Macular Xanthophylls Are Related to Intellectual Ability among Adults with Overweight and Obesity

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Participants
2.2. Measures
2.2.1. Anthropometrics and Adiposity Assessment
2.2.2. Dietary Lutein and Zeaxanthin Assessment
2.2.3. Macular Pigment Optical Density
2.2.4. Intellectual Ability Assessment
2.3. Statistical Analyses
3. Results
3.1. Participant Characteristics and Descriptive Information
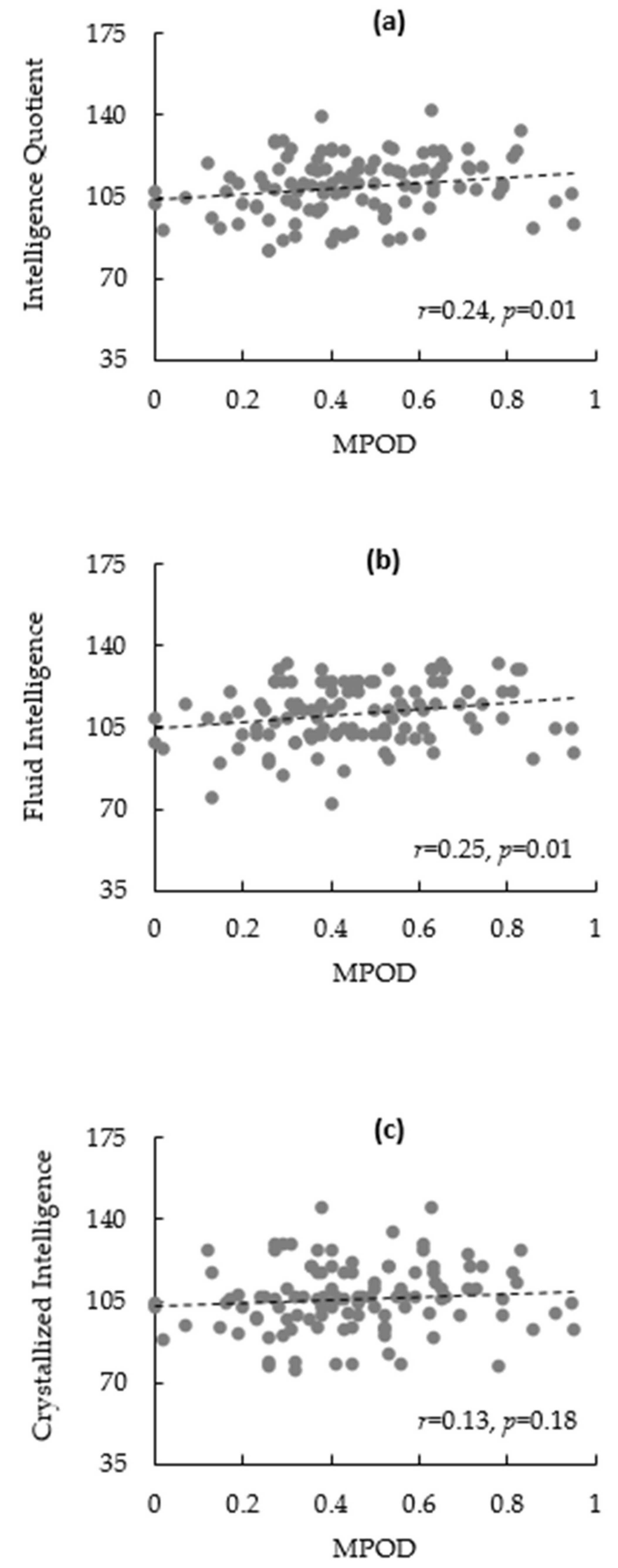
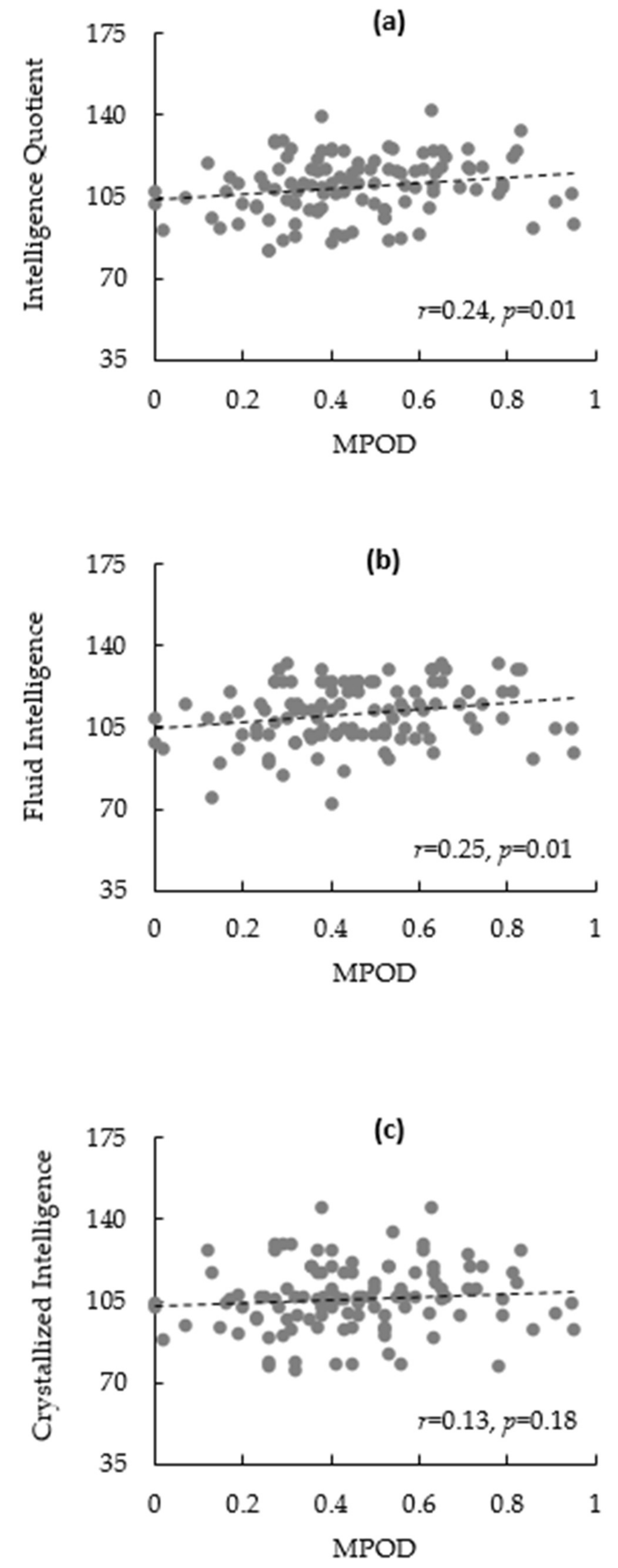
3.2. Bivariate Correlations
3.3. Regression Models
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Hales, C.M.; Carroll, M.D.; Fryar, C.D.; Ogden, C.L. Prevalence of Obesity among Adults and Youth: United States, 2015–2016 Key findings Data from the National Health and Nutrition Examination Survey; US Department of Health and Human Services, Centers for Disease Control and Prevention, National Center for Health Statistics: Atlanta, GA, USA, 2015.
- Malnick, S.D.H. The medical complications of obesity. J. Assoc. Phys. 2006, 99, 565–579. [Google Scholar] [CrossRef] [PubMed]
- Jérôme, A.; Marleau, J.; Baillot, A. Impact of obesity and mood disorders on physical comorbidities, psychological well-being, health behaviours and use of health services. J. Affect. Disord. 2018, 225, 381–388. [Google Scholar] [CrossRef]
- Gustafson, D. Adiposity indices and dementia. Lancet Neurol. 2006, 5, 713–720. [Google Scholar] [CrossRef]
- Luchsinger, J.A.; Gustafson, D.R. Adiposity and Alzheimer’s disease. Curr. Opin. Clin. Nutr. Metab. Care 2009, 12, 15–21. [Google Scholar] [CrossRef] [PubMed]
- Fung, T.T. Association between dietary patterns and plasma biomarkers of obesity and cardiovascular disease risk. Am. J. Clin. Nutr. 2001, 73, 61–67. [Google Scholar] [CrossRef] [PubMed]
- Georgieff, M.K. Nutrition and the developing brain: Nutrient priorities and measurement. Am. J. Clin. Nutr. 2007, 85, 614S–620S. [Google Scholar] [PubMed]
- Sherry, C.L.; Kim, S.S.; Dilger, R.N.; Bauer, L.L.; Moon, M.L.; Tapping, R.I.; Fahey, G.C., Jr.; Tappenden, K.A.; Freund, G.G. Sickness behavior induced by endotoxin can be mitigated by the dietary soluble fiber, pectin, through up-regulation of IL-4 and Th2 polarization. Brain. Behav. Immun. 2010, 24, 631–640. [Google Scholar] [CrossRef] [PubMed]
- Jang, S.; Dilger, R.N.; Johnson, R.W. Luteolin inhibits microglia and alters hippocampal-dependent spatial working memory in aged mice. J. Nutr. 2010, 140, 1892–1898. [Google Scholar] [CrossRef] [PubMed]
- Hoyland, A.; Lawton, C.L.; Dye, L. Acute effects of macronutrient manipulations on cognitive test performance in healthy young adults: A systematic research review. Neurosci. Biobehav. Rev. 2008, 32, 72–85. [Google Scholar] [CrossRef] [PubMed]
- Vishwanathan, R.; Kuchan, M.J.; Sen, S.; Johnson, E.J. Lutein and preterm infants with decreased concentrations of brain carotenoids. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 659–665. [Google Scholar] [CrossRef] [PubMed]
- Friedman, D.; O’Colmain, B.; Munoz, B. Prevalence of age-related macular degeneration in the United States. Arch. Ophthalmol. 2004, 122, 564–572. [Google Scholar] [PubMed]
- Snodderly, D.; Mares, J.; Wooten, B. Macular pigment measurement by heterochromatic flicker photometry in older subjects: The carotenoids and age-related eye disease study. Investig. Ophthalmol. Vis. Sci. 2004, 45, 531–538. [Google Scholar] [CrossRef]
- Johnson, E.; Maras, J.; Rasmussen, H. Intake of lutein and zeaxanthin differ with age, sex, and ethnicity. J. Am. Diet. Assoc. 2010, 110, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Perry, A.; Rasmussen, H. Xanthophyll (lutein, zeaxanthin) content in fruits, vegetables and corn and egg products. Elsevier 2009, 22, 9–15. [Google Scholar] [CrossRef]
- Hammond, B.; Ciulla, T. Macular pigment density is reduced in obese subjects. Investig. Ophthalmol. Vis. Sci. 2002, 43, 47–50. [Google Scholar]
- Nolan, J.; O’Donovan, O.; Kavanagh, H.; Stack, J.; Harrison, M.; Muldoon, A.; Mellerio, J.; Beatty, S. Macular Pigment and Percentage of Body Fat. Investig. Opthalmology Vis. Sci. 2004, 45, 3940–3950. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E. Role of lutein and zeaxanthin in visual and cognitive function throughout the lifespan. Nutr. Rev. 2014, 72, 605–612. [Google Scholar] [CrossRef] [PubMed]
- Renzi-Hammond, L.; Bovier, E.; Fletcher, L.; Miller, L.; Mewborn, C.; Lindbergh, C.; Baxter, J.; Hammond, B. Effects of a Lutein and Zeaxanthin Intervention on Cognitive Function: A Randomized, Double-Masked, Placebo-Controlled Trial of Younger Healthy Adults. Nutrients 2017, 9, 1246. [Google Scholar] [CrossRef] [PubMed]
- Walk, A.; Khan, N.; McCorkle, S.; Drollette, E.; Raine, L.; Kramer, A.; Cohen, N.; Renzi, L.; Hammond, B.; Hillman, C. The Role of Macular Pigment Optical Density in Children’s Attentional Control and Academic Achievement. Psychophysiology 2016, 53, S89. [Google Scholar]
- Colom, R.; Karama, S.; Jung, R.E.; Haier, R.J. Human intelligence and brain networks. Dialogues Clin. Neurosci. 2010, 12, 489–501. [Google Scholar] [PubMed]
- Horn, J.L.; Cattell, R.B. Age differences in fluid and crystallized intelligence. Acta Psychol. 1967, 26, 107–129. [Google Scholar] [CrossRef]
- Horn, J.L.; Cattell, R.B. Refinement and test of the theory of fluid and crystallized general intelligences. J. Educ. Psychol. 1966, 57, 253–270. [Google Scholar] [CrossRef] [PubMed]
- Craik, F.M.; Bialystok, E. Cognition through the lifespan: Mechanisms of change. Trends Cogn. Sci. 2006, 10, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Park, D.C.; Reuter-Lorenz, P. The Adaptive Brain: Aging and Neurocognitive Scaffolding. Annu. Rev. Psychol. 2009, 60, 173–196. [Google Scholar] [CrossRef] [PubMed]
- Zamroziewicz, M.K.; Paul, E.J.; Zwilling, C.E.; Johnson, E.J.; Kuchan, M.J.; Cohen, N.J.; Barbey, A.K. Parahippocampal Cortex Mediates the Relationship between Lutein and Crystallized Intelligence in Healthy, Older Adults. Front. Aging Neurosci. 2016, 8, 297. [Google Scholar] [CrossRef] [PubMed]
- Baumgartner, N.W.; Walk, A.M.; Edwards, C.G.; Covello, A.R.; Chojnacki, M.R.; Reeser, G.E.; Taylor, A.M.; Holscher, H.D.; Khan, N.A. Relationship Between Physical Activity, Adiposity, and Attentional Inhibition. J. Phys. Act. Health 2017, 20, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Stringham, J.M.; Hammond, B.R.; Nolan, J.M.; Wooten, B.R.; Mammen, A.; Smollon, W.; Snodderly, D.M. The utility of using customized heterochromatic flicker photometry (cHFP) to measure macular pigment in patients with age-related macular degeneration. Exp. Eye Res. 2008, 87, 445–453. [Google Scholar] [CrossRef] [PubMed]
- Naugle, R.I.; Chelune, G.J.; Tucker, G.D. Validity of the Kaufman Brief Intelligence Test. Psychol. Assess. 1993, 5, 182–186. [Google Scholar] [CrossRef]
- Wang, J.-J.; Kaufman, A.S. Changes in Fluid and Crystallized Intelligence across the 20- to 90-Year Age Range on the K-Bit. J. Psychoeduc. Assess. 1993, 11, 29–37. [Google Scholar] [CrossRef]
- Swinburn, B.A.; Sacks, G.; Hall, K.D.; McPherson, K.; Finegood, D.T.; Moodie, M.L.; Gortmaker, S.L. The global obesity pandemic: Shaped by global drivers and local environments. Lancet 2011, 378, 804–814. [Google Scholar] [CrossRef]
- Smith, E.; Hay, P.; Campbell, L.; Trollor, J.N. A review of the association between obesity and cognitive function across the lifespan: Implications for novel approaches to prevention and treatment. Obes. Rev. 2011, 12, 740–755. [Google Scholar] [CrossRef] [PubMed]
- Whitmer, R.A.; Gunderson, E.P.; Barrett-Connor, E.; Quesenberry, C.P.; Yaffe, K. Obesity in middle age and future risk of dementia: A 27 year longitudinal population based study. Br. Med. J. 2005, 330, 1360–1364. [Google Scholar] [CrossRef] [PubMed]
- Whitmer, R.A. The epidemiology of adiposity and dementia. Curr. Alzheimer Res. 2007, 4, 117–122. [Google Scholar] [CrossRef] [PubMed]
- Erion, J.R.; Wosiski-Kuhn, M.; Dey, A.; Hao, S.; Davis, C.L.; Pollock, N.K.; Stranahan, A.M. Obesity Elicits Interleukin 1-Mediated Deficits in Hippocampal Synaptic Plasticity. J. Neurosci. 2014, 34, 2618–2631. [Google Scholar] [CrossRef] [PubMed]
- Sanz, C.M.; Ruidavets, J.-B.; Bongard, V.; Marquié, J.-C.; Hanaire, H.; Ferrières, J.; Andrieu, S. Relationship between markers of insulin resistance, markers of adiposity, HbA1c, and cognitive functions in a middle-aged population-based sample: The MONA LISA study. Diabetes Care 2013, 36, 1512–1521. [Google Scholar] [CrossRef] [PubMed]
- Walther, K.; Birdsill, A.C.; Glisky, E.L.; Ryan, L. Structural brain differences and cognitive functioning related to body mass index in older females. Hum. Brain Mapp. 2009, 31, 1052–1064. [Google Scholar] [CrossRef] [PubMed]
- Pannacciulli, N.; Del Parigi, A.; Chen, K.; Le, D.S.N.T.; Reiman, E.M.; Tataranni, P.A. Brain abnormalities in human obesity: A voxel-based morphometric study. Neuroimage 2006, 31, 1419–1425. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.B.; Han, S.P.; Cao, X.G.; Guo, X.R. Intelligence in relation to obesity: A systematic review and meta-analysis. Obes. Rev. 2010, 11, 656–670. [Google Scholar] [CrossRef] [PubMed]
- Sørensen, T.I.A.; Sonne-Holm, S.; Christensen, U.; Krėiner-Møller, S. Reduced Intellectual Performance in Extreme Overweight. Hum. Biol. 1982, 54, 765–775. [Google Scholar] [PubMed]
- Spyridaki, E.C.; Simos, P.; Avgoustinaki, P.D.; Dermitzaki, E.; Venihaki, M.; Bardos, A.N.; Margioris, A.N. The association between obesity and fluid intelligence impairment is mediated by chronic low-grade inflammation. Br. J. Nutr. 2014, 112, 1724–1734. [Google Scholar] [CrossRef] [PubMed]
- Zamroziewicz, M.K.; Talukdar, M.T.; Zwilling, C.E.; Barbey, A.K. Nutritional status, brain network organization, and general intelligence. Neuroimage 2017, 161, 241–250. [Google Scholar] [CrossRef] [PubMed]
- Johnston, C.S.; Taylor, C.A.; Hampl, J.S. More Americans are eating “5 a day” but intakes of dark green and cruciferous vegetables remain low. J. Nutr. 2000, 130, 3063–3067. [Google Scholar] [CrossRef] [PubMed]
- Hammond, B.H., Jr.; Wooten, B. Assessment of the validity of in vivo methods of measuring human macular pigment optical density. Optom. Vis. Sci. 2005, 82, 387–404. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, P.S.; Khachik, F.; Carvalho, L.S.; Muir, G.J.; Zhao, D.-Y.; Katz, N.B. Identification and Quantitation of Carotenoids and their Metabolites in the Tissues of the Human Eye. Exp. Eye Res. 2001, 72, 215–223. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, L.A.; Lau, J.M.; Stein, E.A. Carotenoid composition, concentrations, and relationships in various human organs. Clin. Physiol. Biochem. 1990, 8, 1–10. [Google Scholar] [PubMed]
- Olson, J.A. Serum Levels of Vitamin A and Carotenoids as Reflectors of Nutritional Status. JNCI J. Natl. Cancer Inst. 1984, 73, 1439–1444. [Google Scholar] [CrossRef] [PubMed]
- Gottfredson, L.S. Why g matters: The complexity of everyday life. Intelligence 1997, 24, 79–132. [Google Scholar] [CrossRef]
- Strenze, T. Intelligence and socioeconomic success: A meta-analytic review of longitudinal research. Intelligence 2007, 35, 401–426. [Google Scholar] [CrossRef]
- Power, R.; Coen, R.F.; Beatty, S.; Mulcahy, R.; Moran, R.; Stack, J.; Howard, A.N.; Nolan, J.M. Supplemental Retinal Carotenoids Enhance Memory in Healthy Individuals with Low Levels of Macular Pigment in A Randomized, Double-Blind, Placebo-Controlled Clinical Trial. J. Alzheimer Dis. 2018, 61, 947–961. [Google Scholar] [CrossRef] [PubMed]
- Bergman, I.; Almkvist, O. The effect of age on fluid intelligence is fully mediated by physical health. Arch. Gerontol. Geriatr. 2013, 57, 100–109. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.A.; Shukitt-Hale, B.; Casadesus, G.; Fisher, D. Oxidative Stress and Inflammation in Brain Aging: Nutritional Considerations. Neurochem. Res. 2005, 30, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Witte, A.V.; Kerti, L.; Hermannstädter, H.M.; Fiebach, J.B.; Schreiber, S.J.; Schuchardt, J.P.; Hahn, A.; Flöel, A. Long-Chain Omega-3 Fatty Acids Improve Brain Function and Structure in Older Adults. Cereb. Cortex 2014, 24, 3059–3068. [Google Scholar] [CrossRef] [PubMed]
- Marshalek, B.; Lohman, D.F.; Snow, R.E. The complexity continuum in the radex and hierarchical models of intelligence. Intelligence 1983, 7, 107–127. [Google Scholar] [CrossRef]
- Widomska, J.; Subczynski, W.K. Why has Nature Chosen Lutein and Zeaxanthin to Protect the Retina? J. Clin. Exp. Ophthalmol. 2014, 5, 326. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.; Pan, Y.; Kao, S.-Y.; Li, C.; Kohane, I.; Chan, J.; Yankner, B.A. Gene regulation and DNA damage in the ageing human brain. Nature 2004, 429, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Erdman, J.; Smith, J.; Kuchan, M.; Mohn, E.; Johnson, E.; Rubakhin, S.; Wang, L.; Sweedler, J.; Neuringer, M. Lutein and Brain Function. Foods 2015, 4, 547–564. [Google Scholar] [CrossRef] [PubMed]
- Renzi, L.M.; Dengler, M.J.; Puente, A.; Miller, L.S.; Hammond, B.R. Relationships between macular pigment optical density and cognitive function in unimpaired and mildly cognitively impaired older adults. Neurobiol. Aging 2014, 35, 1695–1699. [Google Scholar] [CrossRef] [PubMed]
- Colom, R.; Haier, R.J.; Head, K.; Álvarez-Linera, J.; Quiroga, M.Á.; Shih, P.C.; Jung, R.E. Gray matter correlates of fluid, crystallized, and spatial intelligence: Testing the P-FIT model. Intelligence 2009, 37, 124–135. [Google Scholar] [CrossRef]
- Lieblein-Boff, J.C.; Johnson, E.J.; Kennedy, A.D.; Lai, C.-S.; Kuchan, M.J. Exploratory Metabolomic Analyses Reveal Compounds Correlated with Lutein Concentration in Frontal Cortex, Hippocampus, and Occipital Cortex of Human Infant Brain. PLoS ONE 2015, 10, e0136904. [Google Scholar] [CrossRef] [PubMed]
- Mewborn, C.M.; Terry, D.P.; Renzi-Hammond, L.M.; Hammond, B.R.; Miller, L.S. Relation of Retinal and Serum Lutein and Zeaxanthin to White Matter Integrity in Older Adults: A Diffusion Tensor Imaging Study. Arch. Clin. Neuropsychol. 2017, 17, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lindbergh, C.A.; Renzi-Hammond, L.M.; Hammond, B.R.; Terry, D.P.; Mewborn, C.M.; Puente, A.N.; Miller, L.S. Lutein and Zeaxanthin Influence Brain Function in Older Adults: A Randomized Controlled Trial. J. Int. Neuropsychol. Soc. 2018, 24, 77–90. [Google Scholar] [CrossRef] [PubMed]
- Curran-Celentano, J.; Hammond, B.R.; Ciulla, T.A.; Cooper, D.A.; Pratt, L.M.; Danis, R.B. Relation between dietary intake, serum concentrations, and retinal concentrations of lutein and zeaxanthin in adults in a Midwest population. Am. J. Clin. Nutr. 2001, 74, 796–802. [Google Scholar] [CrossRef] [PubMed]
- Dhurandhar, N.V.; Schoeller, D.; Brown, A.W.; Heymsfield, S.B.; Thomas, D.; Sørensen, T.I.A.; Speakman, J.R.; Jeansonne, M.; Allison, D.B. Energy balance measurement: When something is not better than nothing. Int. J. Obes. 2015, 39, 1109–1113. [Google Scholar] [CrossRef] [PubMed]
- Subar, A.F.; Freedman, L.S.; Tooze, J.A.; Kirkpatrick, S.I.; Boushey, C.; Neuhouser, M.L.; Thompson, F.E.; Potischman, N.; Guenther, P.M.; Tarasuk, V.; et al. Addressing Current Criticism Regarding the Value of Self-Report Dietary Data. J. Nutr. 2015, 145, 2639–2645. [Google Scholar] [CrossRef] [PubMed]
- Schoeller, D.A.; Thomas, D.; Archer, E.; Heymsfield, S.B.; Blair, S.N.; Goran, M.I.; Hill, J.O.; Atkinson, R.L.; Corkey, B.E.; Foreyt, J.; et al. Self-report–based estimates of energy intake offer an inadequate basis for scientific conclusions. Am. J. Clin. Nutr. 2013, 97, 1413–1415. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Variable | Mean ± SD |
|---|---|
| Age, years | 34.3 ± 6.1 |
| Sex | 69 (F), 45 (M) |
| Income | |
| Low [<$41,000], n (%) | 37 (32) |
| Medium [$41,000–$70,000], n (%) | 36 (32) |
| High [>$70,000], n (%) | 41 (36) |
| Dietary Lutein and Zeaxanthin, µg | 2319.7 ± 3503.0 |
| Body Mass Index, kg/m2 | 31.9 ± 5.3 |
| Overweight, n (%) | 55 (48) |
| Obese, n (%) | 59 (52) |
| Whole body Adiposity, % | 37.7 ± 8.9 |
| Macular Pigment Optical Density | 0.46 ± 0.21 |
| Intelligence Quotient | 109.5 ± 12.9 |
| Crystallized Intelligence | 110.8 ± 12.9 |
| Fluid Intelligence | 106.0 ± 15.0 |
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | ||
|---|---|---|---|---|---|---|---|---|---|
| 1. Age | r | ||||||||
| p | |||||||||
| 2. a Sex | r | −0.04 | |||||||
| p | 0.68 | ||||||||
| 3. Income | r | 0.50 ** | −0.05 | ||||||
| p | 0.00 | 0.57 | |||||||
| 4. Dietary LZ | r | −0.08 | −0.01 | 0.09 | |||||
| p | 0.38 | 0.88 | 0.33 | ||||||
| 5. %Fat | r | 0.06 | −0.77 ** | 0.06 | −0.09 | ||||
| p | 0.55 | 0.00 | 0.52 | 0.35 | |||||
| 6. MPOD | r | −0.18 | 0.18 | −0.06 | 0.06 | −0.19 * | |||
| p | 0.06 | 0.06 | 0.55 | 0.52 | 0.04 | ||||
| 7. IQ | r | 0.02 | 0.26 ** | 0.15 | 0.16 | −0.22 * | 0.24 * | ||
| p | 0.81 | 0.01 | 0.12 | 0.08 | 0.02 | 0.01 | |||
| 8. Fluid Intelligence | r | −0.01 | 0.23 * | 0.08 | 0.14 | −0.24 * | 0.25 ** | 0.80 ** | |
| p | 0.95 | 0.01 | 0.38 | 0.14 | 0.01 | 0.01 | 0.00 | ||
| 9. Crystallized Intelligence | r | 0.11 | 0.20 * | 0.18 | 0.13 | −0.13 | 0.13 | 0.85 ** | 0.36 ** |
| p | 0.27 | 0.03 | 0.05 | 0.16 | 0.16 | 0.18 | 0.00 | 0.00 |
| Intelligence Quotient | Fluid Intelligence | Crystallized Intelligence | |||||||
|---|---|---|---|---|---|---|---|---|---|
| β | p | Model R2 | β | p | Model R2 | β | p | Model R2 | |
| 0.15 ** | 0.13 * | 0.11 | |||||||
| Age | <0.01 | 1.00 | <0.01 | 0.99 | 0.06 | 0.56 | |||
| Sex | 0.24 | 0.09 | 0.13 | 0.37 | 0.27 | 0.07 | |||
| Income | 0.16 | 0.14 | 0.10 | 0.37 | 0.15 | 0.16 | |||
| Dietary LZ | 0.14 | 0.12 | 0.11 | 0.23 | 0.13 | 0.17 | |||
| %Fat | 0.01 | 0.93 | −0.09 | 0.52 | 0.09 | 0.53 | |||
| MPOD | 0.20 * | 0.04 | 0.20 * | 0.03 | 0.11 | 0.25 | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khan, N.A.; Walk, A.M.; Edwards, C.G.; Jones, A.R.; Cannavale, C.N.; Thompson, S.V.; Reeser, G.E.; Holscher, H.D. Macular Xanthophylls Are Related to Intellectual Ability among Adults with Overweight and Obesity. Nutrients 2018, 10, 396. https://doi.org/10.3390/nu10040396
Khan NA, Walk AM, Edwards CG, Jones AR, Cannavale CN, Thompson SV, Reeser GE, Holscher HD. Macular Xanthophylls Are Related to Intellectual Ability among Adults with Overweight and Obesity. Nutrients. 2018; 10(4):396. https://doi.org/10.3390/nu10040396
Chicago/Turabian StyleKhan, Naiman A., Anne. M. Walk, Caitlyn G. Edwards, Alicia R. Jones, Corinne N. Cannavale, Sharon V. Thompson, Ginger E. Reeser, and Hannah D. Holscher. 2018. "Macular Xanthophylls Are Related to Intellectual Ability among Adults with Overweight and Obesity" Nutrients 10, no. 4: 396. https://doi.org/10.3390/nu10040396
APA StyleKhan, N. A., Walk, A. M., Edwards, C. G., Jones, A. R., Cannavale, C. N., Thompson, S. V., Reeser, G. E., & Holscher, H. D. (2018). Macular Xanthophylls Are Related to Intellectual Ability among Adults with Overweight and Obesity. Nutrients, 10(4), 396. https://doi.org/10.3390/nu10040396