0.060 | ||||
| BMI (kg/m2) | 0.323 (0.055) | 0.000 | 0.491 | ||||
| Model 2 | 0.542 | 0.294 | 15.262 | 0.000 | |||
| Age (years) | 0.024 (0.033) | 0.465 | 0.059 | ||||
| BMI (kg/m2) | 0.239 (0.060) | 0.000 | 0.364 | ||||
| Total Energy (kcal/day) | 0.001 (0.000) | 0.004 | 0.267 | ||||
| Model 3 | 0.551 | 0.303 | 11.857 | 0.000 | |||
| Age (years) | 0.023 (0.033) | 0.486 | 0.056 | ||||
| BMI (kg/m2) | 0.223 (0.062) | 0.000 | 0.339 | ||||
| Total Energy (kcal/day) | 0.001 (0.000) | 0.022 | 0.226 | ||||
| Saturated Fat (% Energy) | 0.079 (0.066) | 0.231 | 0.112 | ||||
| Model 4 | 0.571 | 0.326 | 13.199 | 0.000 | |||
| Age (years) | 0.031 (0.033) | 0.354 | 0.074 | ||||
| BMI (kg/m2) | 0.212 (0.060) | 0.001 | 0.323 | ||||
| Total Energy (kcal/day) | 0.001 (0.000) | 0.007 | 0.249 | ||||
| Vitamin E (mg/day) | −0.092 (0.040) | 0.024 | −0.188 | ||||
| Model 5 | 0.587 | 0.345 | 14.354 | 0.000 | |||
| Age (years) | 0.040 (0.033) | 0.224 | 0.097 | ||||
| BMI (kg/m2) | 0.213 (0.059) | 0.000 | 0.323 | ||||
| Total Energy (kcal/day) | 0.001 (0.000) | 0.009 | 0.235 | ||||
| Vitamin C (mg/day) | −0.010 (0004) | 0.004 | −0.238 | ||||
| Model 6 | 0.596 | 0.355 | 11.913 | 0.000 | |||
| Age (years) | 0.041 (0.033) | 0.212 | 0.099 | ||||
| BMI (kg/m2) | 0.201 (0.059) | 0.001 | 0.306 | ||||
| Total Energy (kcal/day) | 0.001 (0.000) | 0.011 | 0.230 | ||||
| Vitamin E (mg/day) | −0.056 (0.043) | 0.189 | −0.115 | ||||
| Vitamin C (mg/day) | −0.008 (0.004) | 0.029 | −0.194 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Włodarczyk, M.; Jabłonowska-Lietz, B.; Olejarz, W.; Nowicka, G. Anthropometric and Dietary Factors as Predictors of DNA Damage in Obese Women. Nutrients 2018, 10, 578. https://doi.org/10.3390/nu10050578
Włodarczyk M, Jabłonowska-Lietz B, Olejarz W, Nowicka G. Anthropometric and Dietary Factors as Predictors of DNA Damage in Obese Women. Nutrients. 2018; 10(5):578. https://doi.org/10.3390/nu10050578
Chicago/Turabian StyleWłodarczyk, Marta, Beata Jabłonowska-Lietz, Wioletta Olejarz, and Grażyna Nowicka. 2018. "Anthropometric and Dietary Factors as Predictors of DNA Damage in Obese Women" Nutrients 10, no. 5: 578. https://doi.org/10.3390/nu10050578
APA StyleWłodarczyk, M., Jabłonowska-Lietz, B., Olejarz, W., & Nowicka, G. (2018). Anthropometric and Dietary Factors as Predictors of DNA Damage in Obese Women. Nutrients, 10(5), 578. https://doi.org/10.3390/nu10050578