Regulation of Muscle Glycogen Metabolism during Exercise: Implications for Endurance Performance and Training Adaptations

{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
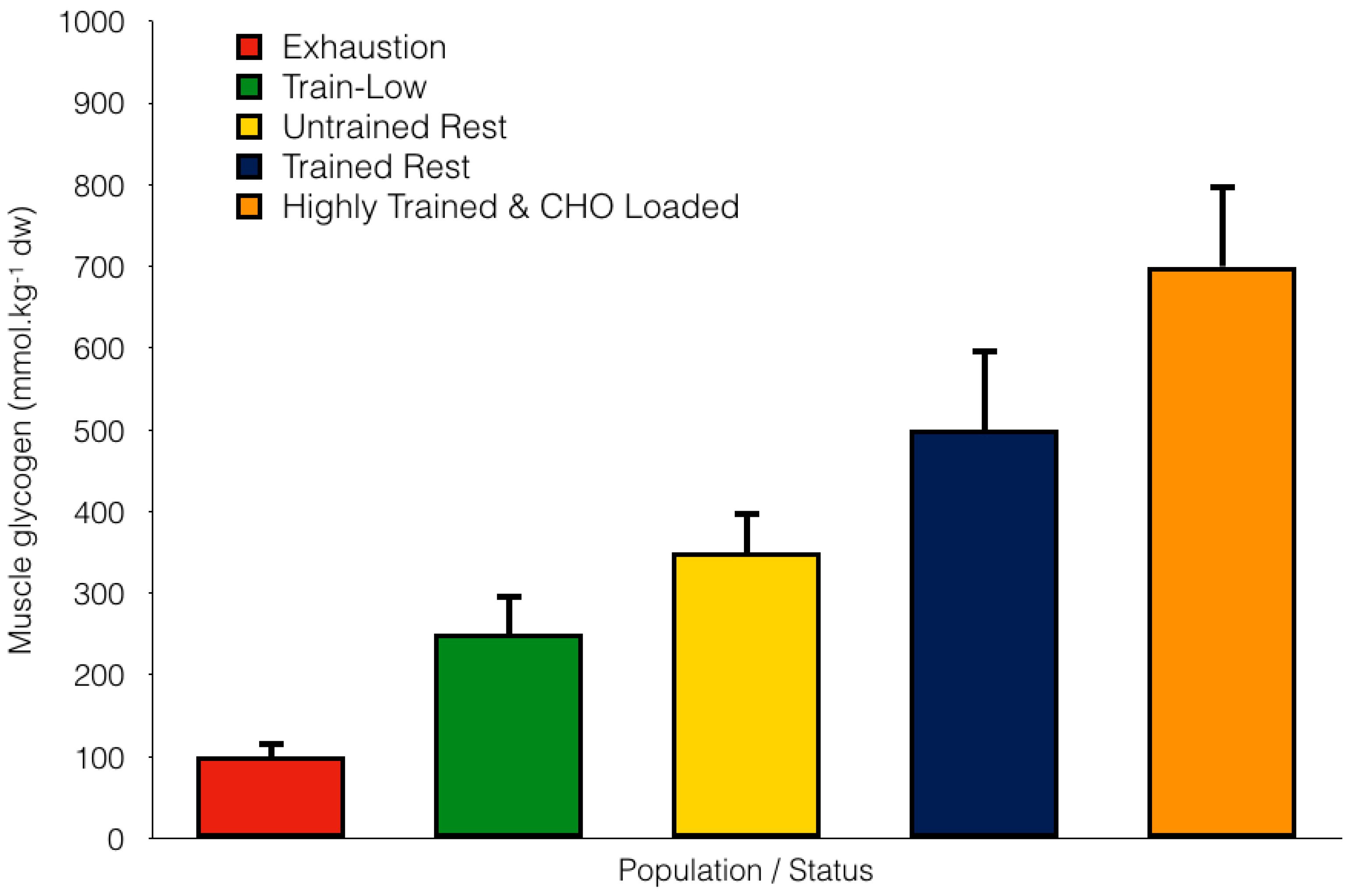
2. Overview of CHO Storage
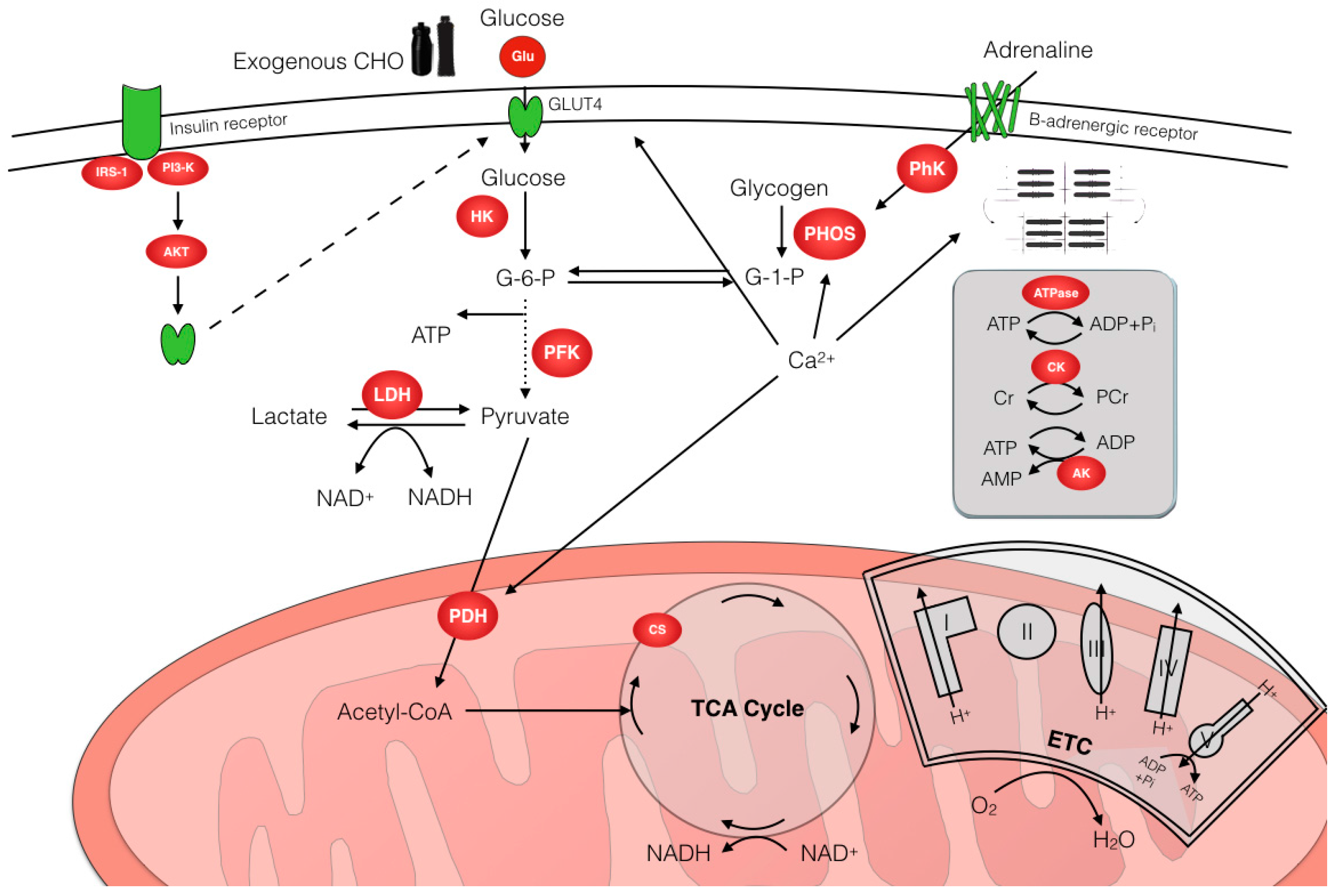
3. Regulation of CHO Metabolism
3.1. Effects of Exercise Intensity and Duration
3.2. Effects of Substrate Availability
3.3. Effects of Training Status
4. CHO and Exercise Performance
4.1. Muscle Glycogen and CHO Loading
4.2. Pre-Exercise CHO Availability
4.3. CHO Feeding during Exercise
5. CHO and Training Adaptations
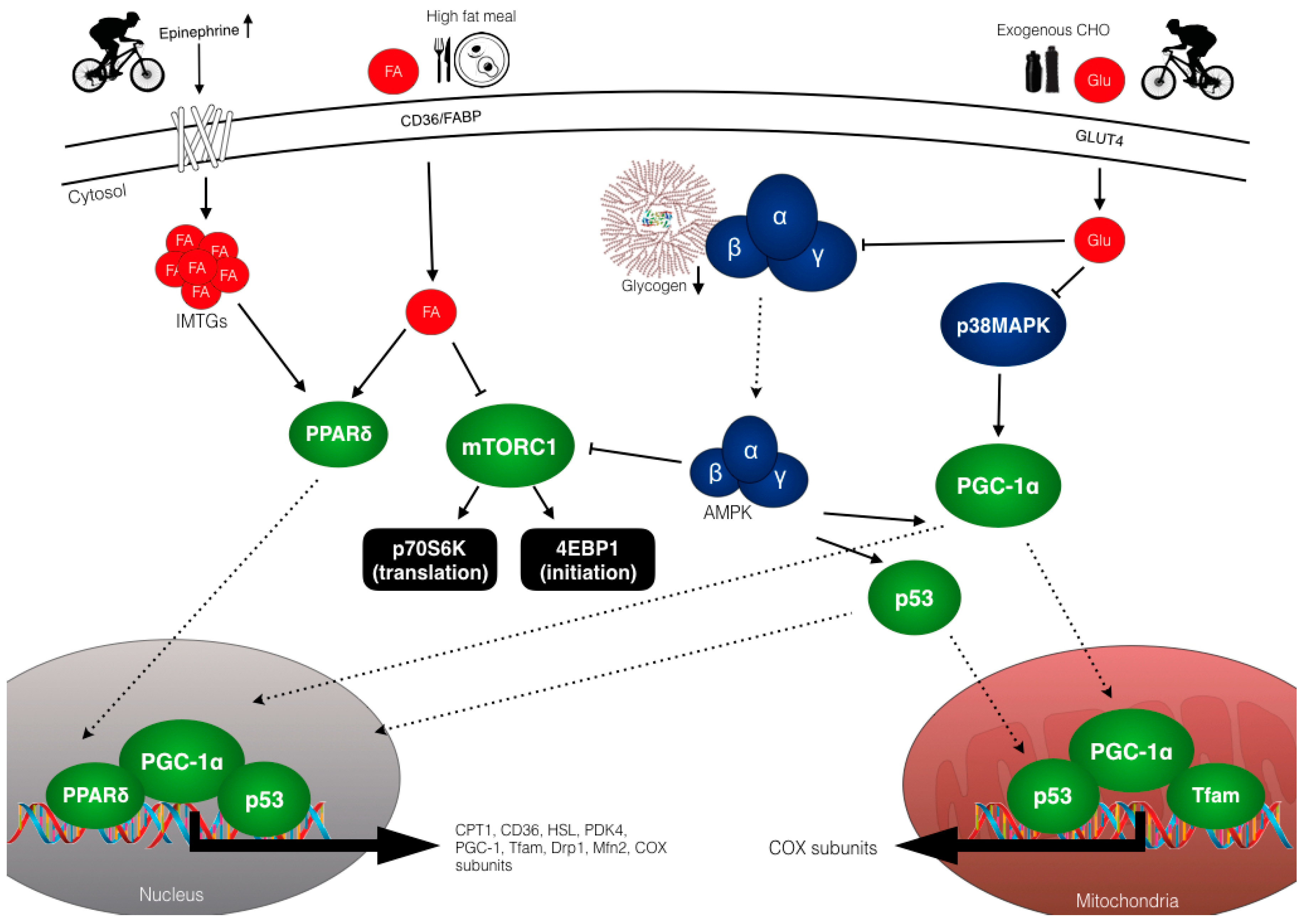
5.1. Overview of Molecular Regulation of Training Adaptations
5.2. Low Muscle Glycogen Availability and Twice per Day Training Models
5.3. Fasted Training
5.4. Post-Exercise CHO Restriction
5.5. Sleep-Low, Train-Low Models
5.6. Critical Limitations of Train-Low Models
5.7. Practical Applications
6. Summary and Future Directions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Krogh, A.; Lindhard, J. The relative value of fat and carbohydrate as sources of muscular energy: With appendices on the correlation between standard metabolism and the respiratory quotient during rest and work. Biochem. J. 1920, 14, 290–363. [Google Scholar] [CrossRef] [PubMed]
- Levine, S.A.; Gordon, B.; Derick, C.L. Some changes in the chemical constituents of the blood following a marathon race. J. Am. Med. Assoc. 1924, 82, 1778–1779. [Google Scholar] [CrossRef]
- Bergstrom, J.; Hermansen, L.; Hultman, E.; Saltin, B. Diet, muscle glycogen and physical performance. Acta Physiol. Scand. 1967, 71, 140–150. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, J.; Hultman, E. Muscle glycogen synthesis after exercise: An enhancing factor localized to the muscle cells in man. Nature 1966, 210, 309–310. [Google Scholar] [CrossRef] [PubMed]
- Bergstrom, J.; Hultman, E. The effect of exercise on muscle glycogen and electrolytes in normals. Scand. J. Clin. Lab. Investig. 1966, 18, 16–20. [Google Scholar] [CrossRef]
- Hermansen, L.; Hultman, E.; Saltin, B. Muscle glycogen during prolonged severe exercise. Acta Physiol. Scand. 1967, 71, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Thomas, D.T.; Erdman, K.A.; Burke, L.M. Position of the academy of nutrition and dietetics, dietitians of Canada, and the American College of sports medicine: Nutrition and athletic performance. J. Acad. Nutr. Diet. 2016, 116, 501–528. [Google Scholar] [CrossRef] [PubMed]
- Coyle, E.F.; Coggan, A.R.; Hemmert, M.K.; Ivy, J.L. Muscle glycogen utilization during prolonged strenuous exercise when fed carbohydrate. J. Appl. Physiol. 1986, 61, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Tsinztas, K.; Williams, C.; Boobis, L.; Greenhaff, P. Carbohydrate ingestion and glycogen utilization in different muscle fibre types in man. J. Physiol. 1995, 489, 243–250. [Google Scholar]
- Bosch, A.N.; Dennis, S.C.; Noakes, T.D. Influence of carbohydrate ingestion on fuel substrate turnover and oxidation during prolonged exercise. J. Appl. Physiol. 1994, 76, 2364–2372. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.; Brouns, F.; Wagenmakers, A.J.; Saris, W.H. Carbohydrate-electrolyte feedings improve 1 h time trial cycling performance. Int. J. Sports Med. 1997, 18, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A.E.; Jentjens, R. Oxidation of carbohydrate feedings during prolonged exercise: Current thoughts, guidelines and directions of future research. Sports Med. 2000, 29, 407–424. [Google Scholar] [CrossRef] [PubMed]
- Casey, A.; Mann, R.; Banister, K.; Fox, J.; Morris, P.G.; Macdonald, I.A.; Greenhaff, P.L. Effect of carbohydrate ingestion on glycogen resynthesis in human liver and skeletal muscle, measured by (13) C MRS. Am. J. Physiol. 2000, 278, E65–E75. [Google Scholar] [CrossRef]
- Carter, J.M.; Jeukendrup, A.E.; Jones, D.A. The effect of carbohydrate mouth rinse on 1-h cycle time trial performance. Med. Sci. Sports Exerc. 2004, 36, 2107–2111. [Google Scholar] [CrossRef] [PubMed]
- Chambers, E.S.; Bridge, M.W.; Jones, D.A. Carbohydrate sensing in the human mouth: Effects on exercise performance and brain activity. J. Physiol. 2009, 587, 1779–1794. [Google Scholar] [CrossRef] [PubMed]
- Philp, A.; Hargreaves, M.; Baar, K. More than a store: Regulatory roles for glycogen in skeletal muscle adaptation to exercise. Am. J. Physiol. Endocrinol. Metab. 2012, 302, E1343–E1351. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.D.; Hawley, J.A.; Morton, J.P. Carbohydrate availability and exercise training adaptation: Too much of a good thing? Eur. J. Sport Sci. 2015, 15, 3–12. [Google Scholar] [CrossRef] [PubMed]
- Graham, T.E.; Yuan, Z.; Hill, A.K.; Wilson, R.J. The regulation of muscle glycogen: The granule and its proteins. Acta Physiol. 2010, 199, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Marchand, I.; Chorneyko, K.; Tarnopolsky, M.; Hamilton, S.; Shearer, J.; Potvin, J.; Graham, T.E. Quantification of subcellular glycogen in resting human muscle: Granule size, number, and location. J. Appl. Physiol. 2002, 93, 1598–1607. [Google Scholar] [CrossRef] [PubMed]
- Essen, B.; Henriksson, J. Glycogen content of individual muscle fibres in man. Acta Physiol. Scand. 1974, 90, 645–647. [Google Scholar] [CrossRef] [PubMed]
- Essen, B.; Jansson, E.; Henriksson, J.; Taylor, AW.; Saltin, B. Metabolic characteristics of fibre types in human skeletal muscle. Acta Physiol. Scand. 1975, 95, 153–165. [Google Scholar] [CrossRef] [PubMed]
- Stellingwerff, T.; Boon, H.; Gijsen, A.P.; Stegen, J.H.; Kuipers, H.; van Loon, L.J. Carbohydrate supplementation during prolonged cycling exercise spares muscle glycogen but does not affect intramyocellular lipid use. Pflug. Arch. 2007, 454, 635–647. [Google Scholar] [CrossRef] [PubMed]
- Tsintzas, O.K.; Williams, C.; Boobis, L.; Greenhaff, P. Carbohydrate ingestion and single muscle fibre glycogen metabolism during prolonged running in men. J. Appl. Physiol. 1996, 81, 801–809. [Google Scholar] [CrossRef] [PubMed]
- Gollnick, P.D.; Piehl, K.; Saltin, B. Selective glycogen depletion pattern in human muscle fibres after exercise of varying intensity and at varying pedalling rates. J. Physiol. 1974, 24, 45–57. [Google Scholar] [CrossRef]
- Krustrup, P.; Mohr, M.; Steensberg, A.; Bencke, J.; Kjaer, M.; Bangsbo, J. Muscle and blood metabolites during a soccer game: Implications for sprint performance. Med. Sci. Sports Exerc. 2006, 38, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, J.; Holmberg, H.C.; Schroder, H.D.; Saltin, B.; Ortenblad, N. Human skeletal muscle glycogen utilization in exhaustive exercise: Role of subcellular localization and fiber type. J. Physiol. 2011, 589, 2871–2885. [Google Scholar] [CrossRef] [PubMed]
- Marchand, I.; Tarnopolsky, M.; Adamo, K.B.; Bourgeois, J.M.; Chorneyko, K.; Graham, T.E. Quantitative assessment of human muscle glycogen granules size and number in subcellular locations during recovery from prolonged exercise. J. Physiol. 2007, 580, 617–628. [Google Scholar] [CrossRef] [PubMed]
- Ortenblad, N.; Nielsen, J.; Saltin, B.; Holmberger, H.C. Role of glycogen availability in sarcoplasmic reticulum Ca2+ kinetics in human skeletal muscle. J. Physiol. 2011, 589, 711–725. [Google Scholar] [CrossRef] [PubMed]
- Gejl, K.D.; Hvid, L.O.; Frandsen, U.; Jensen, K.; Sahlin, K.; Ortenblad, N. Muscle glycogen content modifies SR Ca2+ release rate in elite endurance athletes. Med. Sci. Sports Exerc. 2014, 46, 496–505. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.; Bartlett, J.D.; van de Graaf, C.S.; Louhelainen, J.; Coyne, V.; Iqbal, Z.; MacLaren, D.P.; Gregson, W.; Close, G.L.; Morton, J.P. Protein ingestion does not impair exercise-induced AMPK signalling when in a glycogen-depleted state: Implications for train-low compete-high. Eur. J. Appl. Physiol. 2013, 113, 1457–1468. [Google Scholar] [CrossRef] [PubMed]
- Bartlett, J.D.; Louhelainen, J.; Iqbal, Z.; Cochran, A.J.; Gibala, M.J.; Gregson, W.; Close, G.L.; Drust, B.; Morton, J.P. Reduced carbohydrate availability enhances exercise-induced p53 signalling in human skeletal muscle: Implications for mitochondrial biogenesis. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Arkinstall, M.J.; Bruce, C.R.; Nikolopoulos, V.; Garnham, A.P.; Hawley, J.A. Effect of carbohydrate ingestion of metabolism during running and cycling. J. Appl. Physiol. 2001, 91, 2125–2134. [Google Scholar] [CrossRef] [PubMed]
- Van Loon, L.J.; Greenhaff, P.L.; Constantin-Teodosiu, D.; Saris, W.H.; Wagenmakers, A.J. The effects of increasing exercise intensity on muscle fuel utilisation in humans. J. Physiol. 2001, 536, 295–304. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, J.T.; Fuchs, C.J.; Betts, J.A.; van Loon, L.J. Liver glycogen metabolism during and after prolonged endurance-type exercise. Am. J. Physiol. Endocrinol. Metab. 2016, 311, E543–E553. [Google Scholar] [CrossRef] [PubMed]
- Achten, J.; Jeukendrup, A. Optimising fat oxidation through exercise and diet. Nutrition 2004, 20, 716–727. [Google Scholar] [CrossRef] [PubMed]
- Picton, C.; Klee, C.B.; Cohen, P. The regulation of muscle phosphorylase kinase by calcium ions, calmodulin and troponin-C. Cell Calcium 1981, 2, 281–294. [Google Scholar] [CrossRef]
- Howlett, R.A.; Parolin, M.L.; Dyck, D.J.; Jones, N.L.; Heigenhauser, G.J.; Spriet, L.L. Regulation of skeletal muscle glycogen phosphorylase and PDH at varying exercise power outputs. Am. J. Physiol. 1998, 275, R418–R425. [Google Scholar] [CrossRef] [PubMed]
- Parolin, M.L.; Chesley, A.; Matsos, M.P.; Spriet, L.L.; Jones, N.L.; Heigenhauser, G.J.F. Regulation of skeletal muscle glycogen phosphorylase and PDH during maximal intermittent exercise. Am. J. Physiol. 1999, 277, E890–E900. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.J.; Richter, E.A. Skeletal muscle glucose uptake during exercise: How is it regulated. J. Physiol. 2005, 20, 260–270. [Google Scholar] [CrossRef] [PubMed]
- Trimmer, J.K.; Schwarz, J.M.; Casazza, G.A.; Horning, M.A.; Rodriguez, N.; Brooks, G.A. Measurement of gluconeogenesis in exercising men by mass isotopomer distribution analysis. J. Appl. Physiol. 2002, 93, 233–241. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Kraniou, G.N.; Cameron-Smith, D.; Hargreaves, M. Acute exercise and GLUT4 expression in human skeletal muscle: Influence of exercise intensity. J. Appl. Physiol. 2006, 101, 934–937. [Google Scholar] [CrossRef] [PubMed]
- Martin, I.K.; Katz, A.; Wahren, J. Splanchnic and muscle metabolism during exercise in NIDDM patients. Am. J. Physiol. 1995, 269, E583–E590. [Google Scholar] [CrossRef] [PubMed]
- Katz, A.; Broberg, S.; Sahlin, K.; Wahren, J. Leg glucose uptake during maximal dynamic exercise in humans. Am. J. Physiol. 1986, 251, E65–E70. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J.; Heigenhauser, G.J.; Dyck, D.J.; Spriet, L.L. Intramuscular triacglycerol, glycogen, and acetyl group metabolism during 4 h of moderate exercise in man. J. Physiol. 2002, 541, 969–978. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J.; Heigenhauser, G.J.; LeBlanc, P.J.; Inglis, J.G.; Spriet, L.L.; Peters, S.J. Rapid upregulation of pyruvate dehydrogenase kinase activity in human skeletal muscle during prolonged exercise. J. Appl. Physiol. 2004, 97, 1261–1267. [Google Scholar] [CrossRef] [PubMed]
- Hargreaves, M.; McConell, G.; Proletto, J. Influence of muscle glycogen on glycogenolysis and glucose uptake during exercise in humans. J. Appl. Physiol. 1995, 78, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Arkinstall, M.J.; Bruce, C.R.; Clark, S.A.; Rickards, C.A.; Burke, L.M.; Hawley, J.A. Regulation of fuel metabolism by pre-exercise muscle glycogen content and exercise intensity. J. Appl. Physiol. 2004, 97, 2275–2283. [Google Scholar] [CrossRef] [PubMed]
- Kiilerich, K.; Gudmundsson, M.; Birk, J.B.; Lundby, C.; Taudorf, S.; Plomgard, P.; Saltin, B.; Pedersen, P.A.; Wojtaszewski, J.F.P.; Pilegaard, H. Low muscle glycogen and elevated plasma free fatty acid modify but do not prevent exercise-induced PDH activation in human skeletal muscle. Diabetes 2010, 59, 26–32. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Peters, S.J.; St Amand, T.A.; Howlett, R.A.; Heigenhauser, G.J.F.; Spriet, L.L. Human skeletal muscle pyruvate dehydrogenase kinase activity increases after a low carbohydrate diet. Am. J. Physiol. 1998, 275, E980–E986. [Google Scholar] [CrossRef] [PubMed]
- Roepstorff, C.; Halberg, N.; Hillig, T.; Saha, A.K.; Ruderman, N.B.; Wojtaszewski, J.F.P.; Richter, E.A.; Kiens, B. Malonyl CoA and carnitine in regulation of fat oxidation in human skeletal muscle during exercise. Am. J. Physiol. 2005, 288, E133–E142. [Google Scholar] [CrossRef] [PubMed]
- Wall, B.T.; Stephens, F.B.; Constantin-Teodosiu, D.; Marimuthu, K.; Macdonald, I.A.; Greenhaff, P.L. Chronic oral ingestion of L-carnitine and carbohydrate increases muscle carnitine content and alters muscle fuel metabolism during exercise in humans. J. Physiol. 2011, 589, 963–973. [Google Scholar] [CrossRef] [PubMed]
- Smith, B.K.; Perry, C.G.R.; Koves, T.R.; Wright, D.C.; Smith, J.C.; Neufer, D.P.; Muoio, D.M.; Holloway, G.P. Identification of a novel malonyl-CoA IC50 for CPT-1: Implications for predicting in vivo fatty acid oxidation rates. Biochem. J. 2012, 448, 13–20. [Google Scholar] [CrossRef] [PubMed]
- Gonzales, J.T.; Fuchs, C.J.; Smith, F.E.; Thelwall, P.E.; Taylor, R.; Stevenson, E.J.; Trenell, M.I.; Cermak, N.M.; Van Loon, L.J. Ingestion of glucose or sucrose prevents liver but not muscle glycogen depletion during prolonged endurance-type exercise in trained cyclists. Am. J. Physiol. Endocrinol. Metab. 2015, 309, E1032–E1039. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, J.F.; Mora-Rodriguez, R.; Byerley, L.O.; Coyle, E.F. Lipolytic suppression following carbohydrate ingestion limits fat oxidation during exercise. Am. J. Physiol. 1997, 273, E768–E775. [Google Scholar] [CrossRef] [PubMed]
- Coyle, E.F.; Jeukendrup, A.; Wagenmakers, A.J.; Saris, W.H. Fatty acid oxidation is directly regulated by carbohydrate metabolism during exercise. Am. J. Physiol. 1997, 273, E268–E275. [Google Scholar] [CrossRef] [PubMed]
- Watt, M.J.; Steinberg, G.R.; Chan, S.; Garnham, A.; Kemp, B.E.; Febbraio, M.A. Beta-adrenergic stimulation of skeletal muscle HSL can be overridden by AMPK signalling. FASEB J. 2004, 18, 1445–1446. [Google Scholar] [CrossRef] [PubMed]
- Henriksson, J. Training induced adaptation of skeletal muscle and metabolism during submaximal exercise. J. Physiol. 1977, 270, 661–675. [Google Scholar] [CrossRef] [PubMed]
- Karlsson, J.; Nordesjö, L.O.; Saltin, B. Muscle glycogen utilization during exercise after physical training. Acta Physiol. 1974, 90, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Saltin, B.; Nazar, K.; Costill, D.L.; Stein, E.; Jansson, E.; Essen, B.; Gollnick, P.D. The nature of the training response: Peripheral and central adaptations of one-legged exercise. Acta Physiol. Scand. 1976, 96, 289–305. [Google Scholar] [CrossRef] [PubMed]
- Chesley, A.; Heigenhauser, G.J.; Spriet, L.L. Regulation of glycogen phosphorylase activity following short term endurance training. Am. J. Physiol. 1996, 270, E328–E335. [Google Scholar] [CrossRef] [PubMed]
- Leblanc, P.J.; Howarth, K.R.; Gibala, M.J.; Heigenhauser, G.J. Effects of 7 weeks of endurance training on human skeletal muscle metabolism during submaximal exercise. J. Appl. Physiol. 2004, 97, 2148–2153. [Google Scholar] [CrossRef] [PubMed]
- Bergman, B.C.; Horning, M.A.; Casazza, G.A.; Wolfel, E.E.; Butterfiled, G.E.; Brooks, G.A. Endurance training increases gluconeogenesis during rest and exercise in men. Am. J. Physiol. Endocrinol. Metab. 2000, 278, E244–E251. [Google Scholar] [CrossRef] [PubMed]
- Bergman, B.C.; Butterfield, G.E.; Wolfel, E.E.; Lopaschuk, G.D.; Casazza, G.A.; Horning, M.A.; Brooks, G.A. Muscle net glucose uptake and glucose kinetics after endurance training in men. Am. J. Physiol. 1999, 277, E81–E92. [Google Scholar] [CrossRef] [PubMed]
- Richter, E.A.; Jensen, P.; Kiens, B.; Kristiansen, S. Sarcolemmal glucose transport and GLUT4 translocation during exercise are diminished by endurance training. Am. J. Physiol. 1998, 274, E89–E95. [Google Scholar] [PubMed]
- Phillips, S.M.; Green, H.J.; Tarnapolsky, M.A.; Heigenhauser, G.J.; Hill, R.E.; Grant, S.M. Effects of training duration on substrate turnover and oxidation during exercise. J. Appl. Physiol. 1996, 81, 2182–2191. [Google Scholar] [CrossRef] [PubMed]
- Sherman, W.M.; Costill, D.L.; Fink, W.J.; Miller, J.M. Effect of exercise-diet manipulation on muscle glycogen and its subsequent utilisation during performance. Int. J. Sports Med. 1981, 2, 114–118. [Google Scholar] [CrossRef] [PubMed]
- Bussau, V.A.; Fairchild, T.J.; Rao, A.; Steele, P.; Fournier, P.A. Carbohydrate loading in human muscle: An improved 1 day protocol. Eur. J. Appl. Physiol. 2002, 87, 290–295. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Collier, G.R.; Hargreaves, M. Muscle glycogen storage after prolonged exercise: Effect of the glycemic index of carbohydrate feedings. J. Appl. Physiol. 1993, 75, 1019–1023. [Google Scholar] [CrossRef] [PubMed]
- Hawley, J.A.; Schabort, E.J.; Noakes, T.D.; Dennis, S.C. Carbohydrate loading and exercise performance. Sports Med. 1997, 24, 73–81. [Google Scholar] [CrossRef] [PubMed]
- O’Brien, M.J.; Viguie, C.A.; Mazzeo, R.S.; Brooks, G.A. Carbohydrate dependence during marathon running. Med. Sci. Sports Exerc. 1993, 25, 1009–1017. [Google Scholar] [PubMed]
- Leckey, J.J.; Burke, L.M.; Morton, J.P.; Hawley, J.A. Altering fatty acid availability does not impair prolonged, continuous running to fatigue: Evidence for carbohydrate dependence. J. Appl. Physiol. 2016, 120, 107–113. [Google Scholar] [CrossRef] [PubMed]
- Ortenblad, N.; Westerblad, H.; Nielsen, J. Muscle glycogen stores and fatigue. J. Physiol. 2013, 591, 4405–4413. [Google Scholar] [CrossRef] [PubMed]
- Wee, L.-S.; Williams, C.; Tsintzas, K.; Boobis, L. Ingestion of a high glycemic index meal increases muscle glycogen storage at rest but augments its utilization during subsequent exercise. J. Appl. Physiol. 2005, 99, 707–714. [Google Scholar] [CrossRef] [PubMed]
- Sherman, W.M.; Peden, M.C.; Wright, D.A. Carbohydrate feedings 1 h before exercise improves cycling performance. Am. J. Clin. Nutr. 1991, 54, 866–870. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.A.; Sherman, W.M.; Dernbach, A.R. Carbohydrate feedings before, during or in combination improve cycling endurance performance. J. Appl. Physiol. 1991, 71, 1082–1088. [Google Scholar] [CrossRef] [PubMed]
- Stellingwerff, T.; Cox, G.R. Systematic review: Carbohydrate supplementation on exercise performance or capacity of varying durations. Appl. Physiol. Nutr. Metab. 2014, 39, 998–1011. [Google Scholar] [CrossRef] [PubMed]
- Jeukendrup, A. A step towards personalized sports nutrition: Carbohydrate intake during exercise. Sports Med. 2014, 44, S25–S33. [Google Scholar] [CrossRef] [PubMed]
- Newell, M.L.; Wallis, G.A.; Hunter, A.M.; Tipton, K.D.; Galloway, S.D.R. Metabolic responses to carbohydrate ingestion during exercise: Associations between carbohydrate dose and endurance performance. Nutrients 2018, 10, 37. [Google Scholar] [CrossRef] [PubMed]
- Carter, J.M.; Jeukendrup, A.E.; Mann, C.H.; Jones, D.A. The effect of glucose infusion on glucose kinetics during a 1-h time trial. Med. Sci. Sports Exerc. 2004, 36, 1543–1550. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Maughan, R.J. The Governor has a sweet tooth-mouth sensing of nutrients to enhance sports performance. Eur. J. Sport Sci. 2015, 15, 29–40. [Google Scholar] [CrossRef] [PubMed]
- Lane, S.C.; Bird, S.R.; Burke, L.M.; Hawley, J.A. Effect of a carbohydrate mouth rinse on simulated cycling time-trial performance commenced in a fed or fasted state. Appl. Physiol. Nutr. Metab. 2013, 38, 134–139. [Google Scholar] [CrossRef] [PubMed]
- Kasper, A.M.; Cocking, S.; Cockayne, M.; Barnard, M.; Tench, J.; Parker, L.; McAndrew, J.; Langan-Evans, C.; Close, G.L.; Morton, J.P. Carbohydrate mouth rinse and caffeine improves high-intensity interval running capacity when carbohydrate restricted. Eur. J. Sport Sci. 2015, 16, 560–568. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Yoo, M.J.; Moss, C.; Breier, B.H. Carbohydrate mouth rinsing has no effect on power output during cycling in a glycogen-reduced state. J. Int. Soc. Sports Nutr. 2016, 23, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.J.; Hammond, K.M.; Vasdev, A.; Poole, K.L.; Impey, S.G.; Close, G.L.; Morton, J.P. Self-selecting fluid intake while maintaining high carbohydrate availability does not impair half-marathon performance. Int. J. Sports Med. 2014, 35, 1216–1222. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, B.; Stellingwerff, T.; Zaltas, E.; Jeukendrup, A.E. CHO oxidation from a CHO gel compared with a drink during exercise. Med. Sci. Sports Exerc. 2010, 42, 2038–2045. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, B.; Stellingwerff, T.; Zaltas, E.; Jeukendrup, A.E. Oxidation of solid versus liquid CHO sources during exercise. Med. Sci. Sports Exerc. 2010, 42, 2030–2037. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; O’Kennedy, N.; Morton, J.P. Extreme Variation of Nutritional Composition and Osmolality of Commercially Available Carbohydrate Energy Gels. Int. J. Sport Nutr. Exerc. Metab. 2015, 25, 504–509. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, B.; Stellingwerff, T.; Hodgson, A.B.; Randell, R.; Pottgen, K.; Res, P.; Jeukendrup, A.E. Nutritional intake and gastrointestinal problems during competitive endurance events. Med. Sci. Sports Exerc. 2012, 44, 344–355. [Google Scholar] [CrossRef] [PubMed]
- Hills, S.P.; Russell, M. Carbohydrates for soccer: A focus on skilled actions and half-time practices. Nutrients 2018, 10, 22. [Google Scholar] [CrossRef] [PubMed]
- Holloszy, J.O.; Coyle, E.F. Adaptations of skeletal muscle to endurance exercise and their metabolic consequences. J. Appl. Physiol. 1984, 56, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Egan, B.; Zierath, J.B. Exercise metabolism and the molecular regulation of skeletal muscle adaptation. Cell Metab. 2013, 17, 162–184. [Google Scholar] [CrossRef] [PubMed]
- Wojtaszewski, J.F.; MacDonald, C.; Nielsen, J.N.; Hellsten, Y.; Hardie, D.G.; Kemp, B.E.; Kiens, B.; Richter, E.A. Regulation of 5′AMP-activated protein kinase activity and substrate utilization in exercising human skeletal muscle. Am. J. Physiol. Endocrinol. Metab. 2003, 284, 813–822. [Google Scholar] [CrossRef] [PubMed]
- Cochran, A.J.; Little, J.P.; Tarnopolsky, M.A.; Gibala, M.J. Carbohydrate feeding during recovery alters the skeletal muscle metabolic response to repeated sessions of high-intensity interval exercise in humans. J. Appl. Physiol. 2010, 108, 628–636. [Google Scholar] [CrossRef] [PubMed]
- Rose, A.J.; Frøsig, C.; Kiens, B.; Wojtaszewski, J.F.P.; Richter, E. Effect of endurance exercise training on Ca2+ calmodulin-dependent protein kinase II expression and signalling in skeletal muscle of humans. J. Physiol. 2007, 583, 785–795. [Google Scholar] [CrossRef] [PubMed]
- Perry, C.G.; Lally, J.; Holloway, G.P.; Heigenhauser, G.J.; Bonen, A.; Spriet, L.L. Repeated transient mRNA bursts precede increases in transcriptional and mitochondrial proteins during training in human skeletal muscle. J. Physiol. 2010, 588, 4795–4810. [Google Scholar] [CrossRef] [PubMed]
- Yeo, W.K.; Paton, C.D.; Garnham, A.P.; Burke, L.M.; Carey, A.L.; Hawley, J.A. Skeletal muscle adaptation and performance responses to once versus twice every second day endurance training regimens. J. Appl. Physiol. 2008, 105, 1462–1470. [Google Scholar] [CrossRef] [PubMed]
- Hulston, C.J.; Venables, M.C.; Mann, C.H.; Martin, C.; Philp, A.; Baar, K.; Jeukendrup, A.E. Training with low muscle glycogen enhances fat metabolism in well-trained cyclists. Med. Sci. Sports Exerc. 2010, 42, 2046–2055. [Google Scholar] [CrossRef] [PubMed]
- Hansen, A.K.; Fischer, C.P.; Plomgaard, P.; Andersen, J.L.; Saltin, B.; Pedersen, B.K. Skeletal muscle adaptation: Training twice every second day vs. training once daily. J. Appl. Physiol. 2005, 98, 93–99. [Google Scholar] [CrossRef] [PubMed]
- Marquet, L.A.; Brisswalter, J.; Louis, J.; Tiollier, E.; Burke, L.M.; Hawley, J.A.; Hausswirth, C. Enhanced endurance performance by periodization of carbohydrate intake: “Sleep low” strategy. Med. Sci. Sports Exerc. 2016, 48, 663–672. [Google Scholar] [CrossRef] [PubMed]
- Pilegaard, H.; Keller, C.; Steensberg, A.; Helge, J.W.; Pedersen, B.K.; Saltin, B.; Neufer, P.D. Influence of pre-exercise muscle glycogen content on exercise-induced transcriptional regulation of metabolic genes. J. Physiol. 2002, 541, 261–271. [Google Scholar] [CrossRef] [PubMed]
- Akerstrom, T.C.A.; Birk, J.B.; Klein, D.K.; Erikstrup, C.; Plomgaard, P.; Pedersen, B.K.; Wojtaszewski, J. Oral glucose ingestion attenuates exercise-induced activation of 5′-AMP-activated protein kinase in human skeletal muscle. Biochem. Biophys. Res. Commun. 2006, 342, 949–955. [Google Scholar] [CrossRef] [PubMed]
- Civitarese, A.E.; Hesselink, M.K.; Russell, A.P.; Ravussin, E.; Schrauwen, P. Glucose ingestion during exercise blunts exercise-induced gene expression of skeletal muscle fat oxidative genes. Am. J. Physiol. Endocrinol. Metab. 2005, 289, 1023–1029. [Google Scholar] [CrossRef] [PubMed]
- Cluberton, L.J.; McGee, S.L.; Murphy, R.M.; Hargreaves, M. Effect of carbohydrate ingestion on exercise-induced alterations in metabolic gene expression. J. Appl. Physiol. 2005, 99, 1359–1363. [Google Scholar] [CrossRef] [PubMed]
- Nybo, L.; Pedersen, K.; Christensen, B.; Aagaard, P.; Brandt, N.; Kiens, B. Impact of carbohydrate supplementation during endurance training on glycogen storage and performance. Acta Physiol. 2009, 197, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Van Proeyen, K.; Szlufcik, K.; Nielens, H.; Ramaekers, M.; Hespel, P. Beneficial metabolic adaptations due to endurance exercise training in the fasted state. J. Appl. Physiol. 2011, 110, 236–245. [Google Scholar] [CrossRef] [PubMed]
- Pilegaard, H.; Osada, T.; Andersen, L.T.; Helge, J.W.; Saltin, B.; Neufer, P.D. Substrate availability and transcriptional regulation of metabolic genes in human skeletal muscle during recovery from exercise. Metabolism 2005, 54, 1048–1055. [Google Scholar] [CrossRef] [PubMed]
- Morton, J.P.; Croft, L.; Bartlett, J.D.; Maclaren, D.P.; Reilly, T.; Evans, L.; McArdle, A.; Drust, B. Reduced carbohydrate availability does not modulate training-induced heat shock protein adaptations but does up regulate oxidative enzyme activity in human skeletal muscle. J. Appl. Physiol. 2009, 106, 1513–1521. [Google Scholar] [CrossRef] [PubMed]
- Lane, S.C.; Camera, D.M.; Lassiter, D.G.; Areta, J.L.; Bird, S.R.; Yeo, W.K.; Jeacocke, N.A.; Krook, A.; Zierath, J.R.; Burke, L.M.; et al. Effects of sleeping with reduced carbohydrate availability on acute training responses. J. Appl. Physiol. 2015, 119, 643–655. [Google Scholar] [CrossRef] [PubMed]
- McBride, A.; Ghilagaber, S.; Nikolaev, A.; Hardie, D.G. The glycogen-binding domain on the AMPK beta subunit allows the kinase to act as a glycogen sensor. Cell Metab. 2009, 9, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Chan, S.; McGee, S.L.; Watt, M.J.; Hargreaves, M.; Febbraio, M.A. Altering dietary nutrient intake that reduces glycogen content leads to phosphorylation of nuclear p38 MAP kinase in human skeletal muscle: Association with IL-6 gene transcription during contraction. FASEB J. 2004, 18, 1785–1787. [Google Scholar] [CrossRef] [PubMed]
- Impey, S.G.; Hearris, M.A.; Hammond, K.M.; Bartlett, J.D.; Louis, J.; Close, G.L.; Morton, J.P. Fuel for the work required: A theoretical framework for carbohydrate periodization and the glycogen threshold hypothesis. Sports Med. 2018. [Google Scholar] [CrossRef] [PubMed]
- Gejl, K.D.; Tharns, L.B.; Hansen, M.; Rokkedal-Lausch, T.; Plomgaard, P.; Nybo, L.; Larsen, F.J.; Cardinale, D.A.; Jensen, K.; Holmberg, H.C.; et al. No superior adaptations to carbohydrate periodization in elite endurance athletes. Med. Sci. Sports Exerc. 2017, 49, 2486–2497. [Google Scholar] [CrossRef] [PubMed]
- Burke, L.M.; Ross, M.L.; Garvican-Lewis, L.A.; Welvaert, M.; Heikura, I.A.; Forbes, S.G.; Mirtschin, J.G.; Cato, L.E.; Strobel, N.; Sharma, A.P.; et al. Low carbohydrate, high fat diet impairs exercise economy and negates performance benefit from intensified training in elite race walkers. J. Physiol. 2017, 595, 2785–2807. [Google Scholar] [CrossRef] [PubMed]
- Gleeson, M.; Nieman, D.C.; Pedersen, B.K. Exercise, nutrition and immune function. J. Sports Sci. 2004, 22, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Louis, J.; Marquet, L.A.; Tiollier, E.; Bernon, S.; Hausswirth, C.; Brisswalter, J. The impact of sleeping with reduced glycogen stores on immunity and sleep in triathletes. Eur. J. Appl. Physiol. 2016, 116, 1941–1954. [Google Scholar] [CrossRef] [PubMed]
- Howarth, K.R.; Phillips, S.M.; MacDonald, M.J.; Richards, D.; Moreau, N.A.; Gibala, M.J. Effect of glycogen availability on human skeletal muscle protein turnover during exercise and recovery. J. Appl. Physiol. 2010, 109, 431–438. [Google Scholar] [CrossRef] [PubMed]
- Cox, G.R.; Clark, S.A.; Cox, A.J.; Halson, S.L.; Hargreaves, M.; Hawley, J.A.; Burke, L.M. Daily training with high carbohydrate availability increases exogenous carbohydrate oxidation during endurance cycling. J. Appl. Physiol. 2010, 109, 126–134. [Google Scholar] [CrossRef] [PubMed]
- Impey, S.G.; Hammond, K.M.; Shepherd, S.O.; Sharples, A.P.; Stewart, C.; Limb, M.; Smith, K.; Jeromson, S.; Hamilton, D.L.; Close, G.L.; et al. Fuel for the work required: A practical approach to amalgamating train-low paradigms for endurance athletes. Physiol. Rep. 2016, 4, e12803. [Google Scholar] [CrossRef] [PubMed]



© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hearris, M.A.; Hammond, K.M.; Fell, J.M.; Morton, J.P. Regulation of Muscle Glycogen Metabolism during Exercise: Implications for Endurance Performance and Training Adaptations. Nutrients 2018, 10, 298. https://doi.org/10.3390/nu10030298
Hearris MA, Hammond KM, Fell JM, Morton JP. Regulation of Muscle Glycogen Metabolism during Exercise: Implications for Endurance Performance and Training Adaptations. Nutrients. 2018; 10(3):298. https://doi.org/10.3390/nu10030298
Chicago/Turabian StyleHearris, Mark A., Kelly M. Hammond, J. Marc Fell, and James P. Morton. 2018. "Regulation of Muscle Glycogen Metabolism during Exercise: Implications for Endurance Performance and Training Adaptations" Nutrients 10, no. 3: 298. https://doi.org/10.3390/nu10030298
APA StyleHearris, M. A., Hammond, K. M., Fell, J. M., & Morton, J. P. (2018). Regulation of Muscle Glycogen Metabolism during Exercise: Implications for Endurance Performance and Training Adaptations. Nutrients, 10(3), 298. https://doi.org/10.3390/nu10030298