Water Intake, Water Balance, and the Elusive Daily Water Requirement



Abstract
:1. Introduction
2. Representative Research Evidence
3. Why are Human Water Requirements Elusive?
- The relative influence of physiological processes which maintain water balance (Table 3) varies with different life scenarios. During sedentary daily activities in a mild environment, renal responses and thirst are the primary homeostatic regulators. During continuous-intermittent labor, or prolonged exercise at low intensities (5–18h duration), renal responses and thirst have minor-to-large effects on water regulation, whereas sweat loss presents the foremost challenge to homeostasis [56].
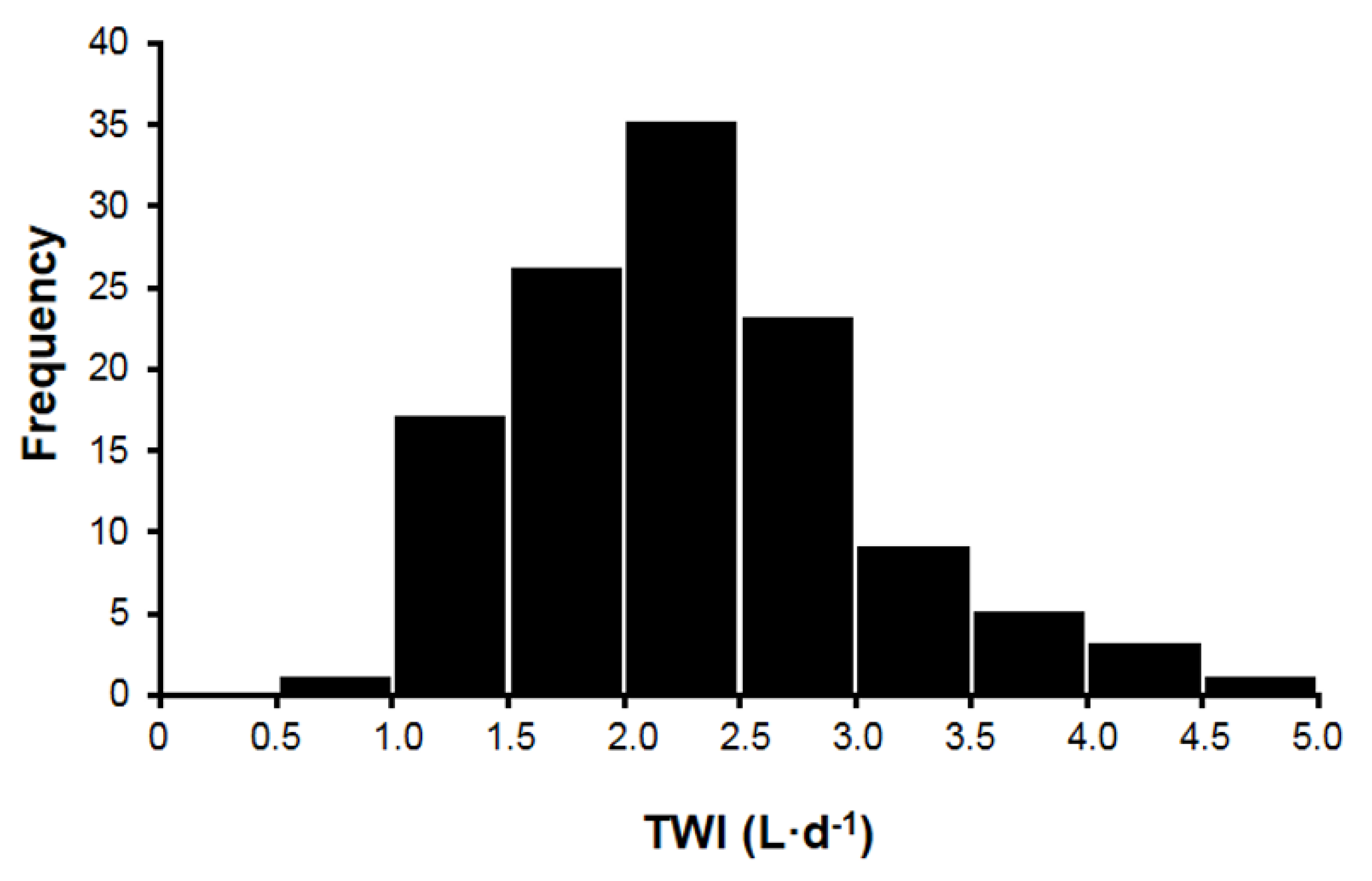
- Large between- and within-subject variances (i.e., of the variables in Table 3) make it difficult to determine a water requirement for all persons within a life stage (Table 1). As an example, Figure 2 illustrates the large between-subject variance of habitual TWI that exists in healthy young women (range, <1.0 to >4.5 L/24h) [13]. A large range of habitual TWI (0.6–5.2 L/24h) has also been reported for women during pregnancy [57]. Similarly, the third National Health and Nutrition Examination Survey [5] reported that the 1st decile and 10th decile of the mean TWI were 1.7–7.9 L/24h for men (n = 3,091) and 1.3–6.1 L/24h for women (n = 2,801). An example of large within-subject variability is also seen in the day-to-day differences of sweat losses that are experienced by athletes [24]. Total sweat loss during sedentary work activity (e.g., 8h of computer programming in an air-conditioned environment) may amount to <0.2 L/24h, whereas the total sweat volume during a 164-km ultradistance cycling event often exceeds 9 L during a 9-h ride [42].
- The 24-h human water requirement varies with anthropomorphic characteristics, especially body mass. Large individuals require a greater daily TWI than small individuals [6].
- The daily water requirement of any life-stage group is influenced by dietary sodium, protein and total solute load, due to individual dietary preferences as well as traditional regional-cultural foods. For example, large differences of mean urine osmolality (UOSM) have been reported for residents of Germany (860 mOsm/kg) and Poland (392 mOsm/kg). These differences are influenced by unique regional customs involving beverages (i.e., water, beer, wine) and food items [1] and the moisture content of solid foods; the latter factor varies among countries and demographic groups: the United States, 20–35% [2,51,58,59]; Germany, 27% [10]; the United Kingdom, 24–28%; and France, 35–38% [14].
- The principle that both water and beverages contribute to rehydration and the maintenance of body water has been fundamental in publications involving large populations [11,25], TWI differences in various countries [14,15], habitual low and high TWI consumers [16,17], water AI recommendations [6,7], the health effects of beverage consumption [60], young versus older adults [61], 12-h or 24-h water restriction [62,63], and experimental interventions which control and modify daily total water intake and beverage types [13,17,36,64]. However, small differences exist in the percentage of water retained (4-h post consumption), primarily due to beverage osmolality and the content of sodium chloride, protein, and/or energy [36,37].
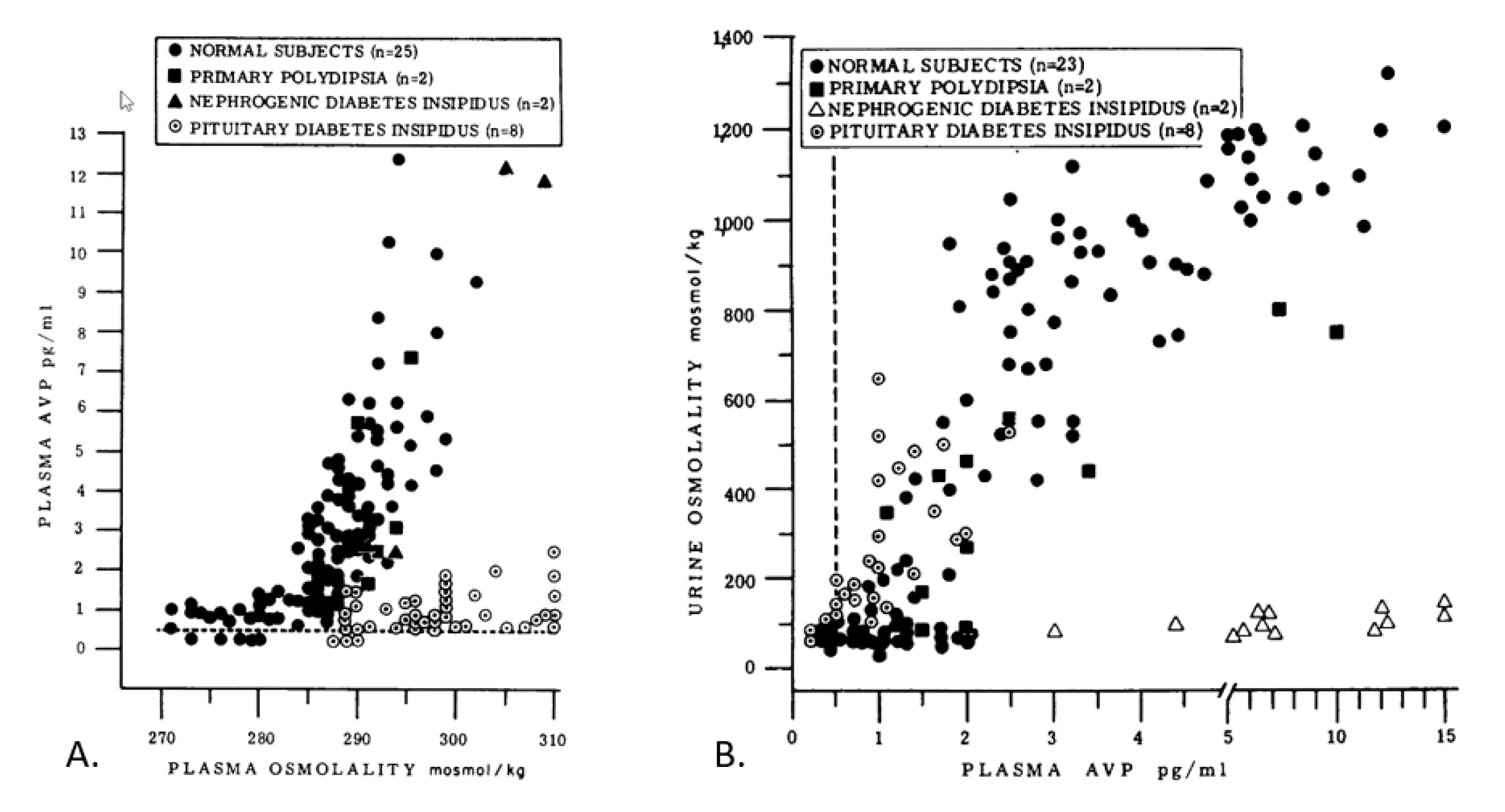
- Although some authorities consider plasma osmolality (POSM) to be the best index of euhydration and hypohydration [2,6], POSM does not assess whole-body hydration validly in all settings, especially when TBW, water intake, and water loss are fluctuating [66]. Furthermore, POSM may not reflect widely accepted physiologic principles, as shown by decreased POSM (6 out of 39 subjects) after losing 3–8% of body mass via sweating [67], and increased POSM at rest (4 out of 30 values) 60 min after ingesting 500 ml of water [68]. These findings likely result from the large between- and within-subject variance that exists in POSM measurements [56].
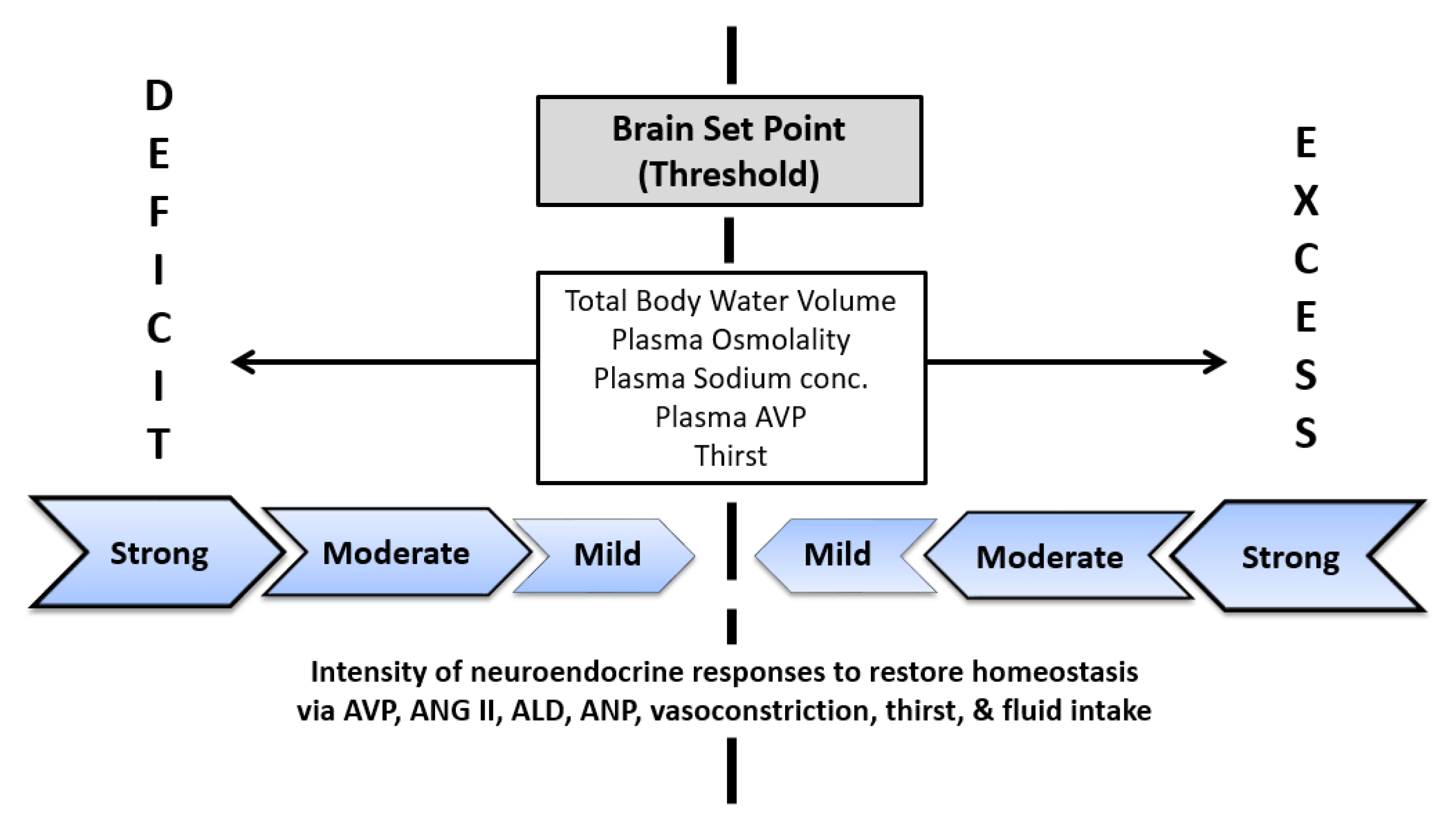
- Arginine vasopressin (AVP) is the body’s primary water-regulating hormone. It functions to maintain body water balance by keeping POSM within narrow limits and allowing the kidneys to alter water excretion in response to the body’s needs, in conjunction with thirst [69]. Dehydration of a large enough volume to result in increased POSM is a stimulus for the release of AVP. Table 4 summarizes research publications that determined the plasma osmotic threshold (i.e., set point) for increased plasma AVP; most of these studies employed intravenous hypertonic saline infusions with serial blood samples. Across these studies, the mean osmotic threshold values range from 280–288 and individual values range from 276–291 mOsm/kg. This large range of POSM values illustrates dynamic complexity, in that the network of fluid-regulatory functions, and water movements between fluid compartments differ across experimental designs and between normal subjects (see column 1, Table 4). Table 5 further describes the complexity of AVP, in terms of its biological functions, factors that influence neurohypohysial AVP release, and diseases which are related to AVP dysfunction.
- Thirst is the primary means by which humans sense dehydration and hypohydration. Several factors influence the onset of thirst, including blood pressure, blood volume, AVP, and angiotensin II [8]. The primary stimulus for thirst, however, is POSM. Table 6 summarizes research studies which determined the plasma osmotic threshold for the appearance of thirst. Across these studies, the mean osmotic threshold values range from 286–298 and individual values range from 276–300 mOsm/kg. As with AVP (see previous item), this large range of POSM values illustrates dynamic complexity, in that the network of fluid-regulatory functions and water movements between fluid compartments differ across experimental designs and among normal subjects (Table 6). This range of POSM values also may explain part of the range in habitual TWI (Figure 2).
- Older adults (>65 years) experience reduced thirst and water intake, reduced maximal renal concentrating ability, greater plasma AVP concentration during water restriction, and reduced ability to excrete a water load when compared to younger adults [61,88,89]. Although the osmotic threshold for thirst apparently does not change during the aging process [88,90], older adults have a reduced autonomic baroreceptor capability to sense a depletion of blood volume [89,91]. In addition, older adults demonstrate changes in water satiation that hinder the ability to hydrate following an osmotic challenge. This deficiency has been linked to changes in cerebral blood flow and/or altered activation of the anterior midcingulate cortex area within the brain [92]. Thus, aging appears to be responsible for large between-subject variances (i.e., of the variables in Table 3) across age groups, which make it difficult to determine a universal water requirement for children, adults, and the elderly (Table 1).
4. A Proposed Method to Assess Daily Water Requirements
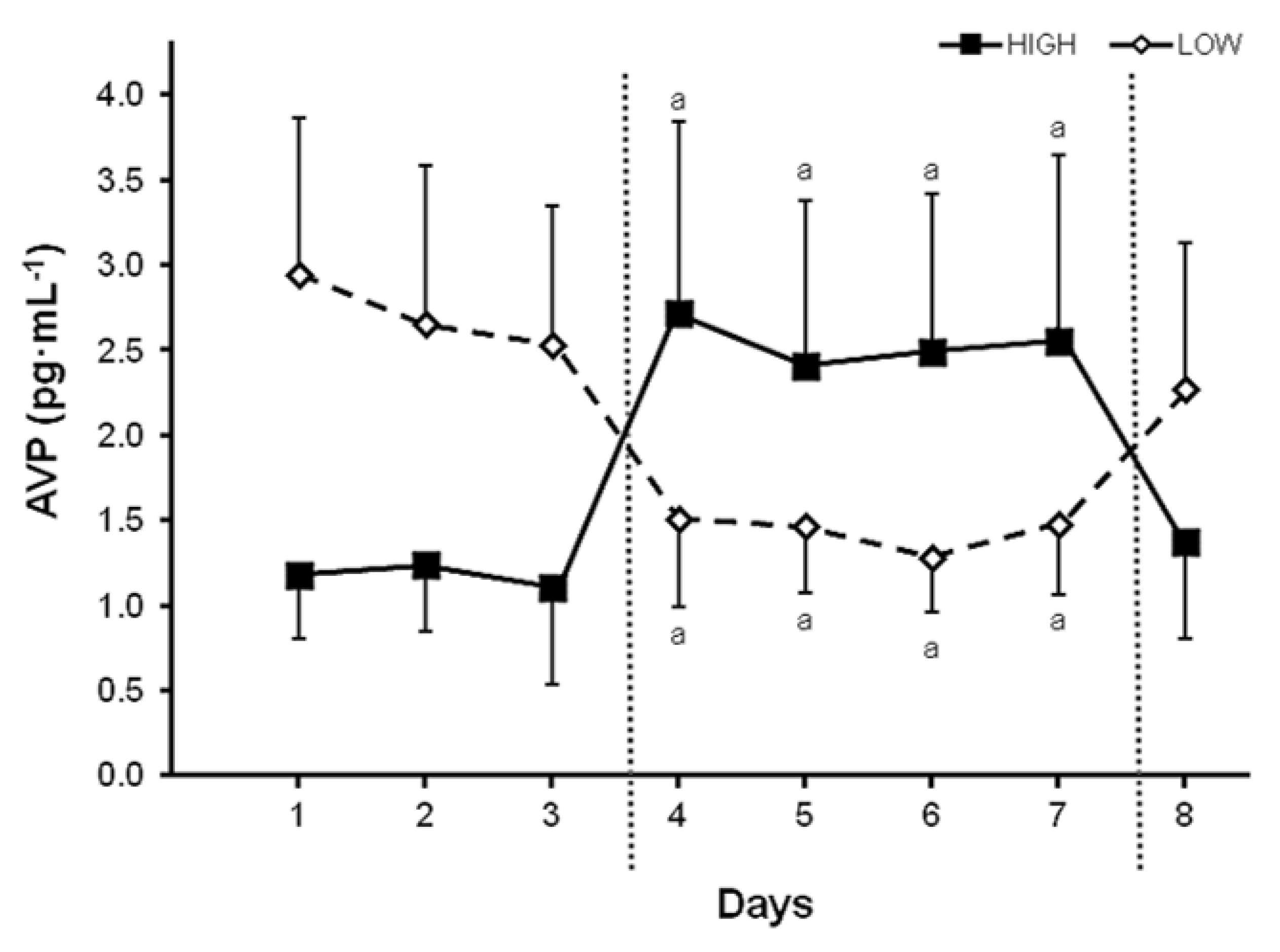
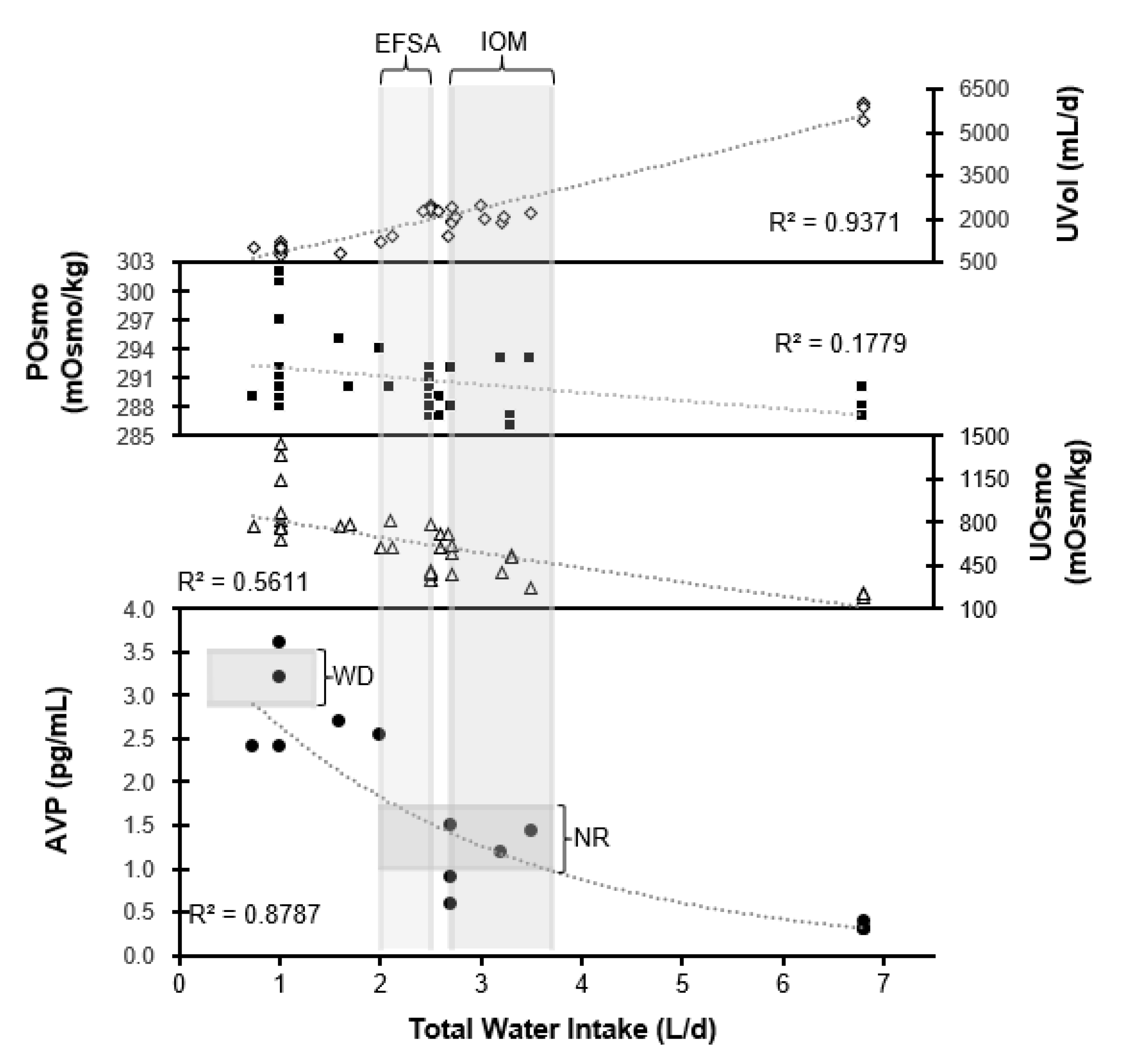
5. Neuroendocrine Responses across a Range of TWIs
6. Evidence for a Role of 24-h TWI in Reducing Disease Risk
7. AVP Influences Glucose Metabolism
8. Summary
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Manz, F.; Wentz, A. 24-h hydration status: Parameters, epidemiology and recommendations. Eur. J. Clin. Nutr. 2003, 57, S101–S108. [Google Scholar] [CrossRef] [PubMed]
- Sawka, M.N.; Cheuvront, S.N.; Carter, R. Human water needs. Nutr. Rev. 2005, 63, S303–S309. [Google Scholar] [CrossRef]
- Greenleaf, J.E. Problem: Thirst, drinking behavior, and involuntary dehydration. Med. Sci. Sports Exerc. 1992, 24, 645–656. [Google Scholar] [CrossRef] [PubMed]
- Kratz, A.; Ferraro, M.; Sluss, P.M.; Lewandrowski, K.B. Laboratory Reference Values. N. Engl. J. Med. 2004, 35, 1548–1563. [Google Scholar] [CrossRef] [PubMed]
- The third national health and nutrition examination survey (NHANES III 1988–1994). Available online: https://wwwn.cdc.gov/nchs/data/nhanes3/3a/VIFSE-acc.pdf (accessed on 25 May 2018).
- Institute of Medicine. Dietary reference intakes for water, potassium, sodium, chloride, and sulfate; Institute of Medicine: Washington, DC, USA, 2004. [Google Scholar]
- EFSA Panel on Dietic Products, Nutrition, and Allergies (NDA). Scientific opinion on dietary reference values for water. EFSA J. 2010, 8, 1459–1507. [Google Scholar]
- Thornton, S.N. Thirst and hydration: Physiology and consequences of dysfunction. Physiol. Behav. 2010, 100, 15–21. [Google Scholar] [CrossRef]
- Gamble, J. Chemical Anatomy, Physiology and Pathology of Extracellular Fluid: A Lecture Syllabus; Harvard University Press: Cambridge, MA, USA, 1947. [Google Scholar]
- Manz, F.; Johner, S.A.; Wentz, A.; Boeing, H.; Remer, T. Water balance throughout the adult life span in a German population. Br. J. Nutr. 2012, 107, 1673–1681. [Google Scholar] [CrossRef]
- Manz, F.; Wentz, A. Hydration status in the United States and Germany. Nutr. Rev. 2005, 63, S55–S62. [Google Scholar] [CrossRef]
- Manz, F.; Wentz, A.; Sichert-Hellert, W. The most essential nutrient: Defining the adequate intake of water. J. Pediatr. 2002, 141, 587–592. [Google Scholar] [CrossRef]
- Johnson, E.C.; Muñoz, C.X.; Jimenez, L.; Le Bellego, L.; Kupchak, B.R.; Kraemer, W.J.; Casa, D.J.; Maresh, C.M.; Armstrong, L.E. Hormonal and thirst modulated maintenance of fluid balance in young women with different levels of habitual fluid consumption. Nutrients 2016, 8, 302. [Google Scholar] [CrossRef]
- Guelinckx, I.; Tavoularis, G.; König, J.; Morin, C.; Gharbi, H.; Gandy, J. Contribution of water from food and fluids to total water intake: Analysis of a French and UK population surveys. Nutrients 2016, 8, 630. [Google Scholar] [CrossRef] [PubMed]
- Guelinckx, I.; Ferreira-Pêgo, C.; Moreno, L.A.; Kavouras, S.A.; Gandy, J.; Martinez, H.; Bardosono, S.; Abdollahi, M.; Nasseri, E.; Jarosz, A.; et al. Intake of water and different beverages in adults across 13 countries. Eur. J. Nutr. 2015, 54, 45–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perrier, E.; Vergne, S.; Klein, A.; Poupin, M.; Rondeau, P.; Le Bellego, L.; Armstrong, L.E.; Lang, F.; Stookey, J.; Tack, I. Hydration biomarkers in free-living adults with different levels of habitual fluid consumption. Br. J. Nutr. 2013, 109, 1678–1687. [Google Scholar] [CrossRef] [PubMed]
- Pross, N.; Demazières, A.; Girard, N.; Barnouin, R.; Metzger, D.; Klein, A.; Perrier, E.; Guelinckx, I. Effects of changes in water intake on mood of high and low drinkers. PLoS ONE 2014, 9, e94754. [Google Scholar] [CrossRef]
- Perrier, E.; Demazières, A.; Girard, N.; Pross, N.; Osbild, D.; Metzger, D.; Guelinckx, I.; Klein, A. Circadian variation and responsiveness of hydration biomarkers to changes in daily water intake. Eur. J. Appl. Physiol. 2013, 113, 2143–2151. [Google Scholar] [CrossRef] [Green Version]
- Lemetais, G.; Melander, O.; Vecchio, M.; Bottin, J.H.; Enhörning, S.; Perrier, E.T. Effect of increased water intake on plasma copeptin in healthy adults. Eur. J. Nutr. 2018, 57, 1883–1890. [Google Scholar] [CrossRef]
- Pross, N.; Demazières, A.; Girard, N.; Barnouin, R.; Santoro, F.; Chevillotte, E.; Klein, A.; Le Bellego, L. Influence of progressive fluid restriction on mood and physiological markers of dehydration in women. Br. J. Nutr. 2013, 109, 313–321. [Google Scholar] [CrossRef]
- National Health and Medical Research Council. National Health and Medical Research Council of Australia. In Australian Dietary Guidlines; National Health and Medical Research Council: Canberra, Australia, 2013. [Google Scholar]
- Armstrong, L.E.; Pumerantz, A.C.; Fiala, K.A.; Roti, M.W.; Kavouras, S.A.; Casa, D.J.; Maresh, C.M. Human hydration indices: Acute and longitudinal reference values. Int. J. Sport Nutr. Exerc. MeTable 2010, 20, 145–153. [Google Scholar] [CrossRef]
- Armstrong, L.E.; Johnson, E.C.; Muñoz, C.X.; Swokla, B.; Le Bellego, L.; Jimenez, L.; Casa, D.J.; Maresh, C.M. Hydration biomarkers and dietary fluid consumption of women. J. Acad. Nutr. Diet. 2012, 112, 1056–1061. [Google Scholar] [CrossRef]
- Leiper, J.B.; Pitsiladis, Y.; Maughan, R.J. Comparison of water turnover rates in men undertaking prolonged cycling exercise and sedentary men. Int. J. Sports Med. 2001, 22, 181–185. [Google Scholar] [CrossRef]
- Raman, A.; Schoeller, D.A.; Subar, A.F.; Troiano, R.P.; Schatzkin, A.; Harris, T.; Bauer, D.; Bingham, S.A.; Everhart, J.E.; Newman, A.B.; et al. Water turnover in 458 American adults 40–79 year of age. Am. J. Physiol. Renal Physiol. 2004, 286, F394–F401. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, A.L.; Armstrong, L.E. Monitoring body water balance in pregnant and nursing women: The validity of urine color. Ann. Nutr. Metab. 2017, 70, 18–22. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.E.; Kenefick, R.W.; Castellani, J.W.; Riebe, D.; Kavouras, S.A.; Kuznicki, J.T.; Maresh, C.M. Bioimpedance spectroscopy technique: Intra-, extracellular, and total body water. Med. Sci. Sports Exerc. 1997, 29, 1657–1663. [Google Scholar] [CrossRef] [PubMed]
- Davis, J.M.; Burgess, W.A.; Slentz, C.A.; Bartoli, W.P.; Pate, R.R. Effects of ingesting 6% and 12% glucose/electrolyte beverages during prolonged intermittent cycling in the heat. Eur. J. Appl. Physiol. Occup. Physiol. 1988, 57, 563–569. [Google Scholar] [CrossRef] [PubMed]
- Sawka, M.N.; Young, A.J.; Francesconi, R.P.; Muza, S.R.; Pandolf, K.B. Thermoregulatory and blood responses during exercise at graded hypohydration levels. J. Appl. Physiol. 1985, 59, 1394–1401. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.E.; Maresh, C.M.; Gabaree, C.V.; Hoffman, J.R.; Kavouras, S.A.; Kenefick, R.W.; Castellani, J.W.; Ahlquist, L.E. Thermal and circulatory responses during exercise: Effects of hypohydration, dehydration, and water intake. J. Appl. Physiol. 1997, 82, 2028–2035. [Google Scholar] [CrossRef] [PubMed]
- Kenefick, R.W.; Maresh, C.M.; Armstrong, L.E.; Castellani, J.W.; Riebe, D.; Echegaray, M.E.; Kavorous, S.A. Plasma vasopressin and aldosterone responses to oral and intravenous saline rehydration. J. Appl. Physiol. 2000, 89, 2117–2122. [Google Scholar] [CrossRef] [PubMed]
- Muñoz, C.X.; Johnson, E.C.; Demartini, J.K.; Huggins, R.A.; McKenzie, A.L.; Casa, D.J.; Maresh, C.M.; Armstrong, L.E. Assessment of hydration biomarkers including salivary osmolality during passive and active dehydration. Eur. J. Clin. Nutr. 2013, 67, 1257–1263. [Google Scholar] [CrossRef]
- Rolls, E.T.; Rolls, B.J.; Rowe, E.A. Sensory-specific and motivation-specific satiety for the sight and taste of food and water in man. Physiol. Behav. 1983, 30, 185–192. [Google Scholar] [CrossRef] [Green Version]
- Geelen, G.; Greenleaf, J.E.; Keil, L.C. Drinking-induced plasma vasopressin and norepinephrine changes in dehydrated humans. J. Clin. Endocrinol. Metab. 1996, 81, 2131–2135. [Google Scholar]
- Engell, D.B.; Maller, O.; Sawka, M.N.; Francesconi, R.N.; Drolet, L.; Young, A.J. Thirst and fluid intake following graded hypohydration levels in humans. Physiol. Behav. 1987, 40, 229–236. [Google Scholar] [CrossRef] [Green Version]
- Maughan, R.J.; Watson, P.; Cordery, P.A.; Walsh, N.P.; Oliver, S.J.; Dolci, A.; Rodriguez-Sanchez, N.; Galloway, S.D. A randomized trial to assess the potential of different beverages to affect hydration status: Development of a beverage hydration index. Am. J. Clin. Nutr. 2016, 103, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Evans, G.H.; James, L.J.; Shirreffs, S.M.; Maughan, R.J. Optimizing the restoration and maintenance of fluid balance after exercise-induced dehydration. J. Appl. Physiol. 2017, 122, 945–951. [Google Scholar] [CrossRef]
- Perrier, E.T.; Buendia-Jimenez, I.; Vecchio, M.; Armstrong, L.E.; Tack, I.; Klein, A. Twenty-four-hour urine osmolality as a physiological index of adequate water intake. Dis. Markers 2015, 2015, 231063. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, A.L.; Muñoz, C.X.; Armstrong, L.E. Accuracy of urine color to detect equal to or greater than 2% body mass loss in men. J Athl. Train 2015, 50, 1306–1309. [Google Scholar] [CrossRef] [PubMed]
- Romano, G.; Bortolotti, N.; Falleti, E.; Favret, G.; Gonano, F.; Bartoli, G.E. The influence of furosemide on free water clearance. Panminerva. Med. 1999, 41, 103–108. [Google Scholar] [PubMed]
- Armstrong, L.E.; Johnson, E.C.; McKenzie, A.L.; Muñoz, C.X. Interpreting common hydration biomarkers on the basis of solute and water excretion. Eur. J. Clin. Nutr. 2013, 67, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.E.; Johnson, E.C.; Ganio, M.S.; Judelson, D.A.; Vingren, J.L.; Kupchak, B.R.; Kunces, L.J.; Muñoz, C.X.; McKenzie, A.L.; Williamson, K.H. Effective body water and body mass changes during summer ultra-endurance road cycling. J. Sports Sci. 2015, 33, 125–135. [Google Scholar] [CrossRef] [PubMed]
- Maughan, R.J.; Shirreffs, S.M.; Merson, S.J.; Horswill, C.A. Fluid and electrolyte balance in elite male football (soccer) players training in a cool environment. J. Sports Sci. 2005, 23, 73–79. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehrer, N.J.; Burke, L.M. Sweat losses during various sports. Aust. J. Nutr. Diet. 1996, 53, S13–S16. [Google Scholar]
- Adolph, E.F. Physiology of Man in the Desert; Adolf, E.F., Ed.; Interscience Pub.: Cummings Park, MA, USA, 1947. [Google Scholar]
- Cheuvront, S.N.; Fraser, C.G.; Kenefick, R.W.; Ely, B.R.; Sawka, M.N. Reference change values for monitoring dehydration. Clin. Chem. Lab. Med. 2011, 49, 1033–1037. [Google Scholar] [CrossRef] [PubMed]
- Cheuvront, S.N.; Ely, B.R.; Kenefick, R.W.; Sawka, M.N. Biological variation and diagnostic accuracy of dehydration assessment markers. Am. J. Clin. Nutr. 2010, 92, 565–573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheuvront, S.N.; Kenefick, R.W. Dehydration: Physiology, assessment, and performance effects. Compr Physiol 2014, 4, 257–285. [Google Scholar] [PubMed]
- Racette, S.B.; Schoeller, D.A.; Luke, A.H.; Shay, K.; Hnilicka, J.; Kushner, R.F. Relative dilution spaces of 2H- and 18O-labeled water in humans. Am. J. Physiol. 1994, 267, E585–E590. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.E.; Maresh, C.M.; Castellani, J.; Bergeron, M.; Kenefick, R.; La Gasse, K.; Riebe, D. Urinary indices of hydration status. Int. J. Sport Nutr. 1994, 4, 265–279. [Google Scholar] [CrossRef] [PubMed]
- Johnson, E.C.; Muñoz, C.X.; Le Bellego, L.; Klein, A.; Casa, D.J.; Maresh, C.M.; Armstrong, L.E. Markers of the hydration process during fluid volume modification in women with habitual high or low daily fluid intakes. Eur. J. Appl. Physiol. 2015, 115, 1067–1074. [Google Scholar] [CrossRef]
- Maughan, R.J.; Shirreffs, S.M.; Leiper, J.B. Errors in the estimation of hydration status from changes in body mass. J. Sports Sci. 2007, 25, 797–804. [Google Scholar] [CrossRef] [Green Version]
- Hoyt, R.W.; Honig, A. Environmental influences on body fluid balance during exercise: Altitude. In Body Fluid Balance: Exercise and Sport; Buskirk, E., Puhl, S.M., Eds.; CRC: Boca Raton, FL, USA, 1996; pp. 183–196. [Google Scholar]
- Shore, A.C.; Markandu, N.D.; Sagnella, G.A.; Singer, D.R.; Forsling, M.L.; Buckley, M.G.; Sugden, A.L.; MacGregor, G.A. Endocrine and renal response to water loading and water restriction in normal man. Clin. Sci. 1988, 75, 171–177. [Google Scholar] [CrossRef]
- Miles, B.E.; Paton, A.; De Wardener, H.E. Maximum urine concentration. Br. Med. J. 1954, 2, 901–905. [Google Scholar] [CrossRef] [PubMed]
- Armstrong, L.E. Assessing hydration status: The elusive gold standard. J. Am. Coll. Nutr. 2007, 26, 575s–584s. [Google Scholar] [CrossRef] [PubMed]
- McKenzie, A.L.; Perrier, E.T.; Guelinckx, I.; Kavouras, S.A.; Aerni, G.; Lee, E.C.; Volek, J.S.; Maresh, C.M.; Armstrong, L.E. Relationships between hydration biomarkers and total fluid intake in pregnant and lactating women. Eur. J. Nutr. 2017, 56, 2161–2170. [Google Scholar] [CrossRef] [PubMed]
- Grandjean, A.C.; Reimers, K.J.; Haven, M.C.; Curtis, G.L. The effect on hydration of two diets, one with and one without plain water. J. Am. Coll. Nutr. 2003, 22, 165–173. [Google Scholar] [CrossRef]
- Tucker, M.A.; Ganio, M.S.; Adams, J.D.; Brown, L.A.; Ridings, C.B.; Burchfield, J.M.; Robinson, F.B.; McDermott, J.L.; Schreiber, B.A.; Moyen, N.E.; et al. Hydration status over 24-H is not affected by ingested beverage composition. J. Am. Coll. Nutr. 2015, 34, 318–327. [Google Scholar] [CrossRef] [PubMed]
- Popkin, B.M.; Armstrong, L.E.; Bray, G.M.; Caballero, B.; Frei, B.; Willett, W.C. A new proposed guidance system for beverage consumption in the United States. Am. J. Clin. Nutr. 2006, 83, 529–542. [Google Scholar] [CrossRef] [Green Version]
- Rolls, B.J.; Phillips, P.A. Aging and disturbances of thirst and fluid balance. Nutr. Rev. 1990, 48, 137–144. [Google Scholar] [CrossRef] [PubMed]
- Davison, J.M.; Gilmore, E.A.; Durr, J.; Robertson, G.L.; Lindheimer, M.D. Altered osmotic thresholds for vasopressin secretion and thirst in human pregnancy. Am. J. Physiol. 1984, 246, F105–F109. [Google Scholar] [CrossRef] [PubMed]
- Geelen, G.; Keil, L.C.; Kravik, S.E.; Wade, C.E.; Thrasher, T.N.; Barnes, P.R.; Pyka, G.; Nesvig, C.; Greenleaf, J.E. Inhibition of plasma vasopressin after drinking in dehydrated humans. Am. J. Physiol. 1984, 247, R968–R971. [Google Scholar] [CrossRef] [PubMed]
- Stookey, J.D.; Hamer, J.; Killilea, D.W. Change in hydration indices associated with an increase in total water intake of more than 0.5 L/day, sustained over 4 weeks, in healthy young men with initial total water intake below 2 L/day. Physiol. Rep. 2017. [Google Scholar] [CrossRef]
- Silverthorn, D.U. Human Physiology: An integrated approach, 5th ed.; Silverthorn, D.U., Ed.; Pearson/Benjamin Cummings: San Franciso, CA, USA, 2009. [Google Scholar]
- Armstrong, L.E.; Maughan, R.J.; Senay, L.C.; Shirreffs, S.M. Limitations to the use of plasma osmolality as a hydration biomarker. Am. J. Clin. Nutr. 2013, 98, 503–504. [Google Scholar] [CrossRef] [Green Version]
- Sawka, M.N.; Montain, S.J.; Latzka, W.A. Body fluid balance during exercise-heat exposure. In Body Fluid Balance: Exercise and Sport; CRC Press: Boca Raton, FL, USA, 1996; pp. 139–157. [Google Scholar]
- Sollanek, K.J.; Kenefick, R.W.; Cheuvront, S.N.; Axtell, R.S. Potential impact of a 500-mL water bolus and body mass on plasma osmolality dilution. Eur. J. Appl. Physiol. 2011, 111, 1999–2004. [Google Scholar] [CrossRef]
- Bankir, L.; Bichet, D.G.; Morgenthaler, N.G. Vasopressin: Physiology, assessment and osmosensation. J. Intern. Med. 2017, 282, 284–297. [Google Scholar] [CrossRef] [PubMed]
- Moses, A.M.; Miller, M. Osmotic threshold for vasopressin release as determined by saline infusion and by dehydration. Neuroendocrinology 1971, 7, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.L.; Mahr, E.A.; Athar, S.; Sinha, T. Development and clinical application of a new method for the radioimmunoassay of arginine vasopressin in human plasma. J. Clin. Invest. 1973, 52, 2340–2352. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.L.; Shelton, R.L.; Athar, S. The osmoregulation of vasopressin. Kidney Int. 1976, 10, 25–37. [Google Scholar] [CrossRef] [PubMed]
- Zerbe, R.L.; Robertson, G.L. Osmoregulation of thirst and vasopressin secretion in human subjects: Effect of various solutes. Am. J. Physiol. 1983, 244, E607–E614. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.J.; Bland, J.; Burd, J.; Baylis, P.H. The osmotic thresholds for thirst and vasopressin release are similar in healthy man. Clin. Sci. 1986, 71, 651–656. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.J.; Davis, S.N.; Butler, P.C.; Charlton, J.A.; Baylis, P.H. Osmoregulation of thirst and vasopressin secretion in insulin-dependent diabetes mellitus. Clin. Sci. 1988, 74, 599–606. [Google Scholar] [CrossRef] [PubMed]
- Thompson, C.J.; Edwards, C.R.W.; Baylis, P.H. Osmotic and non-osmotic regulation of thirst and vasopressin secretion in patients with compulsive water drinking. Clin. Endocrinol. 1991, 35, 221–228. [Google Scholar] [CrossRef]
- Zerbe, R.L.; Miller, J.Z.; Robertson, G.L. The reproducibility and heritability of individual differences in osmoregulatory function in normal human subjects. J. Lab. Clin. Med. 1991, 117, 51–59. [Google Scholar]
- Vokes, T. Water homeostasis. Annu. Rev. Nutr. 1987, 7, 383–406. [Google Scholar] [CrossRef]
- Halter, J.B.; Goldberg, A.P.; Robertson, G.L.; Porte, D., Jr. Selective osmoreceptor dysfunction in the syndrome of chronic hypernatremia. J. Clin. Endocrinol. MeTable 1977, 44, 609–616. [Google Scholar] [CrossRef] [PubMed]
- Hochberg, Z.; Moses, A.M.; Miller, M.; Benderli, A.; Richman, R.A. Altered osmotic threshold for vasopressin release and impaired thirst sensation: Additional abnormalities in Kallmann’s syndrome. J. Clin. Endocrinol. Metab. 1982, 55, 779–782. [Google Scholar] [CrossRef] [PubMed]
- Katz, F.H.; Smith, J.A.; Lock, J.P.; Loeffel, D.E. Plasma vasopressin variation and renin activity in normal active humans. Horm. Res. 1979, 10, 289–302. [Google Scholar] [CrossRef] [PubMed]
- Melander, O. Vasopressin: Novel roles for a new hormone-Emerging therapies in cardiometabolic and renal diseases. J. Intern. Med. 2017, 282, 281–283. [Google Scholar] [CrossRef] [PubMed]
- Moses, A.M.; Miller, M.; Streeten, D.H. Quantitative influence of blood volume expansion on the osmotic threshold for vasopressin release. J. Clin. Endocrinol. Metab. 1967, 27, 655–662. [Google Scholar] [CrossRef] [PubMed]
- Robertson, G.L.; Athar, S. The interaction of blood osmolality and blood volume in regulating plasma vasopressin in man. J. Clin. Endocrinol. Metab. 1976, 42, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Schrier, R.W.; Berl, T.; Anderson, R.J. Osmotic and nonosmotic control of vasopressin release. Am. J. Physiol. 1979, 236, F321–F332. [Google Scholar] [CrossRef] [PubMed]
- Zerbe, R.L.; Robertson, G.L. Osmotic and nonosmotic regulation of thirst and vasopressin secretion. In Clinical Disorders of Fluid and Electrolyte Metabolism, 5th ed.; Narins, R., Ed.; McGraw Hill: New York, NY, USA, 1994; Volume 5, pp. 81–100. [Google Scholar]
- Spruce, B.A.; Baylis, P.H.; Burd, J.; Watson, M.J. Variation in osmoregulation of arginine vasopressin during the human menstrual cycle. Clin. Endocrinol. 1985, 22, 37–42. [Google Scholar] [CrossRef]
- Phillips, P.A.; Rolls, B.J.; Ledingham, J.G.; Forsling, M.L.; Morton, J.J.; Crowe, M.J.; Wollner, L. Reduced thirst after water deprivation in healthy elderly men. N. Engl. J. Med. 1984, 311, 753–759. [Google Scholar] [CrossRef]
- Kenney, W.L.; Chiu, P. Influence of age on thirst and fluid intake. Med. Sci. Sports Exerc. 2001, 33, 1524–1532. [Google Scholar] [CrossRef] [Green Version]
- Davies, I.; O’Neill, P.A.; McLean, K.A.; Catania, J.; Bennett, D. Age-associated alterations in thirst and arginine vasopressin in response to a water or sodium load. Age Ageing 1995, 24, 151–159. [Google Scholar] [CrossRef]
- Stachenfeld, N.S.; DiPietro, L.; Nadel, E.R.; Mack, G.W. Mechanism of attenuated thirst in aging: Role of central volume receptors. Am. J. Physiol. 1997, 272, R148–R157. [Google Scholar] [CrossRef] [PubMed]
- Farrell, M.J.; Zamarripa, F.; Shade, R.; Phillips, P.A.; McKinley, M.; Fox, P.T.; Blair-West, J.; Denton, D.A.; Egan, G.F. Effect of aging on regional cerebral blood flow responses associated with osmotic thirst and its satiation by water drinking: A PET study. Proc. Natl. Acad. Sci. U.S.A. 2008, 8, 382–387. [Google Scholar] [CrossRef]
- Morgenthaler, N.G.; Struck, J.; Jochberger, S.; Dünser, M.W. Copeptin: Clinical use of a new biomarker. Trends Endocrinol. Metab. 2008, 19, 43–49. [Google Scholar] [CrossRef]
- Katan, M.; Christ-Crain, M. The stress hormone copeptin: A new prognostic biomarker in acute illness. Swiss Med. Wkly. 2010, 140, w13101. [Google Scholar] [CrossRef] [PubMed]
- Munro, A.H.G.; Crompton, G.K. Inappropriate antidiuretic hormone secretion in oat-cell carcinoma of bronchus Aggravation of hyponatraemia by intravenous cyclophosphamide. Thorax 1972, 27, 40–642. [Google Scholar] [CrossRef]
- Thornton, S.N. Angiotensin, the hypovolaemia hormone, aggravates hypertension, obesity, diabetes and cancer. J. Intern. Med. 2009, 265, 616–617. [Google Scholar] [CrossRef] [Green Version]
- Eric, L. Bittman Vasopressin: More than just an output of the circadian pacemaker? Focus on “Vasopressin receptor V1a regulates circadian rhythms of locomotor activity and expression of clock-controlled genes in the suprachiasmatic nuclei”. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, R821–R823. [Google Scholar]
- Morgenthaler, N.G.; Struck, J.; Alonso, C.; Bergmann, A. Assay for the measurement of copeptin, a stable peptide derived from the precursor of vasopressin. Clin. Chem. 2006, 52, 112–119. [Google Scholar] [CrossRef]
- Roussel, R.; Fezeu, L.; Marre, M.; Velho, G.; Fumeron, F.; Jungers, P.; Lantieri, O.; Balkau, B.; Bouby, N.; Bankir, L.; et al. Comparison between copeptin and vasopressin in a population from the community and in people with chronic kidney disease. J. Clin. Endocrinol. Metab. 2014, 99, 4656–4663. [Google Scholar] [CrossRef]
- Morgenthaler, N.G. Copeptin: A biomarker of cardiovascular and renal function. Congest. Heart Fail. 2010, 16, S37–S44. [Google Scholar] [CrossRef] [PubMed]
- Balanescu, S.; Kopp, P.; Gaskill, M.B.; Morgenthaler, N.G.; Schindler, C.; Rutishauser, J. Correlation of plasma copeptin and vasopressin concentrations in hypo-, iso-, and hyperosmolar States. J. Clin. Endocrinol. Metab. 2011, 96, 1046–1052. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westermann, I.; Dünser, M.W.; Haas, T.; Jochberger, S.; Luckner, G.; Mayr, V.D.; Wenzel, V.; Stadlbauer, K.H.; Innerhofer, P.; Morgenthaler, N.; et al. Endogenous vasopressin and copeptin response in multiple trauma patients. Shock 2007, 28, 644–649. [Google Scholar] [CrossRef] [PubMed]
- Ettema, E.M.; Heida, J.; Casteleijn, N.F.; Boesten, L.; Westerhuis, R.; Gaillard, C.; Gansevoort, R.T.; Franssen, C.F.M.; Zittema, D. The effect of renal function and hemodialysis treatment on plasma vasopressin and copeptin levels. Kidney Int. Rep. 2017, 2, 410–419. [Google Scholar] [CrossRef] [PubMed]
- Clark, W.F.; Sontrop, J.M.; Huang, S.H.; Gallo, K.; Moist, L.; House, A.A.; Cuerden, M.S.; Weir, M.A.; Bagga, A.; Brimble, S.; et al. Effect of coaching to increase water intake on kidney function decline in adults with chronic kidney disease: The CKD WIT randomized clinical trial. JAMA 2018, 319, 1870–1879. [Google Scholar] [CrossRef] [PubMed]
- Enhörning, S.; Tasevska, I.; Roussel, R.; Bouby, N.; Persson, M.; Burri, P.; Bankir, L.; Melander, O. Effects of hydration on plasma copeptin, glycemia and gluco-regulatory hormones: A water intervention in humans. Eur J. Nutr. 2017. [Google Scholar] [CrossRef]
- Balling, L.; Gustafsson, F. Copeptin as a biomarker in heart failure. Biomark Med. 2014, 8, 841–854. [Google Scholar] [CrossRef]
- Szinnai, G.; Morgenthaler, N.G.; Berneis, K.; Struck, J.; Muller, B.; Keller, U.; Christ-Crain, M. Changes in plasma copeptin, the c-terminal portion of arginine vasopressin during water deprivation and excess in healthy subjects. J. Clin. Endocrinol. Metab. 2007, 92, 3973–3978. [Google Scholar] [CrossRef]
- Valtin, H. “Drink at least eight glasses of water a day.” Really? Is there scientific evidence for “8 × 8”? Am. J. Physiol. Regul. Integr. Comp. Physiol. 2002, 283, R993–R1004. [Google Scholar] [CrossRef]
- Gibson, S.; Shirreffs, S.M. Beverage consumption habits “24/7” among British adults: Association with total water intake and energy intake. Nutr. J. 2013, 12, 9. [Google Scholar] [CrossRef]
- Nissensohn, M.; Sánchez-Villegas, A.; Ortega, R.M.; Aranceta-Bartrina, J.; Gil, A.; Gonzalez-Gross, M.; Varela-Moreiras, G.; Serra-Majem, L. Beverage consumption habits and association with total water and energy intakes in the Spanish Population: Findings of the ANIBES study. Nutrients 2016, 8, 232. [Google Scholar] [CrossRef]
- Chang, A.; Kramer, H. Fluid intake for kidney disease prevention: An urban myth? Clin. J. Am. Soc. Nephrol. 2011, 6, 2558–2560. [Google Scholar] [CrossRef] [PubMed]
- Bouby, N.; Clark, W.F.; Roussel, R.; Taveau, C.; Wang, C.J. Hydration and kidney health. Obes. Facts. 2014, 7, 19–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bankir, L.; Bouby, N.; Ritz, E. Vasopressin: A novel target for the prevention and retardation of kidney disease? Nat. Rev. Nephrol. 2013, 9, 223–239. [Google Scholar] [CrossRef]
- Staub, D.; Morgenthaler, N.G.; Buser, C.; Breidthardt, T.; Potocki, M.; Noveanu, M.; Reichlin, T.; Bergmann, A.; Mueller, C. Use of copeptin in the detection of myocardial ischemia. Clin. Chim. Acta. 2009, 399, 69–73. [Google Scholar] [CrossRef]
- Schrier, R.W.; Masoumi, A.; Elhassan, E. Role of vasopressin and vasopressin receptor antagonists in type I cardiorenal syndrome. Blood Purif. 2009, 27, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Decaux, G.; Soupart, A.; Vassart, G. Non-peptide arginine-vasopressin antagonists: The vaptans. Lancet 2008, 371, 1624–1632. [Google Scholar] [CrossRef]
- Melander, O. Vasopressin, from regulator to disease predictor for diabetes and cardiometabolic risk. Ann. Nutr. Metab. 2016, 68, 24–28. [Google Scholar] [CrossRef]
- Armstrong, L.E. Challenges of linking chronic dehydration and fluid consumption to health outcomes. Nutr. Rev. 2012, 70, S121–S127. [Google Scholar] [CrossRef]
- Enhörning, S.; Bankir, L.; Bouby, N.; Struck, J.; Hedblad, B.; Persson, M.; Morgenthaler, N.G.; Nilsson, P.M.; Melander, O. Copeptin, a marker of vasopressin, in abdominal obesity, diabetes and microalbuminuria: The prospective Malmo Diet and Cancer Study cardiovascular cohort. Int. J. Obes. 2013, 37, 598–603. [Google Scholar]
- Enhörning, S.; Hedblad, B.; Nilsson, P.M.; Engström, G.; Melander, O. Copeptin is an independent predictor of diabetic heart disease and death. Am. Heart J. 2015, 169, 549–556. [Google Scholar] [CrossRef]
- Tasevska, I.; Enhörning, S.; Christensson, A.; Persson, M.; Nilsson, P.M.; Melander, O. Increased levels of copeptin, a surrogate marker of arginine vasopressin, are associated with an increased risk of chronic kidney disease in a general population. Am. J. Nephrol. 2016, 44, 22–28. [Google Scholar] [CrossRef]
- Velho, G.; El Boustany, R.; Lefèvre, G.; Mohammedi, K.; Fumeron, F.; Potier, L.; Bankir, L.; Bouby, N.; Hadjadj, S.; Marre, M.; et al. Plasma Copeptin, Kidney Outcomes, Ischemic Heart Disease, and All-Cause Mortality in People With Long-standing Type 1 Diabetes. Diabetes Care 2016, 39, 2288–2295. [Google Scholar] [CrossRef]
- Borghi, L.; Meschi, T.; Amato, F.; Briganti, A.; Novarini, A.; Giannini, A. Urinary volume, water and recurrences in idiopathic calcium nephrolithiasis: A 5-year randomized prospective study. J. Urol. 1996, 155, 839–843. [Google Scholar] [CrossRef]
- Borghi, L.; Meschi, T.; Schianchi, T.; Allegri, F.; Guerra, A.; Maggiore, U.; Novarini, A. Medical treatment of nephrolithiasis. Endocrinol. Metab. Clin. North Am. 2002, 31, 1051–1064. [Google Scholar] [CrossRef]
- Borghi, L.; Meschi, T.; Schianchi, T.; Briganti, A.; Guerra, A.; Allegri, F.; Novarini, A. Urine volume: Stone risk factor and preventive measure. Nephron 1999, 81, 31–37. [Google Scholar] [CrossRef]
- Siener, R.; Hesse, A. Fluid intake and epidemiology of urolithiasis. Eur. J. Clin. Nutr. 2003, 57, S47–S51. [Google Scholar] [CrossRef]
- Siener, R.; Schade, N.; Nicolay, C.; von Unruh, G.E.; Hesse, A. The efficacy of dietary intervention on urinary risk factors for stone formation in recurrent calcium oxalate stone patients. J. Urol. 2005, 173, 1601–1605. [Google Scholar] [CrossRef]
- Wenzel, U.O.; Hebert, L.A.; Stahl, R.A.; Krenz, I. My doctor said I should drink a lot! Recommendations for fluid intake in patients with chronic kidney disease. Clin. J. Am. Soc. Nephrol. 2006, 1, 344–346. [Google Scholar] [CrossRef]
- Wang, C.J.; Grantham, J.J.; Wetmore, J.B. The medicinal use of water in renal disease. Kidney Int. 2013, 84, 45–53. [Google Scholar] [CrossRef]
- Clark, W.F.; Sontrop, J.M.; Huang, S.H.; Moist, L.; Bouby, N.; Bankir, L. Hydration and Chronic Kidney Disease Progression: A Critical Review of the Evidence. Am. J. Nephrol. 2016, 43, 281–292. [Google Scholar] [CrossRef]
- Sontrop, J.M.; Dixon, S.N.; Garg, A.X.; Buendia-Jimenez, I.; Dohein, O.; Huang, S.H.; Clark, W.F. Association between water intake, chronic kidney disease, and cardiovascular disease: A cross-sectional analysis of NHANES data. Am. J. Nephrol. 2013, 37, 434–442. [Google Scholar] [CrossRef]
- Farini, F. Diabete insipido ed opoterapia. Gazz. Osped. Clin. 1913, 34, 1135–1139. [Google Scholar]
- Ragnarsson, U. The nobel trail of Vincent du Vigneaud. J. Pept. Sci. 2007, 13, 431–433. [Google Scholar] [CrossRef]
- Zerbe, R.L.; Vinicor, F.; Robertson, G.L. Plasma vasopressin in uncontrolled diabetes mellitus. Diabetes 1979, 28, 503–508. [Google Scholar] [CrossRef]
- Enhörning, S.; Wang, T.J.; Nilsson, P.M.; Almgren, P.; Hedblad, B.; Berglund, G.; Struck, J.; Morgenthaler, N.G.; Bergmann, A.; Lindholm, E.; et al. Plasma copeptin and the risk of diabetes mellitus. Circulation 2010, 121, 2102–2108. [Google Scholar]
- Saleem, U.; Khaleghi, M.; Morgenthaler, N.G.; Bergmann, A.; Struck, J.; Mosley, T.H., Jr.; Kullo, I.J. Plasma carboxy-terminal provasopressin (copeptin): A novel marker of insulin resistance and metabolic syndrome. J. Clin. Endocrinol. Metab. 2009, 94, 2558–2564. [Google Scholar] [CrossRef]
- Roussel, R.; Fezeu, L.; Bouby, N.; Balkau, B.; Lantieri, O.; Alhenc-Gelas, F.; Marre, M.; Bankir, L. Low water intake and risk for new-onset hyperglycemia. Diabetes Care 2011, 34, 2551–2554. [Google Scholar] [CrossRef]
- Nielsen, S.; Chou, C.L.; Marples, D.; Christensen, E.I.; Kishore, B.K.; Knepper, M.A. Vasopressin increases water permeability of kidney collecting duct by inducing translocation of aquaporin-CD water channels to plasma membrane. Proc. Natl. Acad. Sci. USA 1995, 92, 1013–1017. [Google Scholar] [CrossRef]
- Nakamura, K.; Velho, G.; Bouby, N. Vasopressin and metabolic disorders: Translation from experimental models to clinical use. J. Intern. Med. 2017, 282, 298–309. [Google Scholar] [CrossRef]
- Taveau, C.; Chollet, C.; Bichet, D.G.; Velho, G.; Guillon, G.; Corbani, M.; Roussel, R.; Bankir, L.; Melander, O.; Bouby, N. Acute and chronic hyperglycemic effects of vasopressin in normal rats: Involvement of V1A receptors. Am. J. Physiol. Endocrinol. Metab. 2017, 312, E127–E135. [Google Scholar] [CrossRef] [PubMed]
- Taveau, C.; Chollet, C.; Waeckel, L.; Desposito, D.; Bichet, D.G.; Arthus, M.F.; Magnan, C.; Philippe, E.; Paradis, V.; Foufelle, F.; et al. Vasopressin and hydration play a major role in the development of glucose intolerance and hepatic steatosis in obese rats. Diabetologia 2015, 58, 1081–1090. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Enhörning, S.; Leosdottir, M.; Wallstrom, P.; Gullberg, B.; Berglund, G.; Wirfalt, E.; Melander, O. Relation between human vasopressin 1a gene variance, fat intake, and diabetes. Am. J. Clin. Nutr. 2009, 89, 400–406. [Google Scholar] [CrossRef] [PubMed]
- Kuo, T.; McQueen, A.; Chen, T.C.; Wang, J.C. Regulation of glucose homeostasis by glucocorticoids. Adv. Exp. Med. Biol. 2015, 872, 99–126. [Google Scholar]
- Johnson, E.C.; Bardis, C.N.; Jansen, L.T.; Adams, J.D.; Kirkland, T.W.; Kavouras, S.A. Reduced water intake deteriorates glucose regulation in patients with type 2 diabetes. Nutr. Res. 2017, 43, 25–32. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Life Stage & Sex | Age | European Food Safety Authority, Parma, Italy b 2010 (ml/day) | National Academy of Medicine, USA 2004 b (ml/day) |
|---|---|---|---|
| Infants | 0–6 months | 680 via milk | 700 |
| 6–12 months | 800–1,000 | 800 | |
| Children | 1–2 years | 1100-1200 | 1300 |
| 2–3 years | 1300 | ||
| 4–8 years | 1600 | 1700 | |
| 9–13 years, boys | 2100 | 2400 | |
| 9–13 years, girls | 1900 | 2100 | |
| 14–18 years, boys | 2500 | 3300 | |
| 14–18 years, girls | 2000 | 2300 | |
| Adults | |||
| Men | 2500 | 3700 | |
| Women | 2000 | 2700 | |
| Pregnant Women | ≥ 19 year | 2300 | 3000 |
| Lactating Women | ≥ 19 year | 2600–2700 | 3800 |
| Elderly | same as adults | same as adults |
| Methods | Variables Measured or Calculated | Relevance | Critique | Representative Publications | |
|---|---|---|---|---|---|
| Individuals | Group | ||||
| Partitioning 24-h urine production into minimum urine volume a and free urine volume b | UVOL, UMAX, UVM, UFUV | X | UMAX is determined via observations of a few males and was applied to individuals. UMAX varies with age and had a large inter-subject variability. | [9] | |
| Calculation of free water reserve c to determine individual 24-hour hydration status d | UVOL, UMAX, UVM, UFWR, UOSM, UTOT, NRWL | X | This population-based method updates the concepts of Gamble (above), does not determine the UMAX of individuals, and estimates NRWL. In single (<24h) samples, confounding factors e may dominate and other hydration biomarkers are preferred. | [10,11,12] | |
| Dietary recall to determine TWI | plain water, beverages, food moisture | X | X | Data are specific to the subject sample, and typically do not provide information regarding water balance or turnover. | [13,14,15] |
| Responses and hydration biomarkers of free-living LD f versus HD f | UOSM, USG, UCOL, UVOL, POSM, SOSM, MB | X | Studies assess the responses of adult groups who have habitually different TWIs. | [13,16,17,18,19,20] | |
| Global, regional, and country water consumption recommendations | TWI (L/24 h) | X | Adequate intakes g for TWI are based on survey data median values. | [6,7,21] | |
| Statistical categories of hydration status for free-living adults | UOSM,USG, UCOL, UVOL, POSM, MB | X | Seven categories range from euhydrated to hypohydrated or hyperhydrated. Variables are expressed per single sample and 24-h collection. | [22,23] | |
| Laboratory water turnover and movement, using the DLW technique or stable isotope of water h | 2H218O, 2H2O, TBW, UVOL, NRWL | X | Mean water turnover (L/24h) incorporates estimates of TWI, metabolic, transcutaneous, and inspired air water. | [22,24,25] | |
| Water balance of free-living adults during daily activities | TBW, TWI, UOSM, UVOL, POSM, TPP, HCT, SR, MB | X | X | Various methods are used to describe the water needs of specific life stage and sex groups. | [2,25,26] |
| Laboratory controlled experiments evaluating dehydration and rehydration | UOSM,USG, UCOL, UVOL, POSM, SOSM, MB,%∆PV | X | Dehydration is accomplished via passive exposure to a hot environment, exercise, or water restriction. Rehydration is accomplished via water and beverage intake or intravenous fluid administration. | [27,28,29,30,31,32] | |
| Laboratory investigations that focus on thirst sensations and drinking behavior | TWI, beverages, UOSM, UVOL, POSM,%∆PV, AVP | X | [33,34,35] | ||
| Laboratory comparison of beverages: rehydration efficacy | FC, UVOL, BHI | X | Common beverages are evaluated to identify retention (relative to still water) in euhydrated, but not dehydrated, adults. The diuretic response is influenced by fluid characteristics including osmolality, energy density, and electrolyte content. | [36,37] | |
| Plasma AVP or copeptin i responses | AVP, copeptin i | X | X | The hormone AVP maintains UVOL, POSM, and body water balance within narrow limits, in conjunction with thirst. | [13,16,19,31] |
| Assessment of specific urine and plasma hydration biomarkers | UOSM, UCOL, POSM, POSM:UOSM ratio | X | X | Most studies focus on the assessment of simple, practical hydration biomarkers for use during daily activities. | [16,18,38,39,40,41] |
| Field studies of hydration status during labor, exercise, or competition | TWI, SR, POSM, UOSM, USG, UCOL, MB | X | Research attempts to optimize health and performance. | [42,43,44,45] | |
| Statistical and graphical determination of the probability of dehydration | POSM, USG, MB | X | X | Predictions are based on a modest dehydration range (−2.1 to −3.5% Mb) in 6 men and 5 women. | [46] |
| Calculated biological variation and diagnostic accuracy of dehydration biomarkers | POSM, SOSM, UOSM, USG, UCOL, MB | X | X | Statistics evaluate biomarkers, on the basis of a functionally important range of −2.0 to −7.0% Mb, induced in 5 women and 13 men across x hours. | [47] |
| Theoretical consideration of intracellular and extracellular dehydration | PNa+, POSM, SOSM, UOSM, USG, UCOL, MB | X | Candidate biomarkers of dehydration must consider intracellular, extracellular, and mixed dehydration stimuli. | [48] | |
| Total water intake a (L/24h) | Intracellular metabolic water production b | Total solute load c (mOsm/24h) | Urine osmolality d (mOsm/kg) | Maximal renal concentrating ability (mOsm/kg) | Urine volume (L/24h) | Non-renal water loss (L/24h) e | Free water reserve f (L/24h) | |
|---|---|---|---|---|---|---|---|---|
| Functions and characteristics | Contributes to TBW | Product of human metabolism | Metabolized and digested products excreted in urine | Regulates TBW and ECV-ICV osmolality | Inherent quality of the kidneys | Regulates TBW and ECV-ICV osmolality | Excretory and secretory processes | Calculated index of euhydration, based on population statistics |
| Influential factors | Meal timing and contents, idiosyncratic thirst, physical activity, body size, cultural and learned preferences | Metabolic rate and substrates, physical activity, diet macronutrient and energy content, NES responses | Metabolic products, dietary contents, body size, idiosyncratic hunger, learned food preferences | TWI, MRCA, solute load, NRWL, physical activity, NES responses | Life-stage group, male or female sex | TWI, total solute load, NRWL, physical activity, NES responses | Diet, ventilatory rate, physical activity, body size | TWI, total solute load, NRWL, physical activity |
| Organs involved | GI, CNS, NES, mouth and throat | CNS, NES | GI, CNS, NES | Kidneys, CNS, NES | Kidneys, CNS, NES | GI, kidneys, CNS, NES | Skin, GI, respiratory organs | GI, kidneys, CNS, NES |
| Conscious or behavioral influence? | Yes, habitual 24-h water intake | No | Yes, solid food consumption | Yes, secondary to TWI and food contents | No | Yes, secondary to TWI and food contents | Yes, eccrine sweat loss during labor or exercise | Yes, secondary to water and food intake |
| Representative mean, median, or range of values for sedentary adults | ♀, 1.8–2.0 and ♂, 1.9–2.4 (FR, UK); ♀&♂, 1.5–2.5 (13countries); ♀, 2.3 (range: 0.8–4.5) (USA); ♂, 3.0 (range: 1.4–7.7) and ♀, 2.5 (range: 1.2–4.6) (USA); ♀, 1.9 and ♂, 2.3 (GE); ♀&♂, 0.2–3.9 (FR) L/24h | ♀, 0.2–0.3; ♂, 0.3–0.4; ♂, 0.4 L/24h | ♀, 669–781 and ♂, 915–992 (GE); ♂, 951 (USA); ♀&♂, 362–1365 (4 countries); ♂, 750 (USA); ♀, 752 and ♂, 941 (GE) mOsm/24h | ♀&♂, 120–1250 (FR); ♀&♂, 555 (UK) mOsm/kg | ♀&♂, 1430 (UK) mOsm/kg; ♀&♂ range, 1100–1300 (GE); ♀&♂, 1010–1330 (USA) | ♀&♂, 0.2–3.9 (FR); ♀&♂, 1.9 (n = 8, UK) L/24h | ♂, 0.3–0.4 (UK); ♀, 0.5–0.7 and ♂, 0.7–1.3 (GE) L/24h | ♀, 0.4–0.5 and ♂, 0.2–0.3 (GE); ♀, 0.5 and ♂, 0.3 (GE) L/24h |
| Reference citations | [11,14,15,25,38,51] | [25,52,53] | [1,11,12,41] | [38,54] | [1,54,55] | [38,54] | [1,24] | [11] |
| Osmotic Threshold b (mOsm/kg) | Participants/Conditions | References |
|---|---|---|
| 282 (280–285) | Normal adults (n = 6♂), dehydration via water restriction, upright posture | Moses and Miller, 1971 [70] |
| 285 (284–286) c | Normal adults (n = 9♂), IVHS | Moses and Miller, 1971 [70] |
| 287 (286–288) c | Normal adults (n = 6♂), IVHS | Moses and Miller, 1971 [70] |
| 288 (287–289) c | Normal adults (n = 6♂), IVHS, then IVHS plus dextran (expanded plasma volume) | Moses and Miller, 1971 [70] |
| 280 (272–284) c | Normal adults (n = 25), recumbent rest, in three states: ad libitum fluid intake, acute water load (20 ml/kg) and water restriction | Robertson et al., 1973 [71] |
| 280 (276–291) c | Normal adults (n = 9♂, 7♀), recumbent rest | Robertson et al., 1976 [72] |
| IVHS, 287 (283–291) M, 286 (282–290) | Normal adults (3♀, 3♂), supine rest, IVHS (5%) and hypertonic mannitol (M, 20%) | Zerbe et al., 1983 [73] |
| 285 (282–289) | Healthy adults (10♂), recumbent rest, IVHS and IVI | Thompson et al., 1986 [74] |
| 287 (286–288) | Healthy adults (7♂), recumbent rest, IVHS | Thompson et al., 1988 [75] |
| 287 (281–290) | Healthy adults (3♂, 4♀), recumbent rest, IVHS | Thompson et al., 1991 [76] |
| MZ, 283 (277–290) DZ, 281 (274–285) | Healthy twins (7♂ monozygotic pairs, 6♂ dizygotic pairs), IVHS | Zerbe et al., 1991 [77] |
| Biological functions |
| Regulates body water and sodium homeostasis by acting on renal nephrons to decrease urine volume and increase the concentration of urine Maintains plasma osmolality within narrow limits Encourages vasodilation in vascular smooth muscle cells by inducing NO production Affects liver metabolism (e.g., gluconeogenesis, glycogenolysis) Stimulates the pancreas to produce either insulin or glucagon, depending on blood glucose concentration |
| Factors which influence neurohypohysial AVP release and plasma AVP concentration |
| Plasma osmolality Angiotensin II Oropharyngeal environment Water restriction and consumption Osmolar content of diet, especially sodium Hypoglycemia Blood volume and pressure Upright posture Emotional stress Exercise Circadian rhythmicity Hypoxia Nausea Pain |
| Diseases and disorders that involve AVP dysfunction |
| Diabetes insipidus and diabetes mellitus Syndrome of inappropriate ADH excess (SIADH) Sepsis Severe bleeding, hemorrhage Chronic hypernatremia Primary polydipsia syndrome, compulsive water drinking Kallmann’s syndrome Autosomal dominant polycystic kidney disease |
| Mean (Range b) Osmotic Threshold (mOsm/kg) | Participants/Conditions | References |
|---|---|---|
| IVHS, 298 (294–300) M, 296 (290–299) | Normal adults (n = 2–5 ♂&♀), supine rest, IVHS (5%) and hypertonic mannitol (M, 20%) | Zerbe et al., 1983 [73] |
| F, 297 (296–298) L, 293 (291–295) | Healthy women (n = 8) were tested in the follicular (F) and luteal (L) phases of the menstrual cycle, IVHS | Spruce et al., 1985 [87] |
| 287 (286–288) | Healthy males (n = 7), recumbent rest, IVHS | Thompson et al., 1988 [75] |
| 287 (282–291) | Healthy adults (3♂, 4♀), recumbent rest, IVHS | Thompson et al., 1991 [76] |
| MZ, 286 (276–293) DZ, 289 (283–296) | Healthy twins (7♂ monozygotic pairs, 6♂ dizygotic pairs), IVHS | Zerbe et al., 1991 [77] |
| Participants | Experimental Design Phase | Plasma Osmolality (mOsm/kg H2O) | Plasma AVP (pg/ml) | Reference |
|---|---|---|---|---|
| 8 ♀ b (21–34 year) | Baseline, EU | 289 ± 2 | 1.3 ± 0.6 | Davison et al., AJP 1984 [62] |
| 12-h WR c | 294 ± 2 | 2.9 ± 1.2 | ||
| 5 ♂ & 3 ♀ b (26–50 year) | Baseline, EU | 292 ± 1 | 1.7 ± 0.2 | Geelen et al., AJP 1984 [63] |
| 24-h WR c | 302 ± 1 | 3.3 ± 0.5 | ||
| 7 ♂ (20–31 year) | Baseline, EU | 288 ± 1 | 1.0 ± 0.3 | Phillips et al., NEJM 1984 [88] |
| 24-h WR c,d | 291 ± 1 | 3.5 ± 0.3 | ||
| 7 ♂ (67–75 year) | Baseline, EU | 288 ± 1 | 1.8 ± 0.3 | Phillips et al., NEJM 1984 [88] |
| 24-h WR c,d | 296 ± 1 | 8.3 ± 0.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Armstrong, L.E.; Johnson, E.C. Water Intake, Water Balance, and the Elusive Daily Water Requirement. Nutrients 2018, 10, 1928. https://doi.org/10.3390/nu10121928
Armstrong LE, Johnson EC. Water Intake, Water Balance, and the Elusive Daily Water Requirement. Nutrients. 2018; 10(12):1928. https://doi.org/10.3390/nu10121928
Chicago/Turabian StyleArmstrong, Lawrence E., and Evan C. Johnson. 2018. "Water Intake, Water Balance, and the Elusive Daily Water Requirement" Nutrients 10, no. 12: 1928. https://doi.org/10.3390/nu10121928