Effect of a Protein Supplement on the Gut Microbiota of Endurance Athletes: A Randomized, Controlled, Double-Blind Pilot Study



 ,
,  ,
, 
Abstract

1. Introduction
2. Materials and Methods
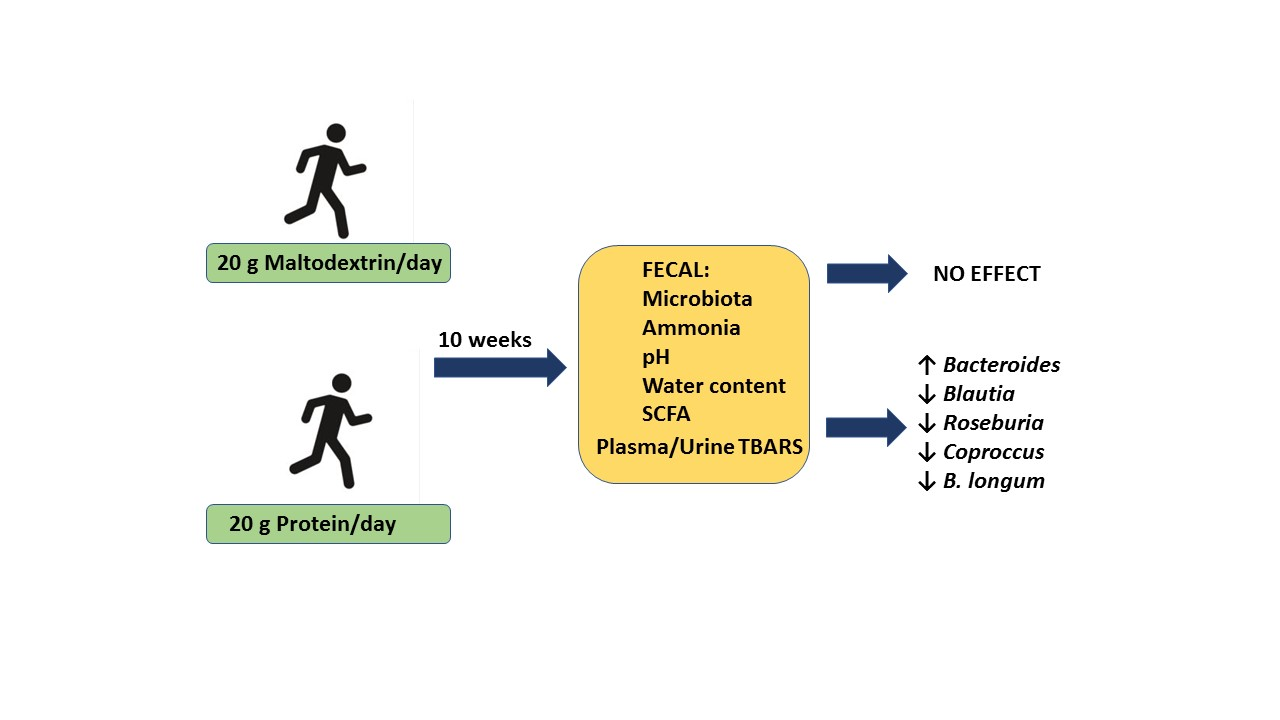
2.1. Study Design
2.2. Subjects and Dietary Supplementation
2.3. Anthropometry and Body Composition
2.4. Food Frequency Questionnaire
2.5. Stool Collection and Bacterial DNA Extraction
2.6. Sequencing and Bioinformatics
2.7. Quantitative PCR Analysis
2.8. Fecal Water, pH, and Ammonia Content
2.9. Short-Chain Fatty Acids
2.10. Urine and Plasma Thiobarbituric Acid-Reacting Substances
2.11. Statistical Analysis
3. Results
3.1. Subjects and Body Composition
3.2. Dietary Intake
3.3. Fecal Water Content, pH, and Ammonia
3.4. Short-Chain Fatty Acids
3.5. Plasma and Urine TBARS
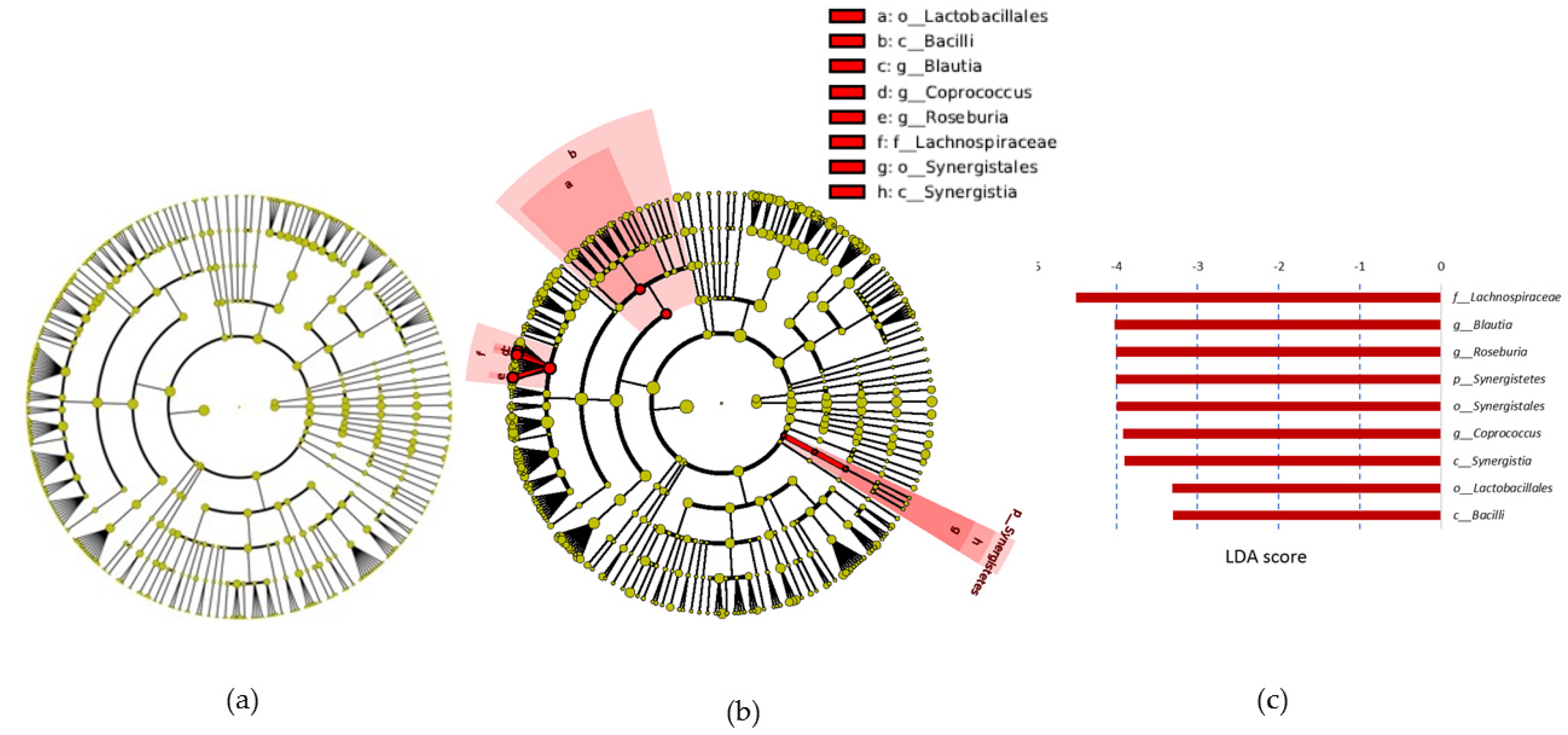
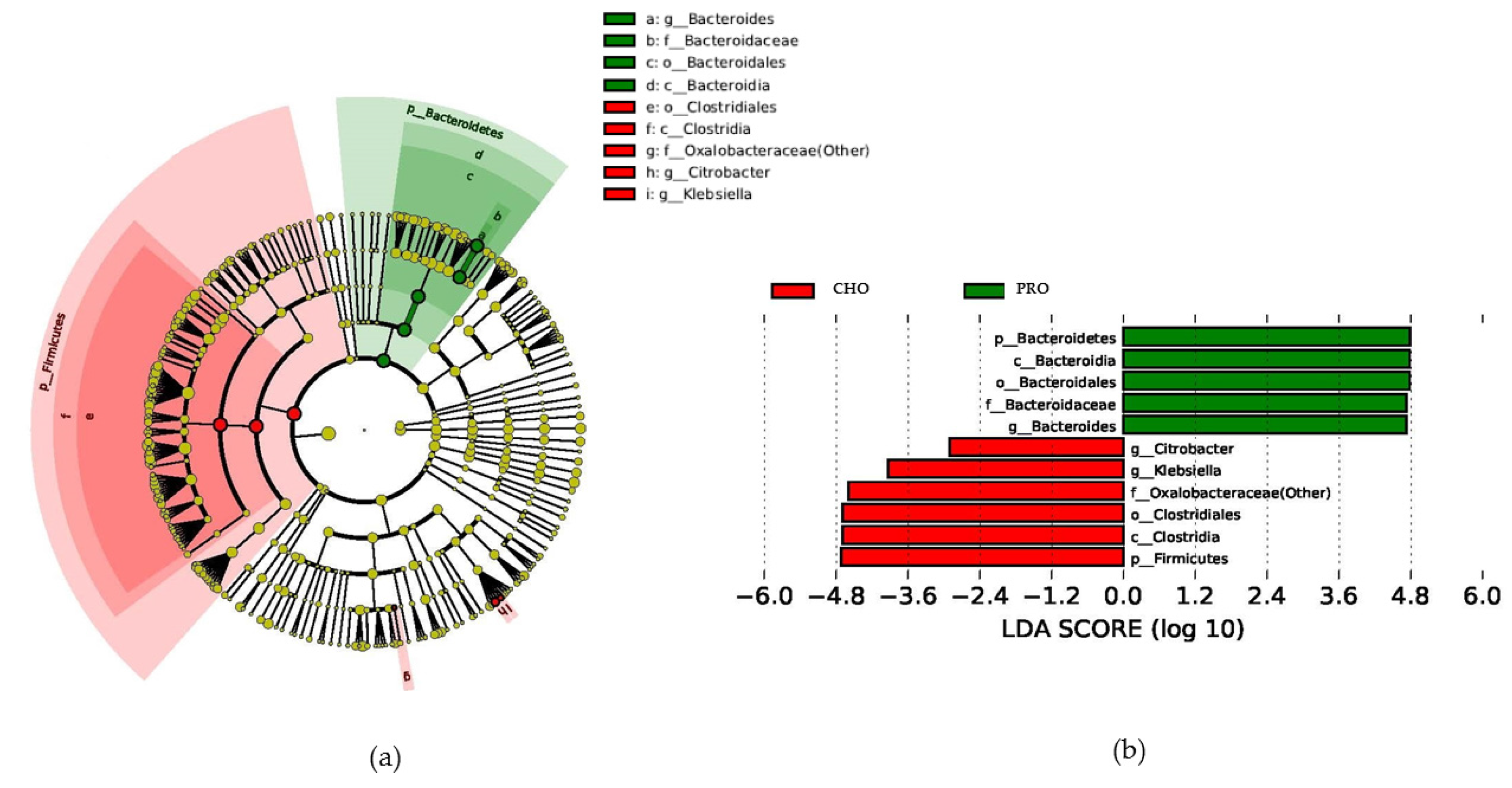
3.6. Fecal Microbiota
3.7. Quantification of Specific Bacterial Species by qPCR
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. American college of sports medicine joint position statement. Nutrition and athletic performance. Med. Sci. Sports Exerc. 2016, 48, 543–568. [Google Scholar] [PubMed]
- Phillips, S.M.; Van Loon, L.J. Dietary protein for athletes: From requirements to optimum adaptation. J. Sports Sci. 2011, 29, S29–S38. [Google Scholar] [CrossRef] [PubMed]
- Phillips, S.M.; Chevalier, S.; Leidy, H.J. Protein “requirements” beyond the rda: Implications for optimizing health. Appl. Physiol. Nutr. Metab. 2016, 41, 565–572. [Google Scholar] [CrossRef] [PubMed]
- Pasiakos, S.M.; McLellan, T.M.; Lieberman, H.R. The effects of protein supplements on muscle mass, strength, and aerobic and anaerobic power in healthy adults: A systematic review. Sports Med. 2015, 45, 111–131. [Google Scholar] [CrossRef] [PubMed]
- Morton, R.W.; Murphy, K.T.; McKellar, S.R.; Schoenfeld, B.J.; Henselmans, M.; Helms, E.; Aragon, A.A.; Devries, M.C.; Banfield, L.; Krieger, J.W.; et al. A systematic review, meta-analysis and meta-regression of the effect of protein supplementation on resistance training-induced gains in muscle mass and strength in healthy adults. Br. J. Sports Med. 2018, 52, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Tekin, K.A.; Kravitz, L. The growing trend of ergogenic drugs and supplements. ACSM’S Health Fit. J. 2004, 8, 15–18. [Google Scholar] [CrossRef]
- Lopez-Legarrea, P.; Fuller, N.R.; Zulet, M.A.; Martinez, J.A.; Caterson, I.D. The influence of mediterranean, carbohydrate and high protein diets on gut microbiota composition in the treatment of obesity and associated inflammatory state. Asia Pac. J. Clin. Nutr. 2014, 23, 360–368. [Google Scholar] [PubMed]
- Portune, K.J.; Beaumont, M.; Davila, A.-M.; Tomé, D.; Blachier, F.; Sanz, Y. Gut microbiota role in dietary protein metabolism and health-related outcomes: The two sides of the coin. Trends Food Sci. Technol. 2016, 57, 213–232. [Google Scholar] [CrossRef]
- Rowland, I.; Gibson, G.; Heinken, A.; Scott, K.; Swann, J.; Thiele, I.; Tuohy, K. Gut microbiota functions: Metabolism of nutrients and other food components. Eur. J. Nutr. 2017, 57, 1–24. [Google Scholar] [CrossRef] [PubMed]
- Clausen, M.R.; Mortensen, P.B.B. Fecal ammonia in patients with adenomatous polyps and cancer of the colon. Nutr. Cancer 1992. [Google Scholar] [CrossRef] [PubMed]
- Hullar, M.A.; Burnett-Hartman, A.N.; Lampe, J.W. Gut microbes, diet, and cancer. In Advances in Nutrition and Cancer; Springer: Berlin, Germany, 2014; pp. 377–399. [Google Scholar]
- Ríos-Covián, D.; Ruas-Madiedo, P.; Margolles, A.; Gueimonde, M.; de los Reyes-Gavilán, C.G.; Salazar, N. Intestinal short chain fatty acids and their link with diet and human health. Front. Microbiol. 2016, 7, 185. [Google Scholar] [CrossRef] [PubMed]
- Neis, E.P.; Dejong, C.H.; Rensen, S.S. The role of microbial amino acid metabolism in host metabolism. Nutrients 2015, 7, 2930–2946. [Google Scholar] [CrossRef] [PubMed]
- Cerdá, B.; Pérez, M.; Pérez-Santiago, J.D.; Tornero-Aguilera, J.F.; González-Soltero, R.; Larrosa, M. Gut microbiota modification: Another piece in the puzzle of the benefits of physical exercise in health? Front. Physiol. 2016, 7, 51. [Google Scholar] [CrossRef] [PubMed]
- Bressa, C.; Bailén-Andrino, M.; Pérez-Santiago, J.; González-Soltero, R.; Pérez, M.; Montalvo-Lominchar, M.G.; Maté-Muñoz, J.L.; Domínguez, R.; Moreno, D.; Larrosa, M. Differences in gut microbiota profile between women with active lifestyle and sedentary women. PLoS ONE 2017, 12, e0171352. [Google Scholar] [CrossRef] [PubMed]
- Clarke, S.F.; Murphy, E.F.; O’sullivan, O.; Lucey, A.J.; Humphreys, M.; Hogan, A.; Hayes, P.; O’reilly, M.; Jeffery, I.B.; Wood-Martin, R.; et al. Exercise and associated dietary extremes impact on gut microbial diversity. Gut 2014, 63, 1913–1920. [Google Scholar] [CrossRef] [PubMed]
- Naclerio, F.; Seijo-Bujia, M.; Larumbe-Zabala, E.; Earnest, C.P. Carbohydrates alone or mixing with beef or whey protein promote similar training outcomes in resistance training males: A double blind, randomized controlled clinical trial. Int. J. Sport Nutr. Exerc. Metab. 2017, 27, 408–420. [Google Scholar] [CrossRef] [PubMed]
- Negro, M.; Vandoni, M.; Ottobrini, S.; Codrons, E.; Correale, L.; Buonocore, D.; Marzatico, F. Protein supplementation with low fat meat after resistance training: Effects on body composition and strength. Nutrients 2014, 6, 3040–3049. [Google Scholar] [CrossRef] [PubMed]
- Haro, C.; Rangel-Zúñiga, O.A.; Alcalá-Díaz, J.F.; Gómez-Delgado, F.; Pérez-Martínez, P.; Delgado-Lista, J.; Quintana-Navarro, G.M.; Landa, B.B.; Navas-Cortés, J.A.; Tena-Sempere, M.; et al. Intestinal microbiota is influenced by gender and body mass index. PLoS ONE 2016, 11, e0154090. [Google Scholar] [CrossRef] [PubMed]
- Bridgewater, L.C.; Zhang, C.; Wu, Y.; Hu, W.; Zhang, Q.; Wang, J.; Li, S.; Zhao, L. Gender-based differences in host behavior and gut microbiota composition in response to high fat diet and stress in a mouse model. Sci. Rep. 2017, 7, 10776. [Google Scholar] [CrossRef] [PubMed]
- Morton, R.W.; McGlory, C.; Phillips, S.M. Nutritional interventions to augment resistance training-induced skeletal muscle hypertrophy. Front. Physiol. 2015, 6, 245. [Google Scholar] [CrossRef] [PubMed]
- Areta, J.L.; Burke, L.M.; Ross, M.L.; Camera, D.M.; West, D.W.; Broad, E.M.; Jeacocke, N.A.; Moore, D.R.; Stellingwerff, T.; Phillips, S.M.; et al. Timing and distribution of protein ingestion during prolonged recovery from resistance exercise alters myofibrillar protein synthesis. J. Physiol. 2013, 591, 2319–2331. [Google Scholar] [CrossRef] [PubMed]
- Cuthbertson, D.; Smith, K.; Babraj, J.; Leese, G.; Waddell, T.; Atherton, P.; Wackerhage, H.; Taylor, P.M.; Rennie, M.J. Anabolic signaling deficits underlie amino acid resistance of wasting, aging muscle. FASEB J. 2005, 19, 422–424. [Google Scholar] [CrossRef] [PubMed]
- Moore, D.R.; Robinson, M.J.; Fry, J.L.; Tang, J.E.; Glover, E.I.; Wilkinson, S.B.; Prior, T.; Tarnopolsky, M.A.; Phillips, S.M. Ingested protein dose response of muscle and albumin protein synthesis after resistance exercise in young men. Am. J. Clin. Nutr. 2008, 89, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Witard, O.C.; Jackman, S.R.; Breen, L.; Smith, K.; Selby, A.; Tipton, K.D. Myofibrillar muscle protein synthesis rates subsequent to a meal in response to increasing doses of whey protein at rest and after resistance exercise. Am. J. Clin. Nutr. 2013, 99, 86–95. [Google Scholar] [CrossRef] [PubMed]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16s ribosomal rna gene pcr primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I. Qiime allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [PubMed]
- Lozupone, C.; Knight, R. Unifrac: A new phylogenetic method for comparing microbial communities. Appl. Environ. Microbiol. 2005, 71, 8228–8235. [Google Scholar] [CrossRef] [PubMed]
- Segata, N.; Izard, J.; Waldron, L.; Gevers, D.; Miropolsky, L.; Garrett, W.S.; Huttenhower, C. Metagenomic biomarker discovery and explanation. Genome Biol. 2011, 12, R60. [Google Scholar] [CrossRef] [PubMed]
- Son, J.S.; Zheng, L.J.; Rowehl, L.M.; Tian, X.; Zhang, Y.; Zhu, W.; Litcher-Kelly, L.; Gadow, K.D.; Gathungu, G.; Robertson, C.E.; et al. Comparison of fecal microbiota in children with autism spectrum disorders and neurotypical siblings in the simons simplex collection. PLoS ONE 2015, 10, e0137725. [Google Scholar] [CrossRef] [PubMed]
- McOrist, A.L.; Warhurst, M.; McOrist, S.; Bird, A.R. Colonic infection by bilophila wadsworthia in pigs. J. Clin. Microbiol. 2001, 39, 1577–1579. [Google Scholar] [CrossRef] [PubMed]
- Ott, S.J.; Musfeldt, M.; Ullmann, U.; Hampe, J.; Schreiber, S. Quantification of intestinal bacterial populations by real-time pcr with a universal primer set and minor groove binder probes: A global approach to the enteric flora. J. Clin. Microbiol. 2004, 42, 2566–2572. [Google Scholar] [CrossRef] [PubMed]
- Brankatschk, R.; Bodenhausen, N.; Zeyer, J.; Bürgmann, H. Simple absolute quantification method correcting for quantitative pcr efficiency variations for microbial community samples. Appl. Environ. Microbiol. 2012, 78, 4481–4489. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Karring, H. A determination and comparison of urease activity in feces and fresh manure from pig and cattle in relation to ammonia production and ph changes. PLoS ONE 2014, 9, e110402. [Google Scholar] [CrossRef] [PubMed]
- García-Villalba, R.; Giménez-Bastida, J.A.; García-Conesa, M.T.; Tomás-Barberán, F.A.; Carlos Espín, J.; Larrosa, M. Alternative method for gas chromatography-mass spectrometry analysis of short-chain fatty acids in faecal samples. J. Sep. Sci. 2012, 35, 1906–1913. [Google Scholar] [CrossRef] [PubMed]
- Junge, W.; Wilke, B.; Halabi, A.; Klein, G. Determination of reference intervals for serum creatinine, creatinine excretion and creatinine clearance with an enzymatic and a modified jaffe method. Clin. Chim. Acta 2004, 344, 137–148. [Google Scholar] [CrossRef] [PubMed]
- Rankin, A.; O’Donavon, C.; Madigan, S.M.; O’Sullivan, O.; Cotter, P.D. ‘Microbes in sport’—The potential role of the gut microbiota in athlete health and performance. Br. J. Sports Med. 2017, 51, 698–699. [Google Scholar] [CrossRef] [PubMed]
- Cermak, N.M.; de Groot, L.C.; Saris, W.H.; van Loon, L.J. Protein supplementation augments the adaptive response of skeletal muscle to resistance-type exercise training: A meta-analysis. Am. J. Clin. Nutr. 2012, 96, 1454–1464. [Google Scholar] [CrossRef] [PubMed]
- De Moraes, W.M.A.M.; Mendes, A.E.P.; Lopes, M.M.M.; Maia, F.M.M. Protein overfeeding is associated with improved lipid and anthropometric profile thus lower malondialdehyde levels in resistance-trained athletes. Int. J. Sports Sci. 2017, 7, 87–93. [Google Scholar]
- Ma, N.; Tian, Y.; Wu, Y.; Ma, X. Contributions of the interaction between dietary protein and gut microbiota to intestinal health. Curr. Protein Pept. Sci. 2017, 18, 795–808. [Google Scholar] [CrossRef] [PubMed]
- Beaumont, M.; Portune, K.J.; Steuer, N.; Lan, A.; Cerrudo, V.; Audebert, M.; Dumont, F.; Mancano, G.; Khodorova, N.; Andriamihaja, M. Quantity and source of dietary protein influence metabolite production by gut microbiota and rectal mucosa gene expression: A randomized, parallel, double-blind trial in overweight humans. Am. J. Clin. Nutr. 2017, 106, 1005–1019. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Blouin, J.M.; Santacruz, A.; Lan, A.; Andriamihaja, M.; Wilkanowicz, S.; Benetti, P.H.; Tome, D.; Sanz, Y.; Blachier, F.; et al. High-protein diet modifies colonic microbiota and luminal environment but not colonocyte metabolism in the rat model: The increased luminal bulk connection. Am. J. Physiol. Gastrointest. Liver Physiol. 2014, 307, G459–G470. [Google Scholar] [CrossRef] [PubMed]
- Butteiger, D.N.; Hibberd, A.A.; McGraw, N.J.; Napawan, N.; Hall-Porter, J.M.; Krul, E.S. Soy protein compared with milk protein in a western diet increases gut microbial diversity and reduces serum lipids in golden syrian hamsters. J. Nutr. 2016, 146, 697–705. [Google Scholar] [CrossRef] [PubMed]
- Conlon, M.A.; Bird, A.R. The impact of diet and lifestyle on gut microbiota and human health. Nutrients 2014, 7, 17–44. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.; Cummings, J.; Allison, C. Protein degradation by human intestinal bacteria. Microbiology 1986, 132, 1647–1656. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.; Allison, C.; Gibson, S.; Cummings, J. Contribution of the microflora to proteolysis in the human large intestine. J. Appl. Microbiol. 1988, 64, 37–46. [Google Scholar] [CrossRef]
- Yatsunenko, T.; Rey, F.E.; Manary, M.J.; Trehan, I.; Dominguez-Bello, M.G.; Contreras, M.; Magris, M.; Hidalgo, G.; Baldassano, R.N.; Anokhin, A.P. Human gut microbiome viewed across age and geography. Nature 2012, 486, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Murphy, E.A.; Velazquez, K.T.; Herbert, K.M. Influence of high-fat-diet on gut microbiota: A driving force for chronic disease risk. Curr. Opin. Clin. Nutr. Metab. Care 2015, 18, 515. [Google Scholar] [CrossRef] [PubMed]
- Macfarlane, G.T. The colonic flora, fermentation and large bowel digestive function. In The Large Intestine: Physiology, Pathophysiology and Disease; Raven: New York, NY, USA, 1991; pp. 51–92. [Google Scholar]
- Davila, A.-M.; Blachier, F.; Gotteland, M.; Andriamihaja, M.; Benetti, P.-H.; Sanz, Y.; Tomé, D. Re-print of “intestinal luminal nitrogen metabolism: Role of the gut microbiota and consequences for the host”. Pharmacol. Res. 2013, 69, 114–126. [Google Scholar] [CrossRef] [PubMed]
- Barcenilla, A.; Pryde, S.E.; Martin, J.C.; Duncan, S.H.; Stewart, C.S.; Henderson, C.; Flint, H.J. Phylogenetic relationships of butyrate-producing bacteria from the human gut. Appl. Environ. Microbiol. 2000, 66, 1654–1661. [Google Scholar] [CrossRef] [PubMed]
- Venkataraman, A.; Schmidt, A.; Waldron, C.; Sieber, J.; Theis, K.; Schmidt, T. Variable responses of human microbiomes to dietary supplementation with resistant starch. Microbiome 2016, 4, 33. [Google Scholar] [CrossRef] [PubMed]
- Russell, W.R.; Gratz, S.W.; Duncan, S.H.; Holtrop, G.; Ince, J.; Scobbie, L.; Duncan, G.; Johnstone, A.M.; Lobley, G.E.; Wallace, R.J.; et al. High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. Am. J. Clin. Nutr. 2011, 93, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Den Besten, G.; van Eunen, K.; Groen, A.K.; Venema, K.; Reijngoud, D.-J.; Bakker, B.M. The role of short-chain fatty acids in the interplay between diet, gut microbiota, and host energy metabolism. J. Lipid Res. 2013, 54, 2325–2340. [Google Scholar] [CrossRef] [PubMed]
- Brinkworth, G.D.; Noakes, M.; Clifton, P.M.; Bird, A.R. Comparative effects of very low-carbohydrate, high-fat and high-carbohydrate, low-fat weight-loss diets on bowel habit and faecal short-chain fatty acids and bacterial populations. Br. J. Nutr. 2009, 101, 1493–1502. [Google Scholar] [CrossRef] [PubMed]
- Verbeke, K.A.; Boobis, A.R.; Chiodini, A.; Edwards, C.A.; Franck, A.; Kleerebezem, M.; Nauta, A.; Raes, J.; Van Tol, E.A.; Tuohy, K.M. Towards microbial fermentation metabolites as markers for health benefits of prebiotics. Nutr. Res. Rev. 2015, 28, 42–66. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Liu, F.; Ling, Z.; Tong, X.; Xiang, C. Human intestinal lumen and mucosa-associated microbiota in patients with colorectal cancer. PLoS ONE 2012, 7, e39743. [Google Scholar] [CrossRef] [PubMed]
- Tamanai-Shacoori, Z.; Smida, I.; Bousarghin, L.; Loreal, O.; Meuric, V.; Fong, S.B.; Bonnaure-Mallet, M.; Jolivet-Gougeon, A. Roseburia spp.: A marker of health? Future Microbiol. 2017, 12, 157–170. [Google Scholar] [CrossRef] [PubMed]
- David, L.A.; Maurice, C.F.; Carmody, R.N.; Gootenberg, D.B.; Button, J.E.; Wolfe, B.E.; Ling, A.V.; Devlin, A.S.; Varma, Y.; Fischbach, M.A. Diet rapidly and reproducibly alters the human gut microbiome. Nature 2014, 505, 559–563. [Google Scholar] [CrossRef] [PubMed]
- Mu, C.; Yang, Y.; Luo, Z.; Zhu, W. Temporal microbiota changes of high-protein diet intake in a rat model. Anaerobe 2017, 47, 218–225. [Google Scholar] [CrossRef] [PubMed]
- Plaza-Díaz, J.; Ruiz-Ojeda, F.J.; Vilchez-Padial, L.M.; Gil, A. Evidence of the anti-inflammatory effects of probiotics and synbiotics in intestinal chronic diseases. Nutrients 2017, 9, 555. [Google Scholar] [CrossRef] [PubMed]
- Williams, B.A.; Zhang, D.; Lisle, A.T.; Mikkelsen, D.; McSweeney, C.S.; Kang, S.; Bryden, W.L.; Gidley, M.J. Soluble arabinoxylan enhances large intestinal microbial health biomarkers in pigs fed a red meat-containing diet. Nutrition 2016, 32, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Jäger, R.; Shields, K.A.; Lowery, R.P.; De Souza, E.O.; Partl, J.M.; Hollmer, C.; Purpura, M.; Wilson, J.M. Probiotic bacillus coagulans gbi-30, 6086 reduces exercise-induced muscle damage and increases recovery. PeerJ 2016, 4, e2276. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Nutritional Components | Maltodextrin | Protein | Orange Drink |
|---|---|---|---|
| Energy (kcal) | 91.6 | 89.9 | 113.2 |
| Carbohydrates (g) | 22.9 | 0.4 | 27.9 |
| Protein (g) | 0 | 19.8 | 0.4 |
| Fat (g) | 0 | 1.0 | 0 |
| Fiber (g) | 0 | 0 | 0 |
| SCFA | Target Ion (m/z) | Qualifier Ions (m/z) | Calibration Curve | r2 |
|---|---|---|---|---|
| Acetic acid | 43 | 45; 60 | y = 9.558 × 10−4 + 1.693 × 10−4 | 0.999210 |
| Propionic acid | 74 | 73; 57 | y = 5.977 × 10−4 + 3.041 × 10−2 | 0.999856 |
| Butyric acid | 73 | 60 | y = 7.540 × 10−4 + 2.692 × 10−3 | 0.999797 |
| Isobutyric acid | 73 | 88 | y = 8.346 × 10−4 + 3.447 × 10−2 | 0.999840 |
| Valeric acid | 73 | 60 | y = 1.228 × 10−5 + 5.442 × 10−2 | 0.999841 |
| Isovaleric acid | 87 | 60 | y = 5.519 × 10−4 + 2.783 × 10−2 | 0.999773 |
| Methylvaleric acid | 73 | 74; 83 |
| Parameters | CHO n = 8 | PRO n = 10 | p |
|---|---|---|---|
| Age (years) | 35.38 ± 9.00 | 34.90 ± 9.49 | 0.915 |
| Body weight | 70.07 ± 5.66 | 68.03 ± 4.91 | 0.424 |
| BMI | 23.37 ± 2.44 | 22.11 ± 1.03 | 0.205 |
| BFP (%) | 11.85 ± 4.31 | 10.41 ± 2.72 | 0.402 |
| BFM (kg) | 15.18 ± 5.19 | 13.06 ± 3.37 | 0.311 |
| VAT (g) | 364.20 ± 180.61 | 333.70 ± 134.47 | 0.717 |
| AI (kg/m2) | 5.41 ± 1.93 | 4.59 ± 1.01 | 0.304 |
| MMI (kg/m2) | 17.33 ± 1.22 | 17.07 ± 0.89 | 0.600 |
| FFMI (kg/m2) | 15.56 ± 1.42 | 15.19 ± 1.10 | 0.541 |
| Diet Intake | CHO | PRO | CHO vs. PRO t = 0 | CHO vs. PRO t = 10 Weeks | ||||
|---|---|---|---|---|---|---|---|---|
| t = 0 | t = 10 Weeks | p | t = 0 | t = 10 Weeks | p | p | p | |
| Energy (kcal) | 2735 ± 735 | 2576 ± 702 | 0.300 | 2617 ± 706 | 2693 ± 847 | 0.613 | 0.735 | 0.753 |
| Carbohydrates (g) | 282.58 ± 87.27 | 255.25 ± 68.78 | 0.155 | 286.73 ± 95.85 | 300.63 ± 98.06 | 0.306 | 0.926 | 0.319 |
| Protein (g) | 130.78 ± 31.03 | 128.76 ± 27.30 | 0.700 | 127.05 ± 24.61 | 128.76 ± 27.30 | 0.969 | 0.779 | 0.992 |
| Fat (g) | 116.01 ± 34.51 | 111.41 ± 37.05 | 0.682 | 103.23 ± 29.18 | 105.63 ± 37.74 | 0.821 | 0.407 | 0.749 |
| Fiber (g) | 21.26 ± 5.65 | 20.91 ± 4.57 | 0.576 | 20.91 ± 4.57 | 19.34 ± 3.46 | 0.496 | 0.607 | 0.419 |
| Carbohydrates (%) of energy | 41.37 ± 4.10 | 40.37 ± 4.80 | 0.582 | 43.80 ± 5.13 | 44.80 ± 5.76 | 0.653 | 0.294 | 0.102 |
| Protein (%) of energy | 19.62 ± 2.87 | 20.37 ± 1.99 | 0.365 | 20.00 ± 2.53 | 19.70 ± 2.31 | 0.591 | 0.773 | 0.319 |
| Fat (%) of energy | 38.87 ± 4.91 | 39.00 ± 4.86 | 0.949 | 36.00 ± 4.18 | 35.60 ± 4.94 | 0.844 | 0.199 | 0.164 |
| Carbohydrates (g/kg bw) | 4.03 ± 1.16 | 3.68 ± 0.95 | 0.201 | 4.20 ± 1.30 | 4.45 ± 1.40 | 0.290 | 0.783 | 0.201 |
| Protein (g/kg bw) | 1.86 ± 0.39 | 1.84 ± 0.34 | 0.809 | 1.87 ± 0.38 | 1.90 ± 0.39 | 0.790 | 0.960 | 0.766 |
| Fat (g/kg bw) | 1.66 ± 0.52 | 1.59 ± 0.49 | 0.656 | 1.52 ± 0.44 | 1.58 ± 0.51 | 0.715 | 0.550 | 0.948 |
| Fiber (g/kg bw) | 0.31 ± 0.09 | 0.30 ± 0.08 | 0.757 | 0.29 ± 0.08 | 0.29 ± 0.06 | 0.760 | 0.797 | 0.719 |
| Carbohydrates (kcal/kg bw) | 16.14 ± 4.64 | 14.73 ± 3.82 | 0.201 | 16.80 ± 5.20 | 17.83 ± 5.60 | 0.290 | 0.783 | 0.201 |
| Protein (kcal/kg bw) | 7.46 ± 1.57 | 7.39 ± 1.37 | 0.809 | 7.49 ± 1.50 | 7.60 ± 1.56 | 0.790 | 0.960 | 0.766 |
| Fat (kcal/kg bw) | 14.96 ± 4.76 | 14.34 ± 4.39 | 0.656 | 13.70 ± 3.96 | 14.20 ± 4.63 | 0.715 | 0.550 | 0.948 |
| Diet Intake | CHO | PRO | CHO vs. PRO t = 0 | CHO vs. PRO t = 10 Weeks | ||||
|---|---|---|---|---|---|---|---|---|
| t = 0 | t = 10 Weeks | p | t = 0 | t = 10 Weeks | p | p | p | |
| Energy (kcal) | 2735 ± 735 | 2656 ± 702 | 0.849 | 2617 ± 706 | 2783 ± 847 | 0.583 | 0.735 | 0.525 |
| Carbohydrates (g) | 282.58 ± 87.27 | 275.25 ± 68.78 | 0.681 | 286.73 ± 95.85 | 300.63 ± 98.06 | 0.306 | 0.926 | 0.573 |
| Protein (g) | 130.78 ± 31.03 | 128.76 ± 27.30 | 0.700 | 127.05 ± 24.61 | 148.76 ± 27.30 | 0.018 | 0.779 | 0.202 |
| Fat (g) | 116.01 ± 34.51 | 111.41 ± 37.05 | 0.682 | 103.23 ± 29.18 | 106.63 ± 37.74 | 0.748 | 0.407 | 0.791 |
| Fiber (g) | 21.26 ± 5.65 | 20.91 ± 4.57 | 0.576 | 20.91 ± 4.57 | 19.34 ± 3.46 | 0.496 | 0.607 | 0.419 |
| Carbohydrates (%) of energy | 41.37 ± 4.10 | 41.82 ± 4.92 | 0.812 | 43.80 ± 5.13 | 42.76 ± 5.42 | 0.607 | 0.294 | 0.709 |
| Protein (%) of energy | 19.62 ± 2.87 | 20.33 ± 2.14 | 0.414 | 20.00 ± 2.53 | 22.57 ± 3.09 | 0.003 | 0.773 | 0.102 |
| Fat (%) of energy | 38.87 ± 4.91 | 39.00 ± 4.97 | 0.949 | 36.00 ± 4.18 | 36.60 ± 4.95 | 0.749 | 0.199 | 0.319 |
| Carbohydrates (g/kg bw) | 4.03 ± 1.16 | 3.97 ± 0.96 | 0.811 | 4.20 ± 1.30 | 4.45 ± 1.40 | 0.642 | 0.783 | 0.416 |
| Protein (g/kg bw) | 1.86 ± 0.39 | 1.84 ± 0.34 | 0.809 | 1.87 ± 0.38 | 2.20 ± 0.39 | 0.008 | 0.960 | 0.059 |
| Fat (g/kg bw) | 1.66 ± 0.52 | 1.59 ± 0.49 | 0.656 | 1.52 ± 0.44 | 1.59 ± 0.51 | 0.642 | 0.550 | 0.998 |
| Fiber (g/kg bw) | 0.31 ± 0.09 | 0.30 ±0.08 | 0.757 | 0.29 ± 0.08 | 0.29 ± 0.06 | 0.760 | 0.797 | 0.719 |
| Carbohydrates (kcal/kg bw) | 16.14 ± 4.64 | 14.73 ± 3.82 | 0.811 | 16.80 ± 5.20 | 17.83 ± 5.60 | 0.642 | 0.783 | 0.416 |
| Protein (kcal/kg bw) | 7.46 ± 1.57 | 7.39 ± 1.37 | 0.809 | 7.49 ± 1.50 | 8.81 ± 1.54 | 0.008 | 0.960 | 0.059 |
| Fat (kcal/kg bw) | 14.96 ± 4.76 | 14.34 ± 4.39 | 0.656 | 13.70 ± 3.96 | 14.33 ± 4.62 | 0.642 | 0.550 | 0.998 |
| SCFA (µg/g) | CHO | PRO | ||||
|---|---|---|---|---|---|---|
| t = 0 | t = 10 Weeks | p | t = 0 | t = 10 Weeks | p | |
| Acetic acid | 1425.30 ± 51.89 | 1532.25 ± 390.71 | 0.674 | 1493.69 ± 531.10 | 1379.55 ± 431.30 | 0.721 |
| Propionic acid | 894.11 ± 338.79 | 791.95 ± 420.49 | 0.401 | 949.78 ± 521.16 | 776.71 ± 349.31 | 0.575 |
| Butyric acid | 1169.94 ± 532.48 | 1084.52 ± 650.59 | 0.484 | 1240.91 ± 888.33 | 957.04 ± 474.89 | 0.241 |
| Isobutyric acid | 119.31 ± 48.54 | 90.85 ± 69.04 | 0.779 | 119.88 ± 47.11 | 111.87 ± 61.57 | 0.508 |
| Valeric acid | 210.43 ± 111.84 | 158.81 ± 96.25 | 0.484 | 171.12 ± 79.78 | 158.00 ± 85.92 | 0.285 |
| Isovaleric acid | 213.10 ± 94.33 | 162.51 ± 145.33 | 0.674 | 222.44 ± 99.41 | 212.55 ± 132.22 | 0.203 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moreno-Pérez, D.; Bressa, C.; Bailén, M.; Hamed-Bousdar, S.; Naclerio, F.; Carmona, M.; Pérez, M.; González-Soltero, R.; Montalvo-Lominchar, M.G.; Carabaña, C.; et al. Effect of a Protein Supplement on the Gut Microbiota of Endurance Athletes: A Randomized, Controlled, Double-Blind Pilot Study. Nutrients 2018, 10, 337. https://doi.org/10.3390/nu10030337
Moreno-Pérez D, Bressa C, Bailén M, Hamed-Bousdar S, Naclerio F, Carmona M, Pérez M, González-Soltero R, Montalvo-Lominchar MG, Carabaña C, et al. Effect of a Protein Supplement on the Gut Microbiota of Endurance Athletes: A Randomized, Controlled, Double-Blind Pilot Study. Nutrients. 2018; 10(3):337. https://doi.org/10.3390/nu10030337
Chicago/Turabian StyleMoreno-Pérez, Diego, Carlo Bressa, María Bailén, Safa Hamed-Bousdar, Fernando Naclerio, Manuel Carmona, Margarita Pérez, Rocío González-Soltero, Maria Gregoria Montalvo-Lominchar, Claudia Carabaña, and et al. 2018. "Effect of a Protein Supplement on the Gut Microbiota of Endurance Athletes: A Randomized, Controlled, Double-Blind Pilot Study" Nutrients 10, no. 3: 337. https://doi.org/10.3390/nu10030337
APA StyleMoreno-Pérez, D., Bressa, C., Bailén, M., Hamed-Bousdar, S., Naclerio, F., Carmona, M., Pérez, M., González-Soltero, R., Montalvo-Lominchar, M. G., Carabaña, C., & Larrosa, M. (2018). Effect of a Protein Supplement on the Gut Microbiota of Endurance Athletes: A Randomized, Controlled, Double-Blind Pilot Study. Nutrients, 10(3), 337. https://doi.org/10.3390/nu10030337