Human Breast Milk Bacteriome in Health and Disease


Abstract
1. Introduction
2. Human Breast Milk Bacteriome
3. Bacterial Diversity in Human Breast Milk
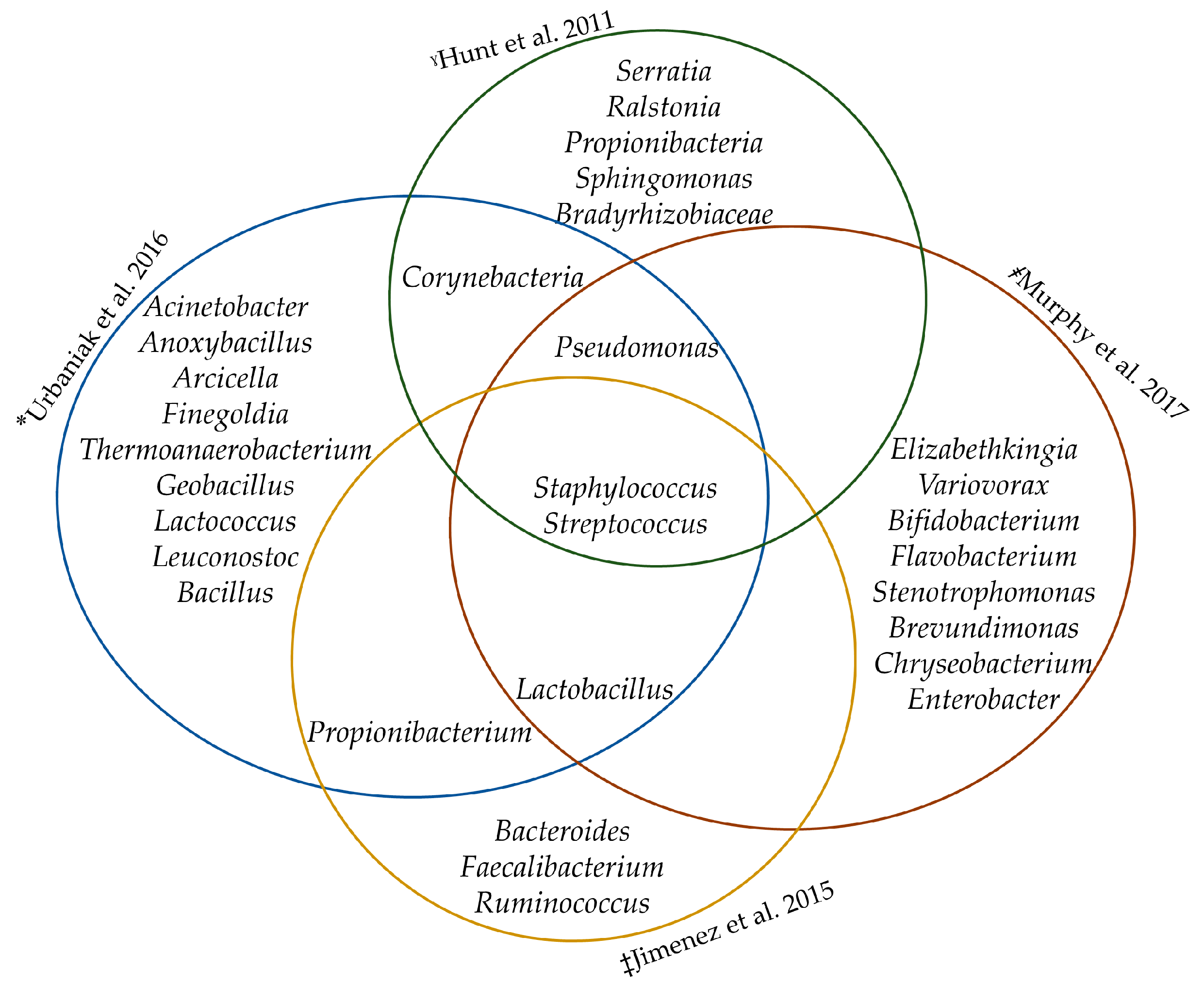
3.1. The Core Milk Bacteriome?
3.2. Origin of The Human Breast Milk Bacteriome
3.2.1. Retrograde Flow
3.2.2. Gut–Mammary-Axis
3.2.3. Mammary Gland Bacteriome
4. Factors Which May Affect the Human Breast Milk Bacteriome
- Mode of delivery: An estimate by qPCR [34,35] claims that women who delivered via caesarean section (CS) have been shown to have a lower abundance of Lactobacillus spp. (L. fermentum and L. salivarius), Bifidobacterium spp. when compared with the higher bacterial counts of women who delivered vaginally. The HBM of mothers who had elective CS also showed decreased members of the family Leuconostocaceae and increased Carnobacteriaceae, when compared with women who delivered vaginally [18]. However, in a study by Urbaniak and colleagues which utilized a more robust statistical analysis [19], no difference in bacterial profiles was observed between women who delivered vaginally and those who delivered via emergency CS. It was suggested that this could be due to the initiation of the labor process, including physiological stress and hormonal signals which may influence increased permeability of intestinal epithelial lining for translocation [18,19].
- Maternal weight: Higher levels of Staphylococcus spp. and lower levels of Bifidobacterium spp. were observed in HBM from overweight mothers as compared with normal-weight mothers [36]; and a less diverse bacterial community has also been observed in obese mothers [18]. This may be due to the metabolic capacity of the bacteriome of obese individuals which has an increased capacity to harvest energy from diet [37].
- Antibiotics and Chemotherapy: A lower abundance of lactobacilli and bifidobacteria was detected in HBM of mothers who were exposed to antibiotics during the perinatal stage [35]. Exposure to anti-cancer chemotherapy also resulted in a reduction of the genera Bifidobacterium, Eubacterium, Staphylococcus and Cloacibacterium [38].
- Maternal health: Gronlund et al. (2007) described that the bacteriome is influenced by maternal health. In his study using direct PCR analysis, allergic women exhibited a significantly lower Bifidobacterium spp. in their BM, with their infants also having lower fecal bifidobacteria counts [39]. African women with HIV–RNA in their HBM had an increased bacterial diversity and higher abundance of Lactobacillus spp. compared to controls [40]. Lower abundance of Bifidobacterium spp. and Bacteroides fragilis group have been detected in HBM of women with celiac disease [41].
- Lactation stage: A higher bacterial diversity but lower total bacterial count and less bifidobacteria species were detected in colostrum when compared with mature HBM [18]. Bifidobacterium spp. and Enterococcus spp. counts, along with total bacteria increased as the lactation stage progressed [34]. In a similar study, however, transition milk samples were observed to possess higher diversity than colostrum and mature milk [34,42].
- Geographical location: The bacterial genera found in HBM of Spanish mothers were different to those of Americans [5,42], or Finnish women [18] using sequencing techniques with a similarly high throughput. In a study by Kumar et al., Chinese women had high levels of Actinobacteria in comparison to the similarly high levels of Bacteroidetes detected in Spanish women [43].
- Gestational age: Bifidobacterium spp. were observed to be higher in HBM of women with term babies than in preterm gestation.
5. Plausible Functions of Human Breast Milk Bacteria
5.1. Vertical Transmission and Seeding of Infant Gut by HBM Bacteria
5.2. Anti-Infective Activities of HBM Bacteria
5.3. Immuno-Modulatory Activities of HBM Bacteria
5.4. Anti-Allergic Properties of HBM Bacteria
5.5. Metabolic Activities of HBM Bacteria
5.6. Anti-Tumor Properties of HBM Bacteria
6. Implications of the Human Breast Milk Bacteriome in Disease
6.1. Mastitis
6.2. Human Immunodeficiency Virus (HIV)
6.3. Cancer
7. Conclusions
Gaps Identified
- There are very limited animal studies demonstrating the specific role of breast milk bacteria in disease conditions.
- There are few integrative studies exploring the interplay between HBM bacteriome and its metabolites, and their collective role in diseased condition.
- There is insufficient data on some of the identified HBM bacterial groups. It isn’t enough to simply characterize the bacterial diversity of HBM; rather, the role that each of the bacterium play in maintaining the microbial ecology of the HBM bacteriome and their role in infant and maternal health must be understood.
- There are few studies on HBM bacteriome and the factors influencing low socio-economic regions, such as many African settings. These regions also boast less research of infant nutrition and this research is necessary to know if there are any underlying genetic mechanisms involved.
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Agostoni, C.; Braegger, C.; Decsi, T.; Kolacek, S.; Koletzko, B.; Michaelsen, K.F.; Mihatsch, W.; Moreno, L.A.; Puntis, J.; Shamir, R.; et al. Breast-feeding: A commentary by the espghan committee on nutrition. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 112–125. [Google Scholar] [CrossRef] [PubMed]
- Civardi, E.; Garofoli, F.; Tzialla, C.; Paolillo, P.; Bollani, L.; Stronati, M. Microorganisms in human milk: Lights and shadows. J. Matern. Fetal Neonatal Med. 2013, 26, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, L.; Langa, S.; Martin, V.; Jimenez, E.; Martin, R.; Rodriguez, J.M. The microbiota of human milk in healthy women. Cell Mol. Biol. 2013, 59, 31–42. [Google Scholar] [PubMed]
- Aakko, J.; Kumar, H.; Rautava, S.; Wise, A.; Autran, C.; Bode, L.; Isolauri, E.; Salminen, S. Human milk oligosaccharide categories define the microbiota composition in human colostrum. Benef. Microbes 2017, 8, 563–567. [Google Scholar] [CrossRef] [PubMed]
- Hunt, K.M.; Foster, J.A.; Forney, L.J.; Schutte, U.M.; Beck, D.L.; Abdo, Z.; Fox, L.K.; Williams, J.E.; McGuire, M.K.; McGuire, M.A. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS ONE 2011, 6, e21313. [Google Scholar] [CrossRef] [PubMed]
- Ward, T.L.; Hosid, S.; Ioshikhes, I.; Altosaar, I. Human milk metagenome: A functional capacity analysis. BMC Microbiol. 2013, 13, 116. [Google Scholar] [CrossRef] [PubMed]
- Boix-Amorós, A.; Martinez-Costa, C.; Querol, A.; Collado, M.C.; Mira, A. Multiple approaches detect the presence of fungi in human breastmilk samples from healthy mothers. Sci. Rep. 2017, 7, 13016. [Google Scholar] [CrossRef] [PubMed]
- Goldman, A.S. Evolution of immune functions of the mammary gland and protection of the infant. Breastfeed. Med. 2012, 7, 132–142. [Google Scholar] [CrossRef] [PubMed]
- Heikkila, M.P.; Saris, P.E. Inhibition of staphylococcus aureus by the commensal bacteria of human milk. J. Appl. Microbiol. 2003, 95, 471–478. [Google Scholar] [CrossRef] [PubMed]
- Asnicar, F.; Manara, S.; Zolfo, M.; Truong, D.T.; Scholz, M.; Armanini, F.; Ferretti, P.; Gorfer, V.; Pedrotti, A.; Tett, A.; et al. Studying vertical microbiome transmission from mothers to infants by strain-level metagenomic profiling. Am. Soc. Microbiol. 2017, 2. [Google Scholar] [CrossRef] [PubMed]
- Jost, T.; Lacroix, C.; Braegger, C.; Chassard, C. Assessment of bacterial diversity in breast milk using culture-dependent and culture-independent approaches. Br. J. Nutr. 2013, 110, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Jost, T.; Lacroix, C.; Braegger, C.P.; Rochat, F.; Chassard, C. Vertical mother-neonate transfer of maternal gut bacteria via breastfeeding. Environ. Microbiol. 2014, 16, 2891–2904. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.; de Andres, J.; Manrique, M.; Pareja-Tobes, P.; Tobes, R.; Martinez-Blanch, J.F.; Codoner, F.M.; Ramon, D.; Fernandez, L.; Rodriguez, J.M. Metagenomic analysis of milk of healthy and mastitis-suffering women. J. Hum. Lact. 2015, 31, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Langa, S.; Reviriego, C.; Jiminez, E.; Marin, M.L.; Xaus, J.; Fernandez, L.; Rodriguez, J.M. Human milk is a source of lactic acid bacteria for the infant gut. J. Pediatr. 2003, 143, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Kenneth, T. Important Groups of Procaryotes, Today’s Online Textbook of Bacteriology. Available online: http://textbookofbacteriology.net/ (accessed on 15 September 2018).
- Mohania, D.; Nagpal, R.; Kumar, M.; Bhardwaj, A.; Yadav, M.; Jain, S.; Marotta, F.; Singh, V.; Parkash, O.; Yadav, H. Molecular approaches for identification and characterization of lactic acid bacteria. J. Dig. Dis. 2008, 9, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Fouhy, F.; Ross, R.P.; Fitzgerald, G.F.; Stanton, C.; Cotter, P.D. Composition of the early intestinal microbiota: Knowledge, knowledge gaps and the use of high-throughput sequencing to address these gaps. Gut Microbes 2012, 3, 203–220. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Rubio, R.; Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E.; Mira, A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am. J. Clin. Nutr. 2012, 96, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, C.; Angelini, M.; Gloor, G.B.; Reid, G. Human milk microbiota profiles in relation to birthing method, gestation and infant gender. Microbiome 2016, 4, 1. [Google Scholar] [CrossRef] [PubMed]
- Fitzstevens, J.L.; Smith, K.C.; Hagadorn, J.I.; Caimano, M.J.; Matson, A.P.; Brownell, E.A. Systematic review of the human milk microbiota. Nutr. Clin. Pract. 2016. [Google Scholar] [CrossRef] [PubMed]
- Murphy, K.; Curley, D.; O’Callaghan, T.F.; O’Shea, C.A.; Dempsey, E.M.; O’Toole, P.W.; Ross, R.P.; Ryan, C.A.; Stanton, C. The composition of human milk and infant faecal microbiota over the first three months of life: A pilot study. Sci. Rep. 2017, 7, 40597. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.; Delgado, S.; Fernandez, L.; Garcia, N.; Albujar, M.; Gomez, A.; Rodriguez, J.M. Assessment of the bacterial diversity of human colostrum and screening of staphylococcal and enterococcal populations for potential virulence factors. Res. Microbiol. 2008, 159, 595–601. [Google Scholar] [CrossRef] [PubMed]
- Shetty, S.A.; Hugenholtz, F.; Lahti, L.; Smidt, H.; de Vos, W.M. Intestinal microbiome landscaping: Insight in community assemblage and implications for microbial modulation strategies. FEMS Microbiol. Rev. 2017, 41, 182–199. [Google Scholar] [CrossRef] [PubMed]
- Salonen, A.; Salojarvi, J.; Lahti, L.; de Vos, W.M. The adult intestinal core microbiota is determined by analysis depth and health status. Clin. Microbiol. Infect. 2012, 18, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Pannaraj, P.S.; Li, F.; Cerini, C.; Bender, J.M.; Yang, S.; Rollie, A.; Adisetiyo, H.; Zabih, S.; Lincez, P.J.; Bittinger, K.; et al. Association between breast milk bacterial communities and establishment and development of the infant gut microbiome. JAMA Pediatr. 2017, 171, 647–654. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, D.T.; Kent, J.C.; Owens, R.A.; Hartmann, P.E. Ultrasound imaging of milk ejection in the breast of lactating women. Pediatrics 2004, 113, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.M. The origin of human milk bacteria: Is there a bacterial entero-mammary pathway during late pregnancy and lactation? Adv. Nutr. 2014, 5, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Perez, P.F.; Dore, J.; Leclerc, M.; Levenez, F.; Benyacoub, J.; Serrant, P.; Segura-Roggero, I.; Schiffrin, E.J.; Donnet-Hughes, A. Bacterial imprinting of the neonatal immune system: Lessons from maternal cells? Pediatrics 2007, 119, e724–e732. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.; Voigt, A.; Paveglio, S.; Weinstock, G.; Matson, A.; Davidovics, Z. Similar bacterial signatures in intestinal tissues, milk, and dendritic cells of lactating mice suggest a possible entero-mammary pathway. Gastroenterology 2017, 152, S172. [Google Scholar] [CrossRef]
- Urbaniak, C.; Cummins, J.; Brackstone, M.; Macklaim, J.M.; Gloor, G.B.; Baban, C.K.; Scott, L.; O’Hanlon, D.M.; Burton, J.P.; Francis, K.P.; et al. Microbiota of human breast tissue. Appl. Environ. Microbiol. 2014, 80, 3007–3014. [Google Scholar] [CrossRef] [PubMed]
- Hieken, T.J.; Chen, J.; Hoskin, T.L.; Walther-Antonio, M.; Johnson, S.; Ramaker, S.; Xiao, J.; Radisky, D.C.; Knutson, K.L.; Kalari, K.R.; et al. The microbiome of aseptically collected human breast tissue in benign and malignant disease. Sci. Rep. 2016, 6, 30751. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.A.; Bashir, M.; Rivas, M.N.; Duvall, K.; Sieling, P.A.; Pieber, T.R.; Vaishampayan, P.A.; Love, S.M.; Lee, D.J. Characterization of the microbiome of nipple aspirate fluid of breast cancer survivors. Sci. Rep. 2016, 6, 28061. [Google Scholar] [CrossRef] [PubMed]
- Petrakis, N.L. Nipple aspirate fluid in epidemiologic studies of breast disease. Epidemiol. Rev. 1993, 15, 188–195. [Google Scholar] [CrossRef] [PubMed]
- Khodayar-Pardo, P.; Mira-Pascual, L.; Collado, M.C.; Martinez-Costa, C. Impact of lactation stage, gestational age and mode of delivery on breast milk microbiota. J. Perinatol. 2014, 34, 599–605. [Google Scholar] [CrossRef] [PubMed]
- Soto, A.; Martin, V.; Jimenez, E.; Mader, I.; Rodriguez, J.M.; Fernandez, L. Lactobacilli and bifidobacteria in human breast milk: Influence of antibiotherapy and other host and clinical factors. J. Pediatr. Gastroenterol. Nutr. 2014, 59, 78–88. [Google Scholar] [CrossRef] [PubMed]
- Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E. Maternal weight and excessive weight gain during pregnancy modify the immunomodulatory potential of breast milk. Pediatr. Res. 2012, 72, 77–85. [Google Scholar] [CrossRef] [PubMed]
- Turnbaugh, P.J.; Ley, R.E.; Mahowald, M.A.; Magrini, V.; Mardis, E.R.; Gordon, J.I. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature 2006, 444, 1027–1131. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, C.; McMillan, A.; Angelini, M.; Gloor, G.B.; Sumarah, M.; Burton, J.P.; Reid, G. Effect of chemotherapy on the microbiota and metabolome of human milk, a case report. Microbiome 2014, 2, 24. [Google Scholar] [CrossRef] [PubMed]
- Gronlund, M.M.; Gueimonde, M.; Laitinen, K.; Kociubinski, G.; Gronroos, T.; Salminen, S.; Isolauri, E. Maternal breast-milk and intestinal bifidobacteria guide the compositional development of the bifidobacterium microbiota in infants at risk of allergic disease. Clin. Exp. Allergy 2007, 37, 1764–1772. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Maldonado, A.; Martin, V.; Mandomando, I.; Fumado, V.; Metzner, K.J.; Sacoor, C.; Fernandez, L.; Macete, E.; Alonso, P.L.; et al. Breast milk and gut microbiota in african mothers and infants from an area of high hiv prevalence. PLoS ONE 2013, 8, e80299. [Google Scholar] [CrossRef] [PubMed]
- Olivares, M.; Albrecht, S.; De Palma, G.; Ferrer, M.D.; Castillejo, G.; Schols, H.A.; Sanz, Y. Human milk composition differs in healthy mothers and mothers with celiac disease. Eur. J. Nutr. 2015, 54, 119–128. [Google Scholar] [CrossRef] [PubMed]
- Boix-Amoros, A.; Collado, M.C.; Mira, A. Relationship between milk microbiota, bacterial load, macronutrients, and human cells during lactation. Front. Microbiol. 2016, 7, 492. [Google Scholar] [CrossRef] [PubMed]
- Kumar, H.; du Toit, E.; Kulkarni, A.; Aakko, J.; Linderborg, K.M.; Zhang, Y.; Nicol, M.P.; Isolauri, E.; Yang, B.; Collado, M.C.; et al. Distinct patterns in human milk microbiota and fatty acid profiles across specific geographic locations. Front. Microbiol. 2016, 7, 1619. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Mancabelli, L.; Lugli, G.A.; Duranti, S.; Turroni, F.; Ferrario, C.; Mangifesta, M.; Viappiani, A.; Ferretti, P.; Gorfer, V.; et al. Exploring vertical transmission of bifidobacteria from mother to child. Appl. Environ. Microbiol. 2015, 81, 7078–7087. [Google Scholar] [CrossRef] [PubMed]
- Solís, G.; de los Reyes-Gavilan, C.G.; Fernández, N.; Margolles, A.; Gueimonde, M. Establishment and development of lactic acid bacteria and bifidobacteria microbiota in breast-milk and the infant gut. Anaerobe 2010, 16, 307–310. [Google Scholar] [CrossRef] [PubMed]
- Walker, W.A.; Iyengar, R.S. Breast milk, microbiota, and intestinal immune homeostasis. Pediatr. Res. 2015, 77, 220–228. [Google Scholar] [CrossRef] [PubMed]
- Azad, M.B.; Konya, T.; Maughan, H.; Guttman, D.S.; Field, C.J.; Chari, R.S.; Sears, M.R.; Becker, A.B.; Scott, J.A.; Kozyrskyj, A.L. Gut microbiota of healthy canadian infants: Profiles by mode of delivery and infant diet at 4 months. Can. Med. Assoc. J. 2013, 185, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Olivares, M.; Díaz-Ropero, M.P.; Martín, R.; Rodríguez, J.M.; Xaus, J. Antimicrobial potential of four lactobacillus strains isolated from breast milk. J. Appl. Microbiol. 2006, 101, 72–79. [Google Scholar] [CrossRef] [PubMed]
- Fons, A.G.; Tuomo Karjalainen, M. Mechanisms of colonisation and colonisation resistance of the digestive tract part 2: Bacteria/bacteria interactions. Microb. Ecol. Health Dis. 2000, 12, 240–246. [Google Scholar]
- Perez-Cano, F.J.; Dong, H.; Yaqoob, P. In vitro immunomodulatory activity of lactobacillus fermentum cect5716 and lactobacillus salivarius cect5713: Two probiotic strains isolated from human breast milk. Immunobiology 2010, 215, 996–1004. [Google Scholar] [CrossRef] [PubMed]
- Lara-Villoslada, F.; Olivares, M.; Sierra, S.; Rodriguez, J.M.; Boza, J.; Xaus, J. Beneficial effects of probiotic bacteria isolated from breast milk. Br. J. Nutr. 2007, 98, S96–S100. [Google Scholar] [CrossRef] [PubMed]
- Taylor, A.L.; Dunstan, J.A.; Prescott, S.L. Probiotic supplementation for the first 6 months of life fails to reduce the risk of atopic dermatitis and increases the risk of allergen sensitization in high-risk children: A randomized controlled trial. J. Allergy Clin. Immunol. 2007, 119, 184–191. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, T.R.; Jakobsson, T.; Böttcher, M.F.; Fredrikson, M.; Jenmalm, M.C.; Björkstén, B.; Oldaeus, G. Probiotics in prevention of ige-associated eczema: A double-blind, randomized, placebo-controlled trial. J. Allergy Clin. Immunol. 2007, 119, 1174–1180. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Kwon, J.H.; Ahn, S.H.; Lee, S.I.; Han, Y.S.; Choi, Y.O.; Lee, S.Y.; Ahn, K.M.; Ji, G.E. Effect of probiotic mix (bifidobacterium bifidum, bifidobacterium lactis, lactobacillus acidophilus) in the primary prevention of eczema: A double-blind, randomized, placebo-controlled trial. Pediatr. Allergy Immunol. 2010, 21, e386–e393. [Google Scholar] [CrossRef] [PubMed]
- Prokopakis, E.; Vardouniotis, A.; Kawauchi, H.; Scadding, G.; Georgalas, C.; Hellings, P.; Velegrakis, G.; Kalogjera, L. The pathophysiology of the hygiene hypothesis. Int. J. Pediatr. Otorhinolaryngol. 2013, 77, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Kirjavainen, P.V.; Apostolou, E.; Arvola, T.; Salminen, S.J.; Gibson, G.R.; Isolauri, E. Characterizing the composition of intestinal microflora as a prospective treatment target in infant allergic disease. FEMS Immunol. Med. Microbiol. 2001, 32, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Rautava, S. Early microbial contact, the breast milk microbiome and child health. J. Dev. Orig. Health Dis. 2016, 7, 5–14. [Google Scholar] [CrossRef] [PubMed]
- Maldonado, J.; Lara-Villoslada, F.; Sierra, S.; Sempere, L.; Gómez, M.; Rodriguez, J.M.; Boza, J.; Xaus, J.; Olivares, M. Safety and tolerance of the human milk probiotic strain lactobacillus salivarius cect5713 in 6-month-old children. Nutrition 2010, 26, 1082–1087. [Google Scholar] [CrossRef] [PubMed]
- Hassan, Z.; Mustafa, S.; Rahim, R.A.; Isa, N.M. Anti-breast cancer effects of live, heat-killed and cytoplasmic fractions of enterococcus faecalis and staphylococcus hominis isolated from human breast milk. IN Vitro Cell. Dev. Biol. Anim. 2016, 52, 337–348. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.Y.; Woo, H.J.; Kim, Y.S.; Kim, K.H.; Lee, H.J. Cell cycle dysregulation induced by cytoplasm of lactococcus lactis ssp lactis in snuc2a, a colon cancer cell line. Nutr. Cancer 2003, 46, 197–201. [Google Scholar] [CrossRef] [PubMed]
- Delgado, S.; Arroyo, R.; Martin, R.; Rodriguez, J.M. Pcr-dgge assessment of the bacterial diversity of breast milk in women with lactational infectious mastitis. BMC Infect. Dis. 2008, 8, 51. [Google Scholar] [CrossRef] [PubMed]
- Foxman, B.; D’Arcy, H.; Gillespie, B.; Bobo, J.K.; Schwartz, K. Lactation mastitis: Occurrence and medical management among 946 breastfeeding women in the united states. Am. J. Epidemiol. 2002, 155, 103–114. [Google Scholar] [CrossRef] [PubMed]
- Cullinane, M.; Amir, L.H. In response to “microbial diversity in milk of women with mastitis: Potential role of coagulase-negative staphylococci, viridans group streptococci, and corynebacteria”. J. Hum. Lact. 2017, 33, 815–816. [Google Scholar] [CrossRef] [PubMed]
- Delgado, S.; Arroyo, R.; Jimenez, E.; Marin, M.L.; del Campo, R.; Fernandez, L.; Rodriguez, J.M. Staphylococcus epidermidis strains isolated from breast milk of women suffering infectious mastitis: Potential virulence traits and resistance to antibiotics. BMC Microbiol. 2009, 9, 82. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, R.; Martín, V.; Maldonado, A.; Jiménez, E.; Fernández, L.; Rodríguez, J.M. Treatment of infectious mastitis during lactation: Antibiotics versus oral administration of lactobacilli isolated from breast milk. Clin. Infect. Dis. 2010, 50, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Patel, S.H.; Vaidya, Y.H.; Patel, R.J.; Pandit, R.J.; Joshi, C.G.; Kunjadiya, A.P. Culture independent assessment of human milk microbial community in lactational mastitis. Sci. Rep. 2017, 7, 7804. [Google Scholar] [CrossRef] [PubMed]
- Bender, J.M.; Li, F.; Martelly, S.; Byrt, E.; Rouzier, V.; Leo, M.; Tobin, N.; Pannaraj, P.S.; Adisetiyo, H.; Rollie, A.; et al. Maternal hiv infection influences the microbiome of hiv uninfected infants. Sci. Transl. Med. 2016, 8, 100–349. [Google Scholar] [CrossRef] [PubMed]
- Urbaniak, C.; Gloor, G.B.; Brackstone, M.; Scott, L.; Tangney, M.; Reid, G. The microbiota of breast tissue and its association with breast cancer. Appl. Environ. Microbiol. 2016, 82, 5039–5048. [Google Scholar] [CrossRef] [PubMed]
- Belkaid, Y.; Hand, T. Role of the microbiota in immunity and inflammation. Cell 2014, 157, 121–141. [Google Scholar] [CrossRef] [PubMed]
- Xuan, C.; Shamonki, J.M.; Chung, A.; DiNome, M.L.; Chung, M.; Sieling, P.A.; Lee, D.J. Microbial dysbiosis is associated with human breast cancer. PloS ONE 2014, 9, e83744. [Google Scholar] [CrossRef] [PubMed]
- Sheikh, K. The breast has its own microbiome. Sci. Am. 2016, 20, 947–957. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Factors Influencing Human Breast Milk Bacteriome | Bacteriome | References |
|---|---|---|
| Mode of delivery | ||
| Caesarean section | ↓ Bifidobacterium spp., ↑ Proteobacteria, ↓ Lactobacillus spp. (L. fermentum and L. salivarius), ↓ Leuconostocaceae, ↑ Carnobacteriaceae | [18,34,35] |
| Vaginal delivery | ↑ Bifidobacterium spp., ↑ Lactobacillus spp. | [34,35] |
| Maternal weight | ||
| Overweight mothers | ↑ Staphylococcus, ↓ Bifidobacterium | [36] |
| Obese mothers | Less diverse bacterial community | [18] |
| Antibiotic and Chemotherapy | ||
| Perinatal antibiotics usage | ↓ Lactobacillus/Bifidobacterium | [35] |
| Chemotherapy | ↓ Bifidobacterium, ↓ Eubacterium, ↓ Staphylococcus and ↓ Cloacibacterium | [38] |
| Maternal health | ||
| Allergy | ↓ Bifidobacterium | [39] |
| Celiac disease | ↓ Bifidobacterium spp., ↓ Bacteroides fragilis group | [41] |
| HIV | ↑ Bacterial diversity, ↑ Lactobacillus spp. | [40] |
| Geographical location | ||
| Spanish women | ↑ Bacteroidetes | [43] |
| Chinese women | ↑ Actinobacteria | [43] |
| Lactation stage | ||
| Colostrum | ↑ Bacterial diversity, ↓ Bifidobacterium spp., ↓ Total bacterial count | [18] |
| Transition milk | ↑ Bifidobacterium, ↑ Enterococcus spp. counts, ↑ Total bacteria count | [34,42] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ojo-Okunola, A.; Nicol, M.; Du Toit, E. Human Breast Milk Bacteriome in Health and Disease. Nutrients 2018, 10, 1643. https://doi.org/10.3390/nu10111643
Ojo-Okunola A, Nicol M, Du Toit E. Human Breast Milk Bacteriome in Health and Disease. Nutrients. 2018; 10(11):1643. https://doi.org/10.3390/nu10111643
Chicago/Turabian StyleOjo-Okunola, Anna, Mark Nicol, and Elloise Du Toit. 2018. "Human Breast Milk Bacteriome in Health and Disease" Nutrients 10, no. 11: 1643. https://doi.org/10.3390/nu10111643
APA StyleOjo-Okunola, A., Nicol, M., & Du Toit, E. (2018). Human Breast Milk Bacteriome in Health and Disease. Nutrients, 10(11), 1643. https://doi.org/10.3390/nu10111643