Bioactivity, Health Benefits, and Related Molecular Mechanisms of Curcumin: Current Progress, Challenges, and Perspectives



Abstract
1. Introduction
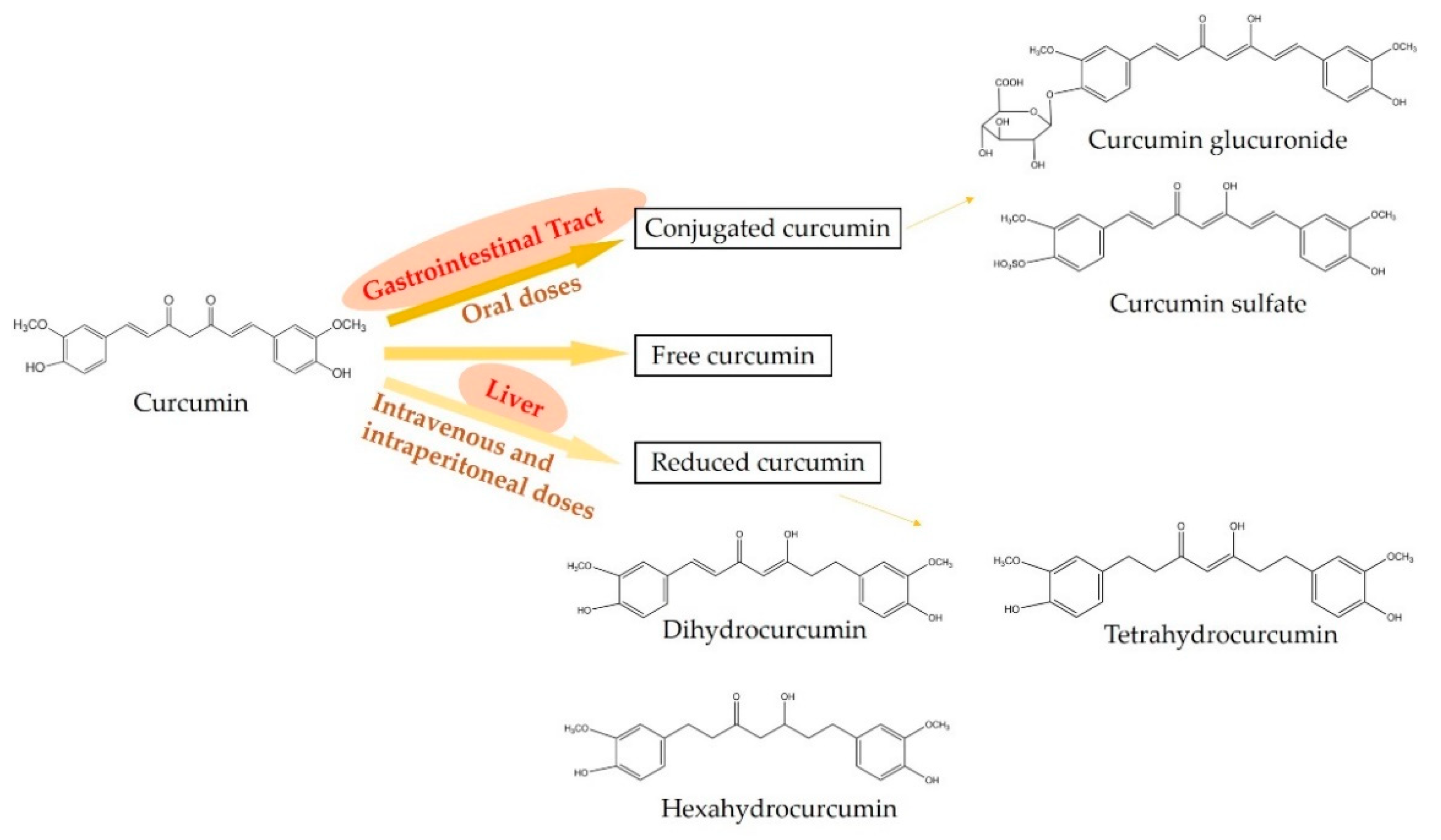
2. The Metabolism of Curcumin
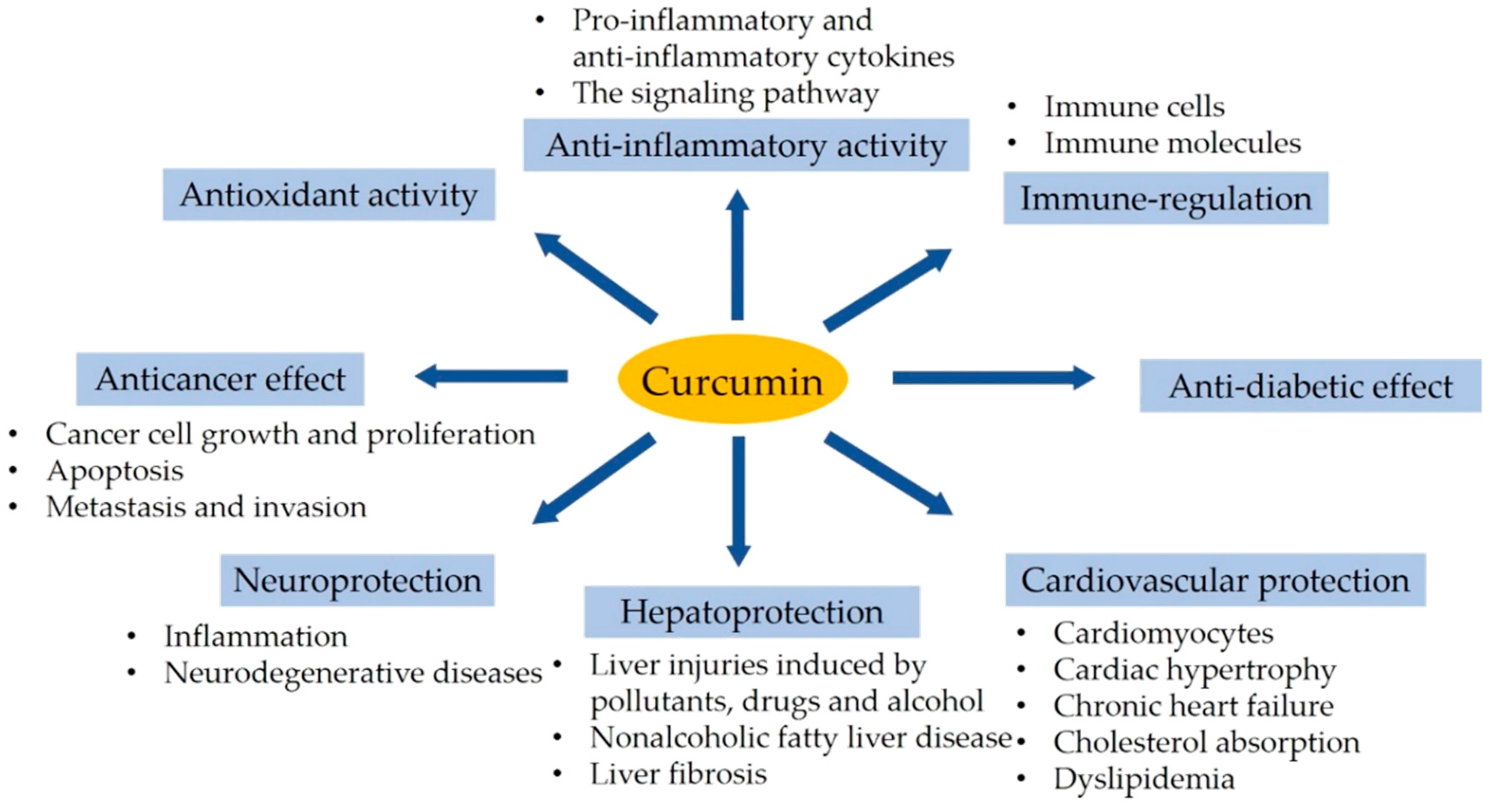
3. Bioactivity of Curcumin
3.1. Antioxidant Activity
3.2. Anti-Inflammatory Activity
3.2.1. Regulation of Pro-Inflammatory and Anti-Inflammatory Cytokines
3.2.2. Regulating Signaling Pathways Associated with Inflammation
3.3. Immune-Regulatory Activity
4. Health Benefits of Curcumin
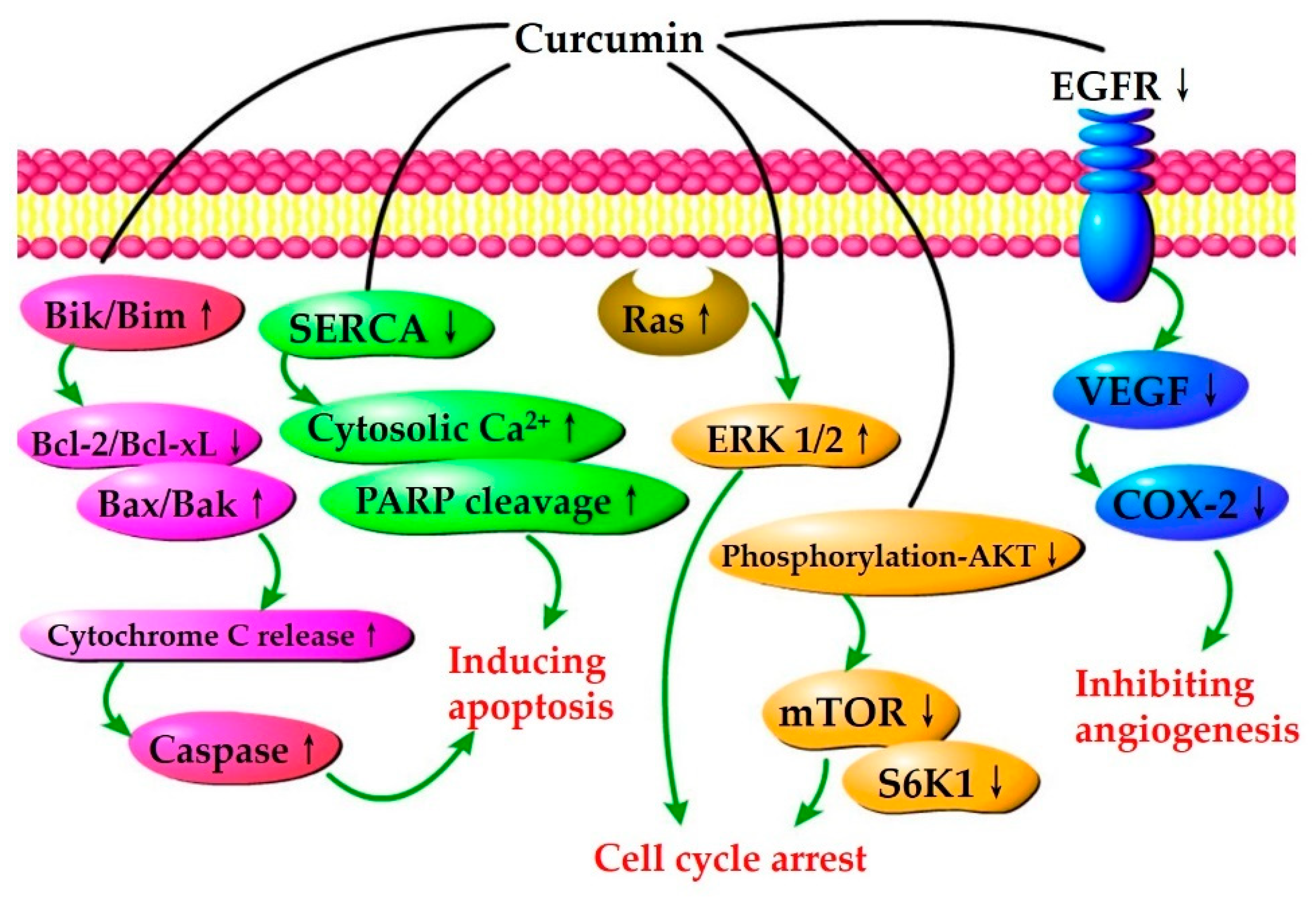
4.1. Anticancer Effect
4.1.1. Inhibition of Cancer Cell Growth and Proliferation
4.1.2. Induction of Cancer Cell Apoptosis
4.1.3. Suppression of Cancer Cell Metastasis and Invasion
4.2. Hepatoprotection
4.2.1. Liver Injuries Induced by Pollutants, Drugs, and Alcohol
4.2.2. Nonalcoholic Fatty Liver Disease
4.2.3. Liver Fibrosis
4.3. Neuroprotection
4.4. Cardiovascular Protection
4.5. Anti-Diabetic Effect
4.6. Other Health Benefits
5. Clinical Trials
6. Challenges and Perspectives
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Maheshwari, R.K.; Singh, A.K.; Gaddipati, J.; Srimal, R.C. Multiple biological activities of curcumin: A short review. Life Sci. 2006, 78, 2081–2087. [Google Scholar] [CrossRef] [PubMed]
- Nelson, K.M.; Dahlin, J.L.; Bisson, J.; Graham, J.; Pauli, G.F.; Walters, M.A. The essential medicinal chemistry of curcumin. J. Med. Chem. 2017, 60, 1620–1637. [Google Scholar] [CrossRef] [PubMed]
- Awasthi, M.; Singh, S.; Pandey, V.P.; Dwivedi, U.N. Curcumin: Structure-activity relationship towards its role as a versatile multi-targeted therapeutics. Mini Rev. Org. Chem. 2017, 14, 311–332. [Google Scholar] [CrossRef]
- Wang, R.; Li, J.B.; Zhao, Y.L.; Li, Y.P.; Yin, L. Investigating the therapeutic potential and mechanism of curcumin in breast cancer based on RNA sequencing and bioinformatics analysis. Breast Cancer 2018, 25, 206–212. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wang, T.; Wang, M. Investigation of the anti-cataractogenic mechanisms of curcumin through in vivo and in vitro studies. BMC Ophthalmol. 2018, 18, 48. [Google Scholar] [CrossRef] [PubMed]
- Pulido-Moran, M.; Moreno-Fernandez, J.; Ramirez-Tortosa, C.; Ramirez-Tortosa, M.C. Curcumin and health. Molecules 2016, 21, 264. [Google Scholar] [CrossRef] [PubMed]
- Jankun, J.; Wyganowska-Swiatkowska, M.; Dettlaff, K.; Jelinska, A.; Surdacka, A.; Watrobska-Swietlikowska, D.; Skrzypczak-Jankun, E. Determining whether curcumin degradation/condensation is actually bioactivation (review). Int. J. Mol. Med. 2016, 37, 1151–1158. [Google Scholar] [CrossRef] [PubMed]
- Holder, G.M.; Plummer, J.L.; Ryan, A.J. The metabolism and excretion of curcumin (1,7-bis-(4-hydroxy-3-methoxyphenyl)-1,6-heptadiene-3,5-dione) in the rat. Xenobiotica 1978, 8, 761–768. [Google Scholar] [CrossRef] [PubMed]
- Asai, A.; Miyazawa, T. Occurrence of orally administered curcuminoid as glucuronide and glucuronide/sulfate conjugates in rat plasma. Life Sci. 2000, 67, 2785–2793. [Google Scholar] [CrossRef]
- Hoehle, S.I.; Pfeiffer, E.; Metzler, M. Glucuronidation of curcuminoids by human microsomal and recombinant UDP-glucuronosyltransferases. Mol. Nutr. Food Res. 2007, 51, 932–938. [Google Scholar] [CrossRef] [PubMed]
- Prasad, S.; Tyagi, A.K.; Aggarwal, B.B. Recent developments in delivery, bioavailability, absorption and metabolism of curcumin: The golden pigment from golden spice. Cancer Res. Treat. 2014, 46, 2–18. [Google Scholar] [CrossRef] [PubMed]
- Tsuda, T. Curcumin as a functional food-derived factor: Degradation products, metabolites, bioactivity, and future perspectives. Food Funct. 2018, 9, 705–714. [Google Scholar] [CrossRef] [PubMed]
- Hoehle, S.I.; Pfeiffer, E.; Solyom, A.M.; Metzler, M. Metabolism of curcuminoids in tissue slices and subcellular fractions from rat liver. J. Agric. Food Chem. 2006, 54, 756–764. [Google Scholar] [CrossRef] [PubMed]
- Pfeiffer, E.; Hoehle, S.I.; Walch, S.G.; Riess, A.; Solyom, A.M.; Metzler, M. Curcuminoids form reactive glucuronides in vitro. J. Agric. Food Chem. 2007, 55, 538–544. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.Y.; Cao, S.J.; Zhang, Q.; Zhang, H.Y.; Fan, Y.Q.; Qiu, F.; Kang, N. Biological and pharmacological effects of hexahydrocurcumin, a metabolite of curcumin. Arch. Biochem. Biophys. 2018, 646, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Dempe, J.S.; Scheerle, R.K.; Pfeiffer, E.; Metzler, M. Metabolism and permeability of curcumin in cultured Caco-2 cells. Mol. Nutr. Food Res. 2013, 57, 1543–1549. [Google Scholar] [CrossRef] [PubMed]
- Metzler, M.; Pfeiffer, E.; Schulz, S.I.; Dempe, J.S. Curcumin uptake and metabolism. BioFactors 2013, 39, 14–20. [Google Scholar] [CrossRef] [PubMed]
- Dempe, J.S.; Pfeiffer, E.; Grimm, A.S.; Metzler, M. Metabolism of curcumin and induction of mitotic catastrophe in human cancer cells. Mol. Nutr. Food Res. 2008, 52, 1074–1081. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, M.R.; Jardim, F.R.; Setzer, W.N.; Nabavi, S.M.; Nabavi, S.F. Curcumin, mitochondrial biogenesis, and mitophagy: Exploring recent data and indicating future needs. Biotechnol. Adv. 2016, 34, 813–826. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Zheng, J.; Hu, H.; Lee, J.; Zeng, S. Application of ultra-performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry to identify curcumin metabolites produced by human intestinal bacteria. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2015, 985, 38–47. [Google Scholar] [CrossRef] [PubMed]
- An, C.-Y.; Sun, Z.-Z.; Shen, L.; Ji, H.-F. Biotransformation of food spice curcumin by gut bacterium Bacillus megaterium DCMB-002 and its pharmacological implications. Food Nutr. Res. 2017, 61. [Google Scholar] [CrossRef]
- Burapan, S.; Kim, M.; Han, J. Curcuminoid demethylation as an alternative metabolism by human intestinal microbiota. J. Agric. Food Chem. 2017, 65, 3306–3311. [Google Scholar] [CrossRef] [PubMed]
- Asouri, M.; Ataee, R.; Ahmadi, A.A.; Amini, A.; Moshaei, M.R. Antioxidant and free radical scavenging activities of curcumin. Asian J. Chem. 2013, 25, 7593–7595. [Google Scholar]
- Derochette, S.; Franck, T.; Mouithys-Mickalad, A.; Ceusters, J.; Deby-Dupont, G.; Lejeune, J.P.; Neven, P.; Serteyn, D. Curcumin and resveratrol act by different ways on NADPH oxidase activity and reactive oxygen species produced by equine neutrophils. Chem.-Biol. Interact. 2013, 206, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.T.; Yang, Z.H.; Yu, L.Y.; Ren, Y.Y.; Huang, Q.X.; Liu, Q.; Ma, X.Y.; Chen, Z.K.; Wang, Z.B.; Zheng, X. Synthesis and antioxidant activity of curcumin analogs. J. Asian Nat. Prod. Res. 2017, 19, 489–503. [Google Scholar] [CrossRef] [PubMed]
- Dikmen, M.; Kaya-Tilki, E.; Engur, S.; Ozturk, Y. Neuritogenic activity of epigallocatechin gallate and curcumin combination on rat adrenal pheochromocytoma cells. Fresenius Environ. Bull. 2017, 26, 4726–4733. [Google Scholar]
- Tapia, E.; Sanchez-Lozada, L.G.; Garcia-Nino, W.R.; Garcia, E.; Cerecedo, A.; Garcia-Arroyo, F.E.; Osorio, H.; Arellano, A.; Cristobal-Garcia, M.; Loredo, M.L.; et al. Curcumin prevents maleate-induced nephrotoxicity: Relation to hemodynamic alterations, oxidative stress, mitochondrial oxygen consumption and activity of respiratory complex I. Free Radic. Res. 2014, 48, 1342–1354. [Google Scholar] [CrossRef] [PubMed]
- Dai, J.; Gu, L.; Su, Y.; Wang, Q.; Zhao, Y.; Chen, X.; Deng, H.; Li, W.; Wang, G.; Li, K. Inhibition of curcumin on influenza a virus infection and influenzal pneumonia via oxidative stress, TLR2/4, p38/jnk mapk and NF-κB pathways. Int. Immunopharmacol. 2018, 54, 177–187. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.S.; Tang, S.S.; Li, D.W.; Zhao, K.N.; Xiao, X.L. Curcumin attenuates quinocetone-induced oxidative stress and genotoxicity in human hepatocyte l02 cells. Toxicol. Mech. Methods 2015, 25, 340–346. [Google Scholar] [CrossRef] [PubMed]
- Maugeri, A.; Mazzone, M.G.; Giuliano, F.; Vinciguerra, M.; Basile, G.; Barchitta, M.; Agodi, A. Curcumin modulates DNA methyltransferase functions in a cellular model of diabetic retinopathy. Oxid. Med. Cell. Longev. 2018, 2018, 5407482. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Rizvi, S.I. Role of curcumin in modulating plasma PON1 arylesterase activity and susceptibility to LDL oxidation in oxidatively challenged Wistar rats. Lett. Drug Des. Discov. 2015, 12, 319–323. [Google Scholar] [CrossRef]
- Joshi, D.; Mittal, D.K.; Shukla, S.; Srivastav, S.K.; Dixit, V.A. Curcuma longa Linn. Extract and curcumin protect CYP 2E1 enzymatic activity against mercuric chloride-induced hepatotoxicity and oxidative stress: A protective approach. Exp. Toxicol. Pathol. 2017, 69, 373–382. [Google Scholar] [CrossRef] [PubMed]
- El-Bahr, S.M. Effect of curcumin on hepatic antioxidant enzymes activities and gene expressions in rats intoxicated with aflatoxin b1. Phytother. Res. 2015, 29, 134–140. [Google Scholar] [CrossRef] [PubMed]
- Goc, Z.; Semla, M.; Kapusta, E.; Gren, A.; Muchacka, R.; Lukacova, J. The effect of curcumin on the activity of antioxidant enzymes in the liver, pancreas and kidneys of Swiss mice. In Proceedings of the 12th International Scientific Conference on Animal Physiology, Boretice, Czech Republic, 13–15 June 2016. [Google Scholar]
- Altintoprak, N.; Kar, M.; Acar, M.; Berkoz, M.; Muluk, N.B.; Cingi, C. Antioxidant activities of curcumin in allergic rhinitis. Eur. Arch. Otorhinolaryngol. 2016, 273, 3765–3773. [Google Scholar] [CrossRef] [PubMed]
- Singh, P.; Kesharwani, R.K.; Misra, K.; Rizvi, S.I. Modulation of erythrocyte plasma membrane redox system activity by curcumin. Biochem. Res. Int. 2016, 2016, 6025245. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.P.; Li, Y.; Meng, X.; Zhou, T.; Zhou, Y.; Zheng, J.; Zhang, J.J.; Li, H.B. Natural antioxidants in foods and medicinal plants: Extraction, assessment and resources. Int. J. Mol. Sci. 2017, 18, 96. [Google Scholar] [CrossRef] [PubMed]
- Pompella, A.; Sies, H.; Wacker, R.; Brouns, F.; Grune, T.; Biesalski, H.K.; Frank, J. The use of total antioxidant capacity as surrogate marker for food quality and its effect on health is to be discouraged. Nutrition 2014, 30, 791–793. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, H.Y.; Zhu, R.R.; Liu, Q.; Fei, J.; Wang, S.L. Anti-inflammatory activity of curcumin-loaded solid lipid nanoparticles in IL-1 beta transgenic mice subjected to the, lipopolysaccharide-induced sepsis. Biomaterials 2015, 53, 475–483. [Google Scholar] [CrossRef] [PubMed]
- Antoine, F.; Girard, D. Curcumin increases gelatinase activity in human neutrophils by a p38 mitogen-activated protein kinase (MAPK)-independent mechanism. J. Immunotoxicol. 2015, 12, 188–193. [Google Scholar] [CrossRef] [PubMed]
- Li, H.Y.; Yang, M.; Li, Z.; Meng, Z. Curcumin inhibits angiotensin II-induced inflammation and proliferation of rat vascular smooth muscle cells by elevating PPAR-gamma activity and reducing oxidative stress. Int. J. Mol. Med. 2017, 39, 1307–1316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.L.; Liu, Z.G.; Wu, J.Z.; Bai, B.; Chen, H.J.; Xiao, Z.X.; Chen, L.F.; Zhao, Y.J.; Lum, H.; Wang, Y.; et al. New MD2 inhibitors derived from curcumin with improved anti-inflammatory activity. Eur. J. Med. Chem. 2018, 148, 291–305. [Google Scholar] [CrossRef] [PubMed]
- Wu, N.C.; Wang, J.J. Curcumin attenuates liver warm ischemia and reperfusion-induced combined restrictive and obstructive lung disease by reducing matrix metalloprotease 9 activity. Transplant. Proc. 2014, 46, 1135–1138. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, V.H.; Nazli, A.; Dizzell, S.E.; Mueller, K.; Kaushic, C. The anti-inflammatory activity of curcumin protects the genital mucosal epithelial barrier from disruption and blocks replication of HIV-1 and HSV-2. PLoS ONE 2015, 10, e0124903. [Google Scholar] [CrossRef] [PubMed]
- Kloesch, B.; Gober, L.; Loebsch, S.; Vcelar, B.; Helson, L.; Steiner, G. In vitro study of a liposomal curcumin formulation (lipocurc (TM)): Toxicity and biological activity in synovial fibroblasts and macrophages. In Vivo 2016, 30, 413–419. [Google Scholar] [PubMed]
- Esatbeyoglu, T.; Ulbrich, K.; Rehberg, C.; Rohn, S.; Rimbach, G. Thermal stability, antioxidant, and anti-inflammatory activity of curcumin and its degradation product 4-vinyl guaiacol. Food Funct. 2015, 6, 887–893. [Google Scholar] [CrossRef] [PubMed]
- Song, Z.L.; Revelo, X.; Shao, W.J.; Tian, L.L.; Zeng, K.J.; Lei, H.; Sun, H.S.; Woo, M.; Winer, D.; Jin, T. Dietary curcumin intervention targets mouse white adipose tissue inflammation and brown adipose tissue ucp1 expression. Obesity 2018, 26, 547–558. [Google Scholar] [CrossRef] [PubMed]
- Hui-Yin, Y.; Ahmad, N.; Azmi, N.; Makmor-Bakry, M. Curcumin: The molecular mechanisms of action in inflammation and cell death during kainate-induced epileptogenesis. Indian J. Pharm. Educ. 2018, 52, 32–41. [Google Scholar] [CrossRef]
- Wang, J.; Kang, Y.X.; Pan, W.; Lei, W.; Feng, B.; Wang, X.J. Enhancement of anti-inflammatory activity of curcumin using phosphatidylserine-containing nanoparticles in cultured macrophages. Int. J. Mol. Sci. 2016, 17, 969. [Google Scholar] [CrossRef] [PubMed]
- Hunter, C.J.; De Plaen, I.G. Inflammatory signaling in nec: Role of NF-kappab, cytokines and other inflammatory mediators. Pathophysiol. 2014, 21, 55–65. [Google Scholar] [CrossRef] [PubMed]
- Kao, N.J.; Hu, J.Y.; Wu, C.S.; Kong, Z.L. Curcumin represses the activity of inhibitor-kappa b kinase in dextran sulfate sodium-induced colitis by S-nitrosylation. Int. Immunopharmacol. 2016, 38, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Dai, C.S.; Ciccotosto, G.D.; Cappai, R.; Tang, S.S.; Li, D.W.; Xie, S.L.; Xiao, X.L.; Velkov, T. Curcumin attenuates colistin-induced neurotoxicity in N2a cells via anti-inflammatory activity, suppression of oxidative stress, and apoptosis. Mol. Neurobiol. 2018, 55, 421–434. [Google Scholar] [CrossRef] [PubMed]
- Cheng, F.; Chen, Y.H.; Zhan, Z.; Liu, Y.; Hu, P.; Ren, H.; Tang, H.D.; Peng, M.L. Curc-MPEG454, a pegylated curcumin derivative, improves anti-inflammatory and antioxidant activities: A comparative study. Inflammation 2018, 41, 579–594. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.F.; Li, Y.L.; Yue, Y.; Zhang, K.; Chen, Q.; Wang, H.Q.; Lu, Y.J.; Huang, M.T.; Zheng, X.; Du, Z.Y. Synthesis and biological evaluation of curcumin derivatives containing NSAIDs for their anti-inflammatory activity. Bioorg. Med. Chem. Lett. 2015, 25, 3044–3051. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.S.; Chen, L.Z.; Zhou, H.P.; Liu, X.H.; Chen, F.H. Diarylpentadienone derivatives (curcumin analogues): Synthesis and anti-inflammatory activity. Bioorg. Med. Chem. Lett. 2017, 27, 1803–1807. [Google Scholar] [CrossRef] [PubMed]
- Yang, H.X.; Du, Z.Y.; Wang, W.C.; Song, M.Y.; Sanidad, K.; Sukamtoh, E.; Zheng, J.; Tian, L.; Xiao, H.; Liu, Z.H.; et al. Structure-activity relationship of curcumin: Role of the methoxy group in anti-inflammatory and anticolitis effects of curcumin. J. Agric. Food Chem. 2017, 65, 4509–4515. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, E.; Momtazi, A.A.; Johnston, T.P.; Sahebkar, A. Therapeutic effects of curcumin in inflammatory and immune-mediated diseases: A nature-made jack-of-all-trades? J. Cell. Physiol. 2018, 233, 830–848. [Google Scholar] [CrossRef] [PubMed]
- Shakeri, F.; Boskabady, M.H. Anti-inflammatory, antioxidant, and immunomodulatory effects of curcumin in ovalbumin-sensitized rat. BioFactors 2017, 43, 567–576. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Luo, B.; Shi, R.; Han, C.; Zhang, Z.; Xiong, J.; Jiang, M.; Zhang, Z. Curcumin ameliorates rat experimental autoimmune neuritis. J. Neurosci. Res. 2014, 92, 743–750. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; He, C.M.; He, J.Y.; Chu, J.; Liu, H.P.; Deng, X.Y. Curcumin-mediated bone marrow mesenchymal stem cell sheets create a favorable immune microenvironment for adult full-thickness cutaneous wound healing. Stem Cell. Res. Ther. 2018, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Afia, M.; Alshehri, M.; Alfaifi, M.; Shakor, A.B.A. Repressive effect of curcumin against 2-amino-3-methylimidazo 4, 5-f quinoline induced hepato- and immunotoxicity in mice. Indian J. Exp. Biol. 2017, 55, 365–371. [Google Scholar]
- Wang, S.; Li, H.; Zhang, M.; Yue, L.T.; Wang, C.C.; Zhang, P.; Liu, Y.; Duan, R.S. Curcumin ameliorates experimental autoimmune myasthenia gravis by diverse immune cells. Neurosci. Lett. 2016, 626, 25–34. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, L.Y.; Shen, Y.; Tan, T.; Xie, N.Z.; Luo, M.; Li, Z.H.; Xie, X.Y. Curcumin ameliorates ischemia-induced limb injury through immunomodulation. Med. Sci. Monit. 2016, 22, 2035–2042. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Sasmal, D.; Jadav, S.S.; Sharma, N. Mechanism of immunoprotective effects of curcumin in DLM-induced thymic apoptosis and altered immune function: An in silico and in vitro study. Immunopharmacol. Immunotoxicol. 2015, 37, 488–498. [Google Scholar] [CrossRef] [PubMed]
- Bose, S.; Panda, A.K.; Mukherjee, S.; Sa, G. Curcumin and tumor immune-editing: Resurrecting the immune system. Cell Div. 2015, 10, 6. [Google Scholar] [CrossRef] [PubMed]
- Mahmoud, H.K.; Al-Sagheer, A.A.; Reda, F.M.; Mahgoub, S.A.; Ayyat, M.S. Dietary curcumin supplement influence on growth, immunity, antioxidant status, and resistance to aeromonas hydrophila in oreochromis niloticus. Aquaculture 2017, 475, 16–23. [Google Scholar] [CrossRef]
- Xun, W.J.; Shi, L.G.; Zhou, H.L.; Hou, G.Y.; Cao, T.; Zhao, C.P. Effects of curcumin on growth performance, jejunal mucosal membrane integrity, morphology and immune status in weaned piglets challenged with enterotoxigenic Escherichia coli. Int. Immunopharmacol. 2015, 27, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.; Chung, H.S.; Lee, K.; Lee, H.; Kim, M.; Bae, H. Curcumin attenuates the scurfy-induced immune disorder, a model of ipex syndrome, with inhibiting Th1/Th2/Th17 responses in mice. Phytomedicine 2017, 33, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.L.; Miller, K.D.; Jemal, A. Cancer statistics, 2017. CA. Cancer J. Clin. 2017, 67, 7–30. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.R.; Gao, S.; Yang, Y.X.; Zhao, X.D.; Fan, Y.M.; Ma, W.X.; Yang, D.R.; Yang, A.M.; Yu, Y. Antitumor activity of curcumin by modulation of apoptosis and autophagy in human lung cancer A549 cells through inhibiting PI3K/Akt/mTOR pathway. Oncol. Rep. 2018, 39, 1523–1531. [Google Scholar] [CrossRef] [PubMed]
- Carroll, R.E.; Benya, R.V.; Turgeon, D.K.; Vareed, S.; Neuman, M.; Rodriguez, L.; Kakarala, M.; Carpenter, P.M.; McLaren, C.; Meyskens, F.L., Jr.; et al. Phase II a clinical trial of curcumin for the prevention of colorectal neoplasia. Cancer Prev. Res. 2011, 4, 354–364. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.Y.; Wang, Y.; Zhang, R.K.; Yang, G.W.; Liang, Z.B.; Wang, Z.W.; Zhang, G.H. Curcumin exerts its antitumor activity through regulation of miR-7/Skp2/p21 in nasopharyngeal carcinoma cells. Onco Targets Ther. 2017, 10, 2377–2388. [Google Scholar] [CrossRef] [PubMed]
- Xi, Y.; Gao, H.; Callaghan, M.U.; Fribley, A.M.; Garshott, D.M.; Xu, Z.X.; Zeng, Q.H.; Li, Y.L. Induction of Bcl2-interacting killer, Bik, is mediated for anti-cancer activity of curcumin in human head and neck squamous cell carcinoma cells. J. Cancer 2015, 6, 327–332. [Google Scholar] [CrossRef] [PubMed]
- Yoysungnoen-Chintana, P.; Bhattarakosol, P.; Patumraj, S. Antitumor and antiangiogenic activities of curcumin in cervical cancer xenografts in nude mice. BioMed Res. Int. 2014, 10, 817972. [Google Scholar] [CrossRef] [PubMed]
- Zhan, Y.Z.; Chen, Y.N.; Liu, R.; Zhang, H.; Zhang, Y.M. Potentiation of paclitaxel activity by curcumin in human breast cancer cell by modulating apoptosis and inhibiting EGFR signaling. Arch. Pharm. Res. 2014, 37, 1086–1095. [Google Scholar] [CrossRef] [PubMed]
- Montgomery, A.; Adeyeni, T.; San, K.; Heuertz, R.M.; Ezekiel, U.R. Curcumin sensitizes silymarin to exert synergistic anticancer activity in colon cancer cells. J. Cancer 2016, 7, 1250–1257. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.X.; Su, J.N.; Feng, S.Y.; Wang, L.X.; Yin, X.Y.; Yan, J.Z.; Wang, Z.W. Antitumor activity of curcumin is involved in down-regulation of YAP/TAZ expression in pancreatic cancer cells. Oncotarget 2016, 7, 79062–79074. [Google Scholar] [CrossRef] [PubMed]
- Kumar, P.; Barua, C.C.; Sulakhiya, K.; Sharma, R.K. Curcumin ameliorates cisplatin-induced nephrotoxicity and potentiates its anticancer activity in SD rats: Potential role of curcumin in breast cancer chemotherapy. Front. Pharmacol. 2017, 8, 132. [Google Scholar] [CrossRef] [PubMed]
- Goncalves, V.D.; Ortega, A.A.C.; Guimaraes, M.R.; Curylofo, F.A.; Rossa, C.; Ribeiro, D.A.; Spolidorio, L.C. Chemopreventive activity of systemically administered curcumin on oral cancer in the 4-nitroquinoline 1-oxide model. J. Cell. Biochem. 2015, 116, 787–796. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, S.; Chamani, J.; Rahimi, H.; Azmoodeh, N.; Ghasemi, F.; Abadi, P.H. An in vitro study on curcumin delivery by nano-micelles for esophageal squamous cell carcinoma (KYSE-30). Rep. Biochem. Mol. Biol. 2018, 6, 137–143. [Google Scholar] [PubMed]
- Meena, R.; Kumar, S.; Kumar, R.; Gaharwar, U.S.; Rajamani, P. Plga-ctab curcumin nanoparticles: Fabrication, characterization and molecular basis of anticancer activity in triple negative breast cancer cell lines (MDA-MB-231 cells). Biomed. Pharmacother. 2017, 94, 944–954. [Google Scholar] [CrossRef] [PubMed]
- Francis, A.P.; Murthy, P.B.; Devasena, T. Bis-demethoxy curcumin analog nanoparticles: Synthesis, characterization, and anticancer activity in vitro. J. Nanosci. Nanotechnol. 2014, 14, 4865–4873. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Higuchi, T.; Takeshima, M.; Chen, C.; Nakano, S. Differential anti-tumor activities of curcumin against Ras- and SRC-activated human adenocarcinoma cells. Biochem. Biophys. Res. Commun. 2013, 436, 186–191. [Google Scholar] [CrossRef] [PubMed]
- Rana, C.; Piplani, H.; Vaish, V.; Nehru, B.; Sanyal, S.N. Downregulation of telomerase activity by diclofenac and curcumin is associated with cell cycle arrest and induction of apoptosis in colon cancer. Tumor Biol. 2015, 36, 5999–6010. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, D.; Ganguli, A.; Dastidar, D.G.; Acharya, B.R.; Das, A.; Chakrabarti, G. Apigenin shows synergistic anticancer activity with curcumin by binding at different sites of tubulin. Biochimie 2013, 95, 1297–1309. [Google Scholar] [CrossRef] [PubMed]
- Gao, X.; Wang, B.L.; Wu, Q.J.; Wei, X.W.; Zheng, F.J.; Men, K.; Shi, H.S.; Huang, N.; Wei, Y.Q.; Gong, C.Y. Combined delivery and anti-cancer activity of paclitaxel and curcumin using polymeric micelles. J. Biomed. Nanotechnol. 2015, 11, 578–589. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.M.; Li, J.J.; Shi, Z.; Yang, Y.; Xie, X.; Lee, S.M.; Wang, Y.T.; Leong, K.W.; Chen, M.W. Ph-sensitive polymeric nanoparticles for co-delivery of doxorubicin and curcumin to treat cancer via enhanced pro-apoptotic and anti-angiogenic activities. Acta Biomater. 2017, 58, 349–364. [Google Scholar] [CrossRef] [PubMed]
- Mahmud, M.; Piwoni, A.; Filiczak, N.; Janicka, M.; Gubernator, J. Long-circulating curcumin-loaded liposome formulations with high incorporation efficiency, stability and anticancer activity towards pancreatic adenocarcinoma cell lines in vitro. PLoS ONE 2016, 11, e0167787. [Google Scholar] [CrossRef] [PubMed]
- Ono, M.; Higuchi, T.; Takeshima, M.; Chen, C.; Nakano, S. Antiproliferative and apoptosis-inducing activity of curcumin against human gallbladder adenocarcinoma cells. Anticancer Res. 2013, 33, 1861–1866. [Google Scholar] [PubMed]
- Sahu, B.P.; Hazarika, H.; Bharadwaj, R.; Loying, P.; Baishya, R.; Dash, S.; Das, M.K. Curcumin-docetaxel co-loaded nanosuspension for enhanced anti-breast cancer activity. Expert Opin. Drug Deliv. 2016, 13, 1065–1074. [Google Scholar] [CrossRef] [PubMed]
- Koohpar, Z.K.; Entezari, M.; Movafagh, A.; Hashemi, M. Anticancer activity of curcumin on human breast adenocarcinoma: Role of mcl-1 gene. Iran J. Cancer Prev. 2015, 8, e2331. [Google Scholar] [CrossRef] [PubMed]
- Seo, J.A.; Kim, B.; Dhanasekaran, D.N.; Tsang, B.K.; Song, Y.S. Curcumin induces apoptosis by inhibiting sarco/endoplasmic reticulum Ca2+ ATPase activity in ovarian cancer cells. Cancer Lett. 2016, 371, 30–37. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.S.; Wang, T.S.; Lin, M.C.; Lin, W.W.; Yang, J.J. Inhibition of curcumin on ZAK alpha activity resultant in apoptosis and anchorage-independent growth in cancer cells. Chin. J. Physiol. 2017, 60, 267–274. [Google Scholar] [CrossRef] [PubMed]
- He, Z.-Y.; Shi, C.-B.; Wen, H.; Li, F.-L.; Wang, B.-L.; Wang, J. Upregulation of p53 expression in patients with colorectal cancer by administration of curcumin. Cancer Investig. 2011, 29, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.H.; Yue, J.M.; Sims, M.; Pfeffer, L.M. The curcumin analog EF24 targets NF-kappa B and miRNA-21, and has potent anticancer activity in vitro and in vivo. PLoS ONE 2013, 8, e71130. [Google Scholar] [CrossRef]
- Zhang, W.; Bai, W.; Zhang, W. Mir-21 suppresses the anticancer activities of curcumin by targeting pten gene in human non-small cell lung cancer A549 cells. Clin. Transl. Oncol. 2014, 16, 708–713. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.; Haggag, Y.A.; Lane, M.E.; McCarron, P.A.; Tambuwala, M.M. Polymeric nano-encapsulation of curcumin enhances its anti-cancer activity in breast (MDA-MB231) and lung (A549) cancer cells through reduction in expression of HIF-1 alpha and nuclear p65 (Rel A). Curr. Drug Deliv. 2018, 15, 286–295. [Google Scholar] [CrossRef] [PubMed]
- Balakrishna, A.; Kumar, M.H. Evaluation of synergetic anticancer activity of berberine and curcumin on different models of A549, Hep-G2, MCF-7, Jurkat, and K562 cell lines. BioMed Res. Int. 2015, 2015, 354614. [Google Scholar] [CrossRef] [PubMed]
- Park, B.H.; Lim, J.E.; Jeon, H.G.; Seo, S.I.; Lee, H.M.; Choi, H.Y.; Jeon, S.S.; Jeong, B.C. Curcumin potentiates antitumor activity of cisplatin in bladder cancer cell lines via ROS-mediated activation of ERK1/2. Oncotarget 2016, 7, 63870–63886. [Google Scholar] [CrossRef] [PubMed]
- Orr, W.S.; Denbo, J.W.; Saab, K.R.; Ng, C.Y.; Wu, J.R.; Li, K.; Garner, J.M.; Morton, C.L.; Du, Z.Y.; Pfeffer, L.M.; et al. Curcumin potentiates rhabdomyosarcoma radiosensitivity by suppressing NF-kappa B activity. PLoS ONE 2013, 8, e51309. [Google Scholar] [CrossRef] [PubMed]
- Shakibaei, M.; Kraehe, P.; Popper, B.; Shayan, P.; Goel, A.; Buhrmann, C. Curcumin potentiates antitumor activity of 5-fluorouracil in a 3D alginate tumor microenvironment of colorectal cancer. BMC Cancer 2015, 15, 250. [Google Scholar] [CrossRef] [PubMed]
- Zhang, P.; Lai, Z.L.; Chen, H.F.; Zhang, M.; Wang, A.; Jia, T.; Sun, W.Q.; Zhu, X.M.; Chen, X.F.; Zhao, Z.; et al. Curcumin synergizes with 5-fluorouracil by impairing AMPK/ULK1-dependent autophagy, AKT activity and enhancing apoptosis in colon cancer cells with tumor growth inhibition in xenograft mice. J. Exp. Clin. Cancer Res. 2017, 36, 190. [Google Scholar] [CrossRef] [PubMed]
- Senft, C.; Polacin, M.; Priester, M.; Seifert, V.; Koegel, D.; Weissenberger, J. The nontoxic natural compound curcumin exerts anti-proliferative, anti-migratory, and anti-invasive properties against malignant gliomas. BMC Cancer 2010, 10, 491. [Google Scholar] [CrossRef] [PubMed]
- Deng, Y.; Verron, E.; Rohanizadeh, R. Molecular mechanisms of anti-metastatic activity of curcumin. Anticancer Res. 2016, 36, 5639–5647. [Google Scholar] [CrossRef] [PubMed]
- Li, S.L.; Fang, C.S.; Zhang, J.Q.; Liu, B.L.; Wei, Z.Q.; Fan, X.Q.; Sui, Z.; Tan, Q.Y. Catanionic lipid nanosystems improve pharmacokinetics and anti-lung cancer activity of curcumin. Nanomedicine 2016, 12, 1567–1579. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.L.; Cui, T.; Liu, L.; Wu, Q.J.; Sun, L.; Li, L.; Wang, N.; Gong, C.Y. Improving anti-tumor activity of curcumin by polymeric micelles in thermosensitive hydrogel system in colorectal peritoneal carcinomatosis model. J. Biomed. Nanotechnol. 2015, 11, 1173–1182. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Zhou, Y.; Li, Y.; Xu, D.P.; Li, S.; Li, H.B. Spices for prevention and treatment of cancers. Nutrients 2016, 8, 495. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zheng, J.; Li, Y.; Xu, D.P.; Li, S.; Chen, Y.M.; Li, H.B. Natural polyphenols for prevention and treatment of cancer. Nutrients 2016, 8, 515. [Google Scholar] [CrossRef] [PubMed]
- Abd-Allah, G.A.; El-Bakry, K.A.; Bahnasawy, M.H.; El-Khodary, E.R. Protective effects of curcumin and ginger on liver cirrhosis induced by carbon tetrachloride in rats. Int. J. Pharmacol. 2016, 12, 361–369. [Google Scholar] [CrossRef]
- Nabavi, S.F.; Daglia, M.; Moghaddam, A.H.; Habtemariam, S.; Nabavi, S.M. Curcumin and liver disease: From chemistry to medicine. Compr. Rev. Food Sci. Food Saf. 2014, 13, 62–77. [Google Scholar] [CrossRef]
- Peng, X.Y.; Dai, C.S.; Liu, Q.W.; Li, J.K.; Qiu, J.R. Curcumin attenuates on carbon tetrachloride-induced acute liver injury in mice via modulation of the Nrf2/HO-1 and TGF-1/Smad3 pathway. Molecules 2018, 23, 215. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, S.T.; Das, N.; Ghosh, S.; Ghosh, D.; Chakraborty, S.; Ali, N. Vesicular (liposomal and nanoparticulated) delivery of curcumin: A comparative study on carbon tetrachloride-mediated oxidative hepatocellular damage in rat model. Int. J. Nanomed. 2016, 11, 2179–2193. [Google Scholar] [CrossRef]
- Tung, B.T.; Hai, N.T.; Son, P.K. Hepatoprotective effect of phytosome curcumin against paracetamol-induced liver toxicity in mice. Braz. J. Pharm. Sci. 2017, 53. [Google Scholar] [CrossRef]
- Lee, H.Y.; Kim, S.W.; Lee, G.H.; Choi, M.K.; Jung, H.W.; Kim, Y.J.; Kwon, H.J.; Chae, H.J. Turmeric extract and its active compound, curcumin, protect against chronic CCl4-induced liver damage by enhancing antioxidation. BMC Complement. Altern. Med. 2016, 16, 316. [Google Scholar] [CrossRef] [PubMed]
- Bayindir, N.; Esrefoglu, M.; Kumas, M.; Iraz, M.; Kesgin, S.; Kilic, E. Protective effect of curcumin on cadmium-induced liver apoptosis in rats. Bezmialem Sci. 2016, 4, 99–105. [Google Scholar] [CrossRef]
- Afrin, R.; Arumugam, S.; Soetikno, V.; Thandavarayan, R.A.; Pitchaimani, V.; Karuppagounder, V.; Sreedhar, R.; Harima, M.; Suzuki, H.; Miyashita, S.; et al. Curcumin ameliorates streptozotocin-induced liver damage through modulation of endoplasmic reticulum stress-mediated apoptosis in diabetic rats. Free Radic. Res. 2015, 49, 279–289. [Google Scholar] [CrossRef] [PubMed]
- Granados-Castro, L.F.; Rodriguez-Rangel, D.S.; Fernandez-Rojas, B.; Leon-Contreras, J.C.; Hernandez-Pando, R.; Medina-Campos, O.N.; Eugenio-Perez, D.; Pinzon, E.; Pedraza-Chaverri, J. Curcumin prevents paracetamol-induced liver mitochondrial alterations. J. Pharm. Pharmacol. 2016, 68, 245–256. [Google Scholar] [CrossRef] [PubMed]
- Guo, C.; Ma, J.F.; Zhong, Q.H.; Zhao, M.Y.; Hu, T.X.; Chen, T.; Qiu, L.X.; Wen, L.P. Curcumin improves alcoholic fatty liver by inhibiting fatty acid biosynthesis. Toxicol. Appl. Pharmacol. 2017, 328, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.F.; Xu, W.X.; Zhang, F.; Shao, J.J.; Zheng, S.Z. Nrf2 knockdown disrupts the protective effect of curcumin on alcohol-induced hepatocyte necroptosis. Mol. Pharm. 2016, 13, 4043–4053. [Google Scholar] [CrossRef] [PubMed]
- Buzzetti, E.; Pinzani, M.; Tsochatzis, E.A. The multiple-hit pathogenesis of non-alcoholic fatty liver disease (NAFLD). Metab. Clin. Exp. 2016, 65, 1038–1048. [Google Scholar] [CrossRef] [PubMed]
- Zabihi, N.A.; Pirro, M.; Johnston, T.P.; Sahebkar, A. Is there a role for curcumin supplementation in the treatment of non-alcoholic fatty liver disease? The data suggest yes. Curr. Pharm. Des. 2017, 23, 969–982. [Google Scholar] [CrossRef] [PubMed]
- Inzaugarat, M.E.; De Matteo, E.; Baz, P.; Lucero, D.; Garcia, C.C.; Ballerga, E.G.; Daruich, J.; Sorda, J.A.; Wald, M.R.; Chernavsky, A.C. New evidence for the therapeutic potential of curcumin to treat nonalcoholic fatty liver disease in humans. PLoS ONE 2017, 12, e0172900. [Google Scholar] [CrossRef] [PubMed]
- Watanabe, K.; Karuppagounder, V.; Arumugam, S.; Thandavarayan, R.A.; Pitchaimani, V.; Sreedhar, R.; Afrin, R.; Harima, M.; Suzuki, H.; Suzuki, K.; et al. Pruni cortex ameliorates skin inflammation possibly through HMGB1-NF kappa B pathway in house dust mite induced atopic dermatitis NC/NGA transgenic mice. J. Clin. Biochem. Nutr. 2015, 56, 186–194. [Google Scholar] [CrossRef] [PubMed]
- Afrin, R.; Arumugam, S.; Rahman, A.; Wahed, M.I.I.; Karuppagounder, V.; Harima, M.; Suzuki, H.; Miyashita, S.; Suzuki, K.; Yoneyama, H.; et al. Curcumin ameliorates liver damage and progression of nash in NASH-HCC mouse model possibly by modulating HMGB1-Nf-kappa B translocation. Int. Immunopharmacol. 2017, 44, 174–182. [Google Scholar] [CrossRef] [PubMed]
- Kheradpezhouh, E.; Barritt, G.J.; Rychkov, G.Y. Curcumin inhibits activation of TRPM2 channels in rat hepatocytes. Redox Biol. 2016, 7, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Chen, N.Z.; Geng, Q.Q.; Zheng, J.B.; He, S.; Huo, X.W.; Sun, X.J. Suppression of the TGF-beta/Smad signaling pathway and inhibition of hepatic stellate cell proliferation play a role in the hepatoprotective effects of curcumin against alcohol-induced hepatic fibrosis. Int. J. Mol. Med. 2014, 34, 1110–1116. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.L.; Guo, Y.; Zhang, S.; Zhang, Y.; Wang, Y.Q.; Ni, W.X.; Kong, D.S.; Chen, W.J.; Zheng, S.Z. Curcumin modulates cannabinoid receptors in liver fibrosis in vivo and inhibits extracellular matrix expression in hepatic stellate cells by suppressing cannabinoid receptor type-1 in vitro. Eur. J. Pharmacol. 2013, 721, 133–140. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Zhang, Z.L.; Chen, L.; Kong, D.S.; Zhang, X.P.; Lu, C.F.; Lu, Y.; Zheng, S.Z. Curcumin attenuates angiogenesis in liver fibrosis and inhibits angiogenic properties of hepatic stellate cells. J. Cell. Mol. Med. 2014, 18, 1392–1406. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.J.; Wu, C.Z.; Lin, Z.; Guo, Y.; Shi, L.; Dong, P.H.; Lu, Z.Q.; Gao, S.M.; Liao, Y.; Chen, B.C.; et al. Curcumin up-regulates phosphatase and tensin homologue deleted on chromosome 10 through microRNA-mediated control of DNA methylation—A novel mechanism suppressing liver fibrosis. FEBS J. 2014, 281, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.Y.; Yang, M.F.; Cao, M.Z.; Li, D.W.; Yang, X.Y.; Sun, J.Y.; Zhang, Z.Y.; Mao, L.L.; Zhang, S.; Wang, F.Z.; et al. Strategy to suppress oxidative damage-induced neurotoxicity in PC12 cells by curcumin: The role of ROS-mediated DNA damage and the MAPK and Akt pathways. Mol. Neurobiol. 2016, 53, 369–378. [Google Scholar] [CrossRef] [PubMed]
- Szczepanowicz, K.; Jantas, D.; Piotrowski, M.; Staron, J.; Leskiewicz, M.; Regulska, M.; Lason, W.; Warszynski, P. Encapsulation of curcumin in polyelectrolyte nanocapsules and their neuroprotective activity. Nanotechnology 2016, 27, 355101. [Google Scholar] [CrossRef] [PubMed]
- Jin, M.L.; Park, S.Y.; Shen, Q.; Lai, Y.H.; Ou, X.M.; Mao, Z.; Lin, D.X.; Yu, Y.Y.; Zhang, W.Z. Anti-neuroinflammatory effect of curcumin on Pam3CSK4-stimulated microglial cells. Int. J. Mol. Med. 2018, 41, 521–530. [Google Scholar] [CrossRef] [PubMed]
- Kodali, M.; Hattiangady, B.; Shetty, G.A.; Bates, A.; Shuai, B.; Shetty, A.K. Curcumin treatment leads to better cognitive and mood function in a model of Gulf War Illness with enhanced neurogenesis, and alleviation of inflammation and mitochondrial dysfunction in the hippocampus. Brain Behav. Immun. 2018, 69, 499–514. [Google Scholar] [CrossRef] [PubMed]
- Tegenge, M.A.; Rajbhandari, L.; Shrestha, S.; Mithal, A.; Hosmane, S.; Venkatesan, A. Curcumin protects axons from degeneration in the setting of local neuroinflammation. Exp. Neurol. 2014, 253, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.B.; Li, Q.; Zhang, M.T.; Mao-Ying, Q.L.; Hu, L.Y.; Wu, G.C.; Mi, W.L.; Wang, Y.Q. Curcumin ameliorates neuropathic pain by down-regulating spinal IL-1 beta via suppressing astroglial NALP1 inflammasome and JAK2-STAT3 signalling. Sci. Rep. 2016, 6, 28956. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.Y.; Li, Q.; Chang, R.M.; Yang, D.; Song, Z.B.; Guo, Q.L.; Huang, C.S. Curcumin alleviates neuropathic pain by inhibiting p300/CBP histone acetyltransferase activity-regulated expression of BDNF and Cox-2 in a rat model. PLoS ONE 2014, 9, e91303. [Google Scholar] [CrossRef] [PubMed]
- Dong, W.W.; Yang, B.; Wang, L.L.; Li, B.X.; Guo, X.S.; Zhang, M.; Jiang, Z.F.; Fu, J.Q.; Pi, J.B.; Guan, D.W.; et al. Curcumin plays neuroprotective roles against traumatic brain injury partly via nrf2 signaling. Toxicol. Appl. Pharmacol. 2018, 346, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.F.; Chen, C.W.; Zhang, X.; Li, X.; Chen, Z.P.; Yang, C.; Liang, X.L.; Zhu, G.C.; Xu, Z. Neuroprotective effect of curcumin against cerebral ischemia-reperfusion via mediating autophagy and inflammation. J. Mol. Neurosci. 2018, 64, 129–139. [Google Scholar] [CrossRef] [PubMed]
- Hu, S.X.; Maiti, P.; Ma, Q.L.; Zuo, X.H.; Jones, M.R.; Cole, G.M.; Frautschy, S.A. Clinical development of curcumin in neurodegenerative disease. Expert Rev. Neurother. 2015, 15, 629–637. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.J.; Li, Z.H.; Liu, L.; Tang, W.X.; Wang, Y.; Dong, M.R.; Xiao, C. Curcumin attenuates beta-amyloid-induced neuroinflammation via activation of peroxisome proliferator-activated receptor-gamma function in a rat model of Alzheimer’s disease. Front. Pharmacol. 2016, 7, 261. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.L.; Li, J.F.; Wang, Y.T.; Xu, C.Y.; Hua, L.L.; Yang, X.P.; Geng, S.; Wang, S.S.; Wang, Z.; Yin, H.L. Curcumin reduces hippocampal neuron apoptosis and JNK-3 phosphorylation in rats with A beta-induced Alzheimer’s disease: Protecting spatial learning and memory. J. Neurorestoratol. 2017, 5, 117–123. [Google Scholar] [CrossRef]
- Huang, H.C.; Zheng, B.W.; Guo, Y.; Zhao, J.; Zhao, J.Y.; Ma, X.W.; Jiang, Z.F. Antioxidative and neuroprotective effects of curcumin in an Alzheimer’s disease rat model co-treated with intracerebroventricular streptozotocin and subcutaneous D-galactose. J. Alzheimers Dis. 2016, 52, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Jiang, T.F.; Zhang, Y.J.; Zhou, H.Y.; Wang, H.M.; Tian, L.P.; Liu, J.; Ding, J.Q.; Chen, S.D. Curcumin ameliorates the neurodegenerative pathology in A53T alpha-synuclein cell model of Parkinson’s disease through the downregulation of mTOR/p70s6K signaling and the recovery of macroautophagy. J. Neuroimmune Pharmacol. 2013, 8, 356–369. [Google Scholar] [CrossRef] [PubMed]
- Sharma, N.; Nehru, B. Curcumin affords neuroprotection and inhibits alpha-synuclein aggregation in lipopolysaccharide-induced Parkinson’s disease model. Inflammopharmacology 2018, 26, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Khatri, D.K.; Jnvekar, A.R. Neuroprotective effect of curcumin as evinced by abrogation of rotenone-induced motor deficits, oxidative and mitochondrial dysfunctions in mouse model of Parkinson’s disease. Pharmacol. Biochem. Behav. 2016, 150, 39–47. [Google Scholar] [CrossRef] [PubMed]
- Meesarapee, B.; Thampithak, A.; Jaisin, Y.; Sanvarinda, P.; Suksamrarn, A.; Tuchinda, P.; Morales, N.P.; Sanvarinda, Y. Curcumin I mediates neuroprotective effect through attenuation of quinoprotein formation, p-p38 MAPK expression, and caspase-3 activation in 6-hydroxydopamine treated SH-SY5Y cells. Phytother. Res. 2014, 28, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Abbaoui, A.; Chatoui, H.; El Hiba, O.; Gamrani, H. Neuroprotective effect of curcumin-i in copper-induced dopaminergic neurotoxicity in rats: A possible link with Parkinson’s disease. Neurosci. Lett. 2017, 660, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, P.; Yadav, R.S.; Chandravanshi, L.P.; Shukla, R.K.; Dhuriya, Y.K.; Chauhan, L.K.S.; Dwivedi, H.N.; Pant, A.B.; Khanna, V.K. Unraveling the mechanism of neuroprotection of curcumin in arsenic induced cholinergic dysfunctions in rats. Toxicol. Appl. Pharmacol. 2014, 279, 428–440. [Google Scholar] [CrossRef] [PubMed]
- Agthong, S.; Kaewsema, A.; Charoensub, T. Curcumin ameliorates functional and structural abnormalities in cisplatin-induced neuropathy. Exp. Neurobiol. 2015, 24, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.L.; Li, H.C.; Zhao, W.; Peng, H.H.; Huang, F.; Jiang, W.H.; Xu, S.Y. Curcumin attenuated bupivacaine-induced neurotoxicity in SH-SY5Y cells via activation of the Akt signaling pathway. Neurochem. Res. 2016, 41, 2425–2432. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Tian, S.Q.; Yang, X.; Liu, J.J.; Wang, Y.H.; Sun, K. Celecoxib-induced inhibition of neurogenesis in fetal frontal cortex is attenuated by curcumin via Wnt/beta-catenin pathway. Life Sci. 2017, 185, 95–102. [Google Scholar] [CrossRef] [PubMed]
- Carmona-Ramirez, I.; Santamaria, A.; Tobon-Velasco, J.C.; Orozco-Ibarra, M.; Gonzalez-Herrera, I.G.; Pedraza-Chaverri, J.; Maldonado, P.D. Curcumin restores Nrf2 levels and prevents quinolinic acid-induced neurotoxicity. J. Nutr. Biochem. 2013, 24, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, M.; Wicz, S.; Corral, R.S. Cardioprotective actions of curcumin on the pathogenic NFAT/COX-2/prostaglandin E-2 pathway induced during Trypanosoma cruzi infection. Phytomedicine 2016, 23, 1392–1400. [Google Scholar] [CrossRef] [PubMed]
- Yu, W.; Zha, W.L.; Ke, Z.Q.; Min, Q.; Li, C.R.; Sun, H.R.; Liu, C. Curcumin protects neonatal rat cardiomyocytes against high glucose-induced apoptosis via PI3K/Akt signalling pathway. J. Diabetes Res. 2016, 2016, 4158591. [Google Scholar] [CrossRef] [PubMed]
- Kohli, S.; Chhabra, A.; Jaiswal, A.; Rustagi, Y.; Sharma, M.; Rani, V. Curcumin suppresses gelatinase B mediated norepinephrine induced stress in H9c2 cardiomyocytes. PLoS ONE 2013, 8, e76519. [Google Scholar] [CrossRef] [PubMed]
- Manghani, C.; Gupta, A.; Tripathi, V.; Rani, V. Cardioprotective potential of curcumin against norepinephrine-induced cell death: A microscopic study. J. Microsc. 2017, 265, 232–244. [Google Scholar] [CrossRef] [PubMed]
- Naserzadeh, P.; Mehr, S.N.; Sadabadi, Z.; Seydi, E.; Salimi, A.; Pourahmad, J. Curcumin protects mitochondria and cardiomyocytes from oxidative damage and apoptosis induced by hemiscorpius lepturus venom. Drug Res. 2018, 68, 113–120. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.B.; Jiang, H.; Shi, Y. Upregulation of heme oxygenase-1 expression by curcumin conferring protection from hydrogen peroxide-induced apoptosis in H9c2 cardiomyoblasts. Cell Biosci. 2017, 7, 523. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.J.; Hao, J.T.; Wang, J.; Zhang, W.F.; Yan, C.P.; Zhao, J.H. Curcumin inhibits cardiac hypertrophy and improves cardiovascular function via enhanced Na+/Ca2+ exchanger expression after transverse abdominal aortic constriction in rats. Pharmacol. Rep. 2018, 70, 60–68. [Google Scholar] [CrossRef] [PubMed]
- Chen, R.C.; Peng, X.F.; Du, W.M.; Wu, Y.; Huang, B.; Xue, L.; Wu, Q.; Qiu, H.M.; Jiang, Q.S. Curcumin attenuates cardiomyocyte hypertrophy induced by high glucose and insulin via the PPAR gamma/Akt/NO signaling pathway. Diabetes Res. Clin. Pract. 2015, 108, 235–242. [Google Scholar] [CrossRef] [PubMed]
- Katanasaka, Y.; Sunagawa, Y.; Hasegawa, K.; Morimoto, T. Application of curcumin to heart failure therapy by targeting transcriptional pathway in cardiomyocytes. Biol. Pharm. Bull. 2013, 36, 13–17. [Google Scholar] [CrossRef] [PubMed]
- Cao, Q.; Zhang, J.X.; Gao, L.; Zhang, Y.J.; Dai, M.Y.; Bao, M.W. Dickkopf-3 upregulation mediates the cardioprotective effects of curcumin on chronic heart failure. Mol. Med. Rep. 2018, 17, 7249–7257. [Google Scholar] [CrossRef] [PubMed]
- Feng, D.; Zou, J.; Zhang, S.S.; Li, X.C.; Lui, M.Q. Hypocholesterolemic activity of curcumin is mediated by down-regulating the expression of Niemann-Pick C1-like 1 in hamsters. J. Agric. Food Chem. 2017, 65, 276–280. [Google Scholar] [CrossRef] [PubMed]
- Meng, B.; Li, J.; Cao, H. Antioxidant and antiinflammatory activities of curcumin on diabetes mellitus and its complications. Curr. Pharm. Des. 2013, 19, 2101–2113. [Google Scholar] [PubMed]
- Chuengsamarn, S.; Rattanamongkolgul, S.; Luechapudiporn, R.; Phisalaphong, C.; Jirawatnotai, S. Curcumin extract for prevention of type 2 diabetes. Diabetes Care 2012, 35, 2121–2127. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, P.P.; Ying, J.; Chen, Z.; Yu, S.P. Curcumin attenuates retinal vascular leakage by inhibiting calcium/calmodulin-dependent protein kinase ii activity in streptozotocin-induced diabetes. Cell. Physiol. Biochem. 2016, 39, 1196–1208. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, P.P.; Zhu, Y.X.; Chen, Z.; Shi, T.Y.; Lei, W.S.; Yu, S.P. Curcumin inhibits neuronal loss in the retina and elevates Ca2+/calmodulin-dependent protein kinase II activity in diabetic rats. J. Ocul. Pharmacol. Ther. 2015, 31, 555–562. [Google Scholar] [CrossRef] [PubMed]
- Wojcik, M.; Krawczyk, M.; Wojcik, P.; Cypryk, K.; Wozniak, L.A. Molecular mechanisms underlying curcumin-mediated therapeutic effects in type 2 diabetes and cancer. Oxid. Med. Cell. Longev. 2018, 2018, 9698258. [Google Scholar] [CrossRef] [PubMed]
- Jhong, C.-H.; Riyaphan, J.; Lin, S.-H.; Chia, Y.-C.; Weng, C.-F. Screening alpha-glucosidase and alpha-amylase inhibitors from natural compounds by molecular docking in silico. BioFactors 2015, 41, 242–251. [Google Scholar] [CrossRef] [PubMed]
- Akolade, J.O.; Oloyede, H.O.B.; Onyenekwe, P.C. Encapsulation in chitosan-based polyelectrolyte complexes enhances antidiabetic activity of curcumin. J. Funct. Foods 2017, 35, 584–594. [Google Scholar] [CrossRef]
- Gutierres, V.O.; Campos, M.L.; Arcaro, C.A.; Assis, R.P.; Baldan-Cimatti, H.M.; Peccinini, R.G.; Paula-Gomes, S.; Kettelhut, I.C.; Baviera, A.M.; Brunetti, I.L. Curcumin pharmacokinetic and pharmacodynamic evidences in streptozotocin-diabetic rats support the antidiabetic activity to be via metabolite(s). Evid. Based Complement. Alternat. Med. 2015, 2015, 678218. [Google Scholar] [CrossRef] [PubMed]
- Chanpoo, M.; Petchpiboonthai, H.; Panyarachun, B.; Anupunpisit, V. Effect of curcumin in the amelioration of pancreatic islets in streptozotocin-induced diabetic mice. J. Med. Assoc. Thai. 2010, 93 (Suppl. 6), S152–S159. [Google Scholar]
- Ou, J.L.; Mizushina, Y.; Wang, S.Y.; Chuang, D.Y.; Nadar, M.; Hsu, W.L. Structure-activity relationship analysis of curcumin analogues on anti-influenza virus activity. FEBS J. 2013, 280, 5829–5840. [Google Scholar] [CrossRef] [PubMed]
- Abdollahi, M.F.; Zandi, M.; Shokrollahi, P.; Ehsani, M. Synthesis and characterization of curcumin segmented polyurethane with induced antiplatelet activity. J. Polym. Res. 2015, 22, 179. [Google Scholar] [CrossRef]
- Haghighizad, H.; Touhidi, A.; Pourmotabbed, A.; Moradpour, F.; Nedaei, S.E.; Pourmotabbed, T. Curcumin improves chronic stress induced potentiated seizure activity in experimental model of epilepsy. J. Neurol. Sci. Turk. 2017, 34, 76–85. [Google Scholar]
- Mantzorou, M.; Pavlidou, E.; Vasios, G.; Tsagalioti, E.; Giaginis, C. Effects of curcumin consumption on human chronic diseases: A narrative review of the most recent clinical data. Phytother. Res. 2018, 32, 957–975. [Google Scholar] [CrossRef] [PubMed]
- Bilia, A.R.; Bergonzi, M.C.; Isacchi, B.; Antiga, E.; Caproni, M. Curcumin nanoparticles potentiate therapeutic effectiveness of acitrein in moderate-to-severe psoriasis patients and control serum cholesterol levels. J. Pharm. Pharmacol. 2018, 70, 919–928. [Google Scholar] [CrossRef] [PubMed]
- Greil, R.; Greil-Ressler, S.; Weiss, L.; Schonlieb, C.; Magnes, T.; Radl, B.; Bolger, G.T.; Vcelar, B.; Sordillo, P.P. A phase 1 dose-escalation study on the safety, tolerability and activity of liposomal curcumin (lipocurc((TM))) in patients with locally advanced or metastatic cancer. Cancer Chemother. Pharmacol. 2018, 82, 695–706. [Google Scholar] [CrossRef] [PubMed]
- Jimenez-Osorio, A.S.; Garcia-Nino, W.R.; Gonzalez-Reyes, S.; Alvarez-Mejia, A.E.; Guerra-Leon, S.; Salazar-Segovia, J.; Falcon, I.; de Oca-Solano, H.M.; Madero, M.; Pedraza-Chaverri, J. The effect of dietary supplementation with curcumin on redox status and Nrf2 activation in patients with nondiabetic or diabetic proteinuric chronic kidney disease: A pilot study. J. Renal Nutr. 2016, 26, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Amalraj, A.; Varma, K.; Jacob, J.; Divya, C.; Kunnumakkara, A.B.; Stohs, S.J.; Gopi, S. A novel highly bioavailable curcumin formulation improves symptoms and diagnostic indicators in rheumatoid arthritis patients: A randomized, double-blind, placebo-controlled, two-dose, three-arm, and parallel-group study. J. Med. Food 2017, 20, 1022–1030. [Google Scholar] [CrossRef] [PubMed]
- Henrotin, Y.; Gharbi, M.; Dierckxsens, Y.; Priem, F.; Marty, M.; Seidel, L.; Albert, A.; Heuse, E.; Bonnet, V.; Castermans, C. Decrease of a specific biomarker of collagen degradation in osteoarthritis, Coll2-1, by treatment with highly bioavailable curcumin during an exploratory clinical trial. BMC Complement. Altern. Med. 2014, 14, 159. [Google Scholar] [CrossRef] [PubMed]
- Antiga, E.; Bonciolini, V.; Volpi, W.; Del Bianco, E.; Caproni, M. Oral curcumin (Meriva) is effective as an adjuvant treatment and is able to reduce IL-22 serum levels in patients with psoriasis vulgaris. BioMed Res. Int. 2015, 2015, 283634. [Google Scholar] [CrossRef] [PubMed]
- Abdolahi, M.; Sarraf, P.; Javanbakht, M.H.; Honarvar, N.M.; Hatami, M.; Soveyd, N.; Tafakhori, A.; Sedighiyan, M.; Djalali, M.; Jafarieh, A.; et al. A novel combination of omega-3 fatty acids and nano-curcumin modulates interleukin-6 gene expression and high sensitivity c-reactive protein serum levels in patients with migraine: A randomized clinical trial study. CNS Neurol. Disord. Drug Targets 2018, 17, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Kuriakose, M.A.; Ramdas, K.; Dey, B.; Iyer, S.; Rajan, G.; Elango, K.K.; Suresh, A.; Ravindran, D.; Kumar, R.R.; Prathiba, R.; et al. A randomized double-blind placebo-controlled phase iib trial of curcumin in oral leukoplakia. Cancer Prev. Res. 2016, 9, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Duetzmann, S.; Schiborr, C.; Kocher, A.; Pilatus, U.; Hattingen, E.; Weissenberger, J.; Gessler, F.; Quick-Weller, J.; Franz, K.; Seifert, V.; et al. Intratumoral concentrations and effects of orally administered micellar curcuminoids in glioblastoma patients. Nutr. Cancer 2016, 68, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Funamoto, M.; Sunagawa, Y.; Katanasaka, Y.; Miyazaki, Y.; Imaizumi, A.; Kakeya, H.; Yamakage, H.; Satoh-Asahara, N.; Komiyama, M.; Wada, H.; et al. Highly absorptive curcumin reduces serum atherosclerotic low-density lipoprotein levels in patients with mild copd. Int. J. Chron. Obstruct. Pulmon. Dis. 2016, 11, 2029–2034. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.S.; Su, Y.F.; Yang, H.W.; Lee, Y.H.; Chou, J.I.; Ueng, K.C. Lipid-lowering effects of curcumin in patients with metabolic syndrome: A randomized, double-blind, placebo-controlled trial. Phytother. Res. 2014, 28, 1770–1777. [Google Scholar] [CrossRef] [PubMed]
- Chuengsamarn, S.; Rattanamongkolgul, S.; Phonrat, B.; Tungtrongchitr, R.; Jirawatnotai, S. Reduction of atherogenic risk in patients with type 2 diabetes by curcuminoid extract: A randomized controlled trial. J. Nutr. Biochem. 2014, 25, 144–150. [Google Scholar] [CrossRef] [PubMed]
- Mirzabeigi, P.; Mohammadpour, A.H.; Salarifar, M.; Gholami, K.; Mojtahedzadeh, M.; Javadi, M.R. The effect of curcumin on some of traditional and non-traditional cardiovascular risk factors: A pilot randomized, double-blind, placebo-controlled trial. Iran. J. Pharm. Res. 2015, 14, 479–486. [Google Scholar] [PubMed]
- Neerati, P.; Devde, R.; Gangi, A.K. Evaluation of the effect of curcumin capsules on glyburide therapy in patients with type-2 diabetes mellitus. Phytother. Res. 2014, 28, 1796–1800. [Google Scholar] [CrossRef] [PubMed]
- Rahimi, H.R.; Mohammadpour, A.H.; Dastani, M.; Jaafari, M.R.; Abnous, K.; Mobarhan, M.G.; Oskuee, R.K. The effect of nano-curcumin on HBA1C, fasting blood glucose, and lipid profile in diabetic subjects: A randomized clinical trial. Avicenna J. Phytomed. 2016, 6, 567–577. [Google Scholar] [PubMed]
- Kunati, S.R.; Yang, S.M.; William, B.M.; Xu, Y. An LC-MS/MS method for simultaneous determination of curcumin, curcumin glucuronide and curcumin sulfate in a phase II clinical trial. J. Pharm. Biomed. Anal. 2018, 156, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Adahoun, M.A.; Al-Akhras, M.A.H.; Jaafar, M.S.; Bououdina, M. Enhanced anti-cancer and antimicrobial activities of curcumin nanoparticles. Artif. Cells Nanomed. Biotechnol. 2017, 45, 98–107. [Google Scholar] [CrossRef] [PubMed]
- Bai, F.; Diao, J.J.; Wang, Y.; Sun, S.X.; Zhang, H.M.; Liu, Y.Y.; Wang, Y.Q.; Cao, J. A new water-soluble nanomicelle formed through self-assembly of pectin-curcumin conjugates: Preparation, characterization, and anticancer activity evaluation. J. Agric. Food Chem. 2017, 65, 6840–6847. [Google Scholar] [CrossRef] [PubMed]
- Khayyal, M.T.; El-Hazek, R.M.; El-Sabbagh, W.A.; Frank, J.; Behnam, D.; Abdel-Tawab, M. Micellar solubilisation enhances the antiinflammatory activities of curcumin and boswellic acids in rats with adjuvant-induced arthritis. Nutrition 2018, 54, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Akbar, M.U.; Zia, K.M.; Nazir, A.; Iqbal, J.; Ejaz, S.A.; Akash, M.S.H. Pluronic-based mixed polymeric micelles enhance the therapeutic potential of curcumin. AAPS PharmSciTech 2018, 19, 2719–2739. [Google Scholar] [CrossRef] [PubMed]
- Schiborr, C.; Kocher, A.; Behnam, D.; Jandasek, J.; Toelstede, S.; Frank, J. The oral bioavailability of curcumin from micronized powder and liquid micelles is significantly increased in healthy humans and differs between sexes. Mol. Nutr. Food Res. 2014, 58, 516–527. [Google Scholar] [CrossRef] [PubMed]
- Kocher, A.; Bohnert, L.; Schiborr, C.; Frank, J. Highly bioavailable micellar curcuminoids accumulate in blood, are safe and do not reduce blood lipids and inflammation markers in moderately hyperlipidemic individuals. Mol. Nutr. Food Res. 2016, 60, 1555–1563. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, T.; Xu, L.; Zhang, Z.; Li, L.; Chen, H. Preparation of curcumin micelles and the in vitro and in vivo evaluation for cancer therapy. J. Biomed. Nanotechnol. 2014, 10, 1458–1468. [Google Scholar] [CrossRef] [PubMed]
- Hagl, S.; Kocher, A.; Schiborr, C.; Kolesova, N.; Frank, J.; Eckert, G.P. Curcumin micelles improve mitochondrial function in neuronal PC12 cells and brains of nmri mice–impact on bioavailability. Neurochem. Int. 2015, 89, 234–242. [Google Scholar] [CrossRef] [PubMed]
- Baker, M. Deceptive curcumin offers cautionary tale for chemists. Nature 2017, 541, 144–145. [Google Scholar] [CrossRef] [PubMed]
- Bahadori, F.; Demiray, M. A realistic view on “the essential medicinal chemistry of curcumin”. ACS Med. Chem. Lett. 2017, 8, 893–896. [Google Scholar] [CrossRef] [PubMed]
- Heger, M. Drug screening: Don’t discount all curcumin trial data. Nature 2017, 543, 40. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.R.; Song, Z.L.; Weng, J.P.; Fantus, I.G. Curcumin and other dietary polyphenols: Potential mechanisms of metabolic actions and therapy for diabetes and obesity. Am. J. Physiol. Endocrinol. Metab. 2018, 314, E201–E205. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Type | Subjects | Dose | Potential Mechanisms | Ref. |
|---|---|---|---|---|
| Effects on cancer cell growth and proliferation | ||||
| In vitro | HNSCC cells | 12.5 μM | Upregulating pro-apoptotic Bik Downregulating survival signaling by Akt and NF-κB Reducing STAT3 expression Suppressing cyclin D1 and cyclin D2 expression | [73] |
| In vitro | NPC cells | 5, 10, and 15 μM | Upregulating miR-7 expression Inhibiting Skp2 expression Increasing the G2/M phase fraction | [72] |
| In vivo | CaSki-implanted nude mice | 1000 and 1500 mg/kg b.w. | Downregulating VEGF, COX-2, and EGFR expression Reducing increased capillary networks Attenuating abnormalities of the capillary network pattern | [74] |
| In vitro | MCF-7 breast cancer cells | 2.5 μM | Increasing Bcl-2 expression Decreasing Bax expression Inhibiting EGRF expression | [75] |
| In vitro | Patu8988 pancreatic cancer cells | 10, 15, and 20 μM | Downregulating YAP and TAZ expression Inhibiting Notch-1 expression Inducing arrest in the G2/M phase | [77] |
| In vivo | Albino rats with oral carcinogenesis | 30 and 100 mg/kg b.w. | Decreasing the expression of PCNA, Bcl-2, SOCS1, and STAT3 Diminishing the expression of genes associated with epithelial-mesenchymal transition (EMT) | [79] |
| In vitro | MDA-MB-231 triple negative breast cancer cells | 40 μM | Activating p38-MAPK Reducing the level of CDK2, CDK4, cyclin D1, and cyclin E Inducing cell cycle arrest in the G1/S and G2/M phase | [81] |
| In vitro | Ras-activated HAG-1 human adenocarcinoma cells | 25 μM | Enhancing ERK1/2 Inhibiting Akt, mTOR, and S6K1 expression Inducing arrest in the G2/M phase | [83] |
| In vivo | Male Sprague–Dawley rats | 50 mg/kg b.w. | Co-treatment with diclofenac Inhibiting the telomerase activity Upregulating the tumor suppressor proteins, p51, Rb, and p21 Inducing cell cycle arrest | [84] |
| In vitro | Lung epithelium cancer A549 cells | 5 and 10 μM | Co-treatment with apigen inIncreasing p53 expression Blocking cell cycle progression in the G2/M phase | [85] |
| Effects on tumor cell apoptosis | ||||
| In vitro | Src-activated HAG-1 human adenocarcinoma cells | 25 μM | Suppressing Bcl-xL expression Enhancing Bax expression | [83] |
| In vitro | MCF-7 breast cancer cells | 50 μg/mL | Reducing Mcl-1 gene expression Declining the viability of cells | [91] |
| In vitro | MDAH 2774, SKOV3 and PA1 human ovarian cancer cells | 15 μM | Suppressing SERCA activity Disrupting Ca2+ homeostasis | [92] |
| In vitro | KB human oral epidermoid carcinoma cells | 5 and 12.5 μM | Inhibiting the activity of ZAKα | [93] |
| In vivo & in vitro | DU145 human prostate cancer cells and B16 murine melanoma cells | 5 μM | Curcumin analog EF24 Inhibiting miR-21 expression Enhances PTEN and PDCD4 expression Suppressing cyclin D1 and Ki67 expression | [95] |
| Male NOD scid γ mice (NSG) mice | 200 μg/kg b.w. | |||
| In vitro | MDA-MB-231 metastatic breast and A549 lung cancer cells | 10, 20 and 30 μM | Reducing the expressions of HIF 1-α and nuclear p65 (Rel A) | [97] |
| In vitro | A549 lung cancer cells | 40 μM | Suppressing miR-21 expression Elevating the protein level of PTEN | [96] |
| In vitro | DLD-1, LoVo, HCT116 human colon cancer cells | 12.5 µM | Co-treatment with silymarin Induced five-fold higher caspase 3/7 activity | [76] |
| In vitro | 253J-Bv and T24 human bladder cancer cells | 10 μM | Co-treatment with cisplatin Triggering ROS production Activating caspase 3 Upregulating p-ERK1/2 signaling | [99] |
| In vitro | Rh30 and Rh41 human alveolar rhabdomyosarcoma-derived cells | 10, 25, and 50 μM | Blocking the NF-κB pathway Increasing sensitivity to ionizing radiation | [100] |
| In vitro | HCT116 human colon cancer cells | 5 μM | Downregulating NF-κB activation and regulated gene products Potentiating the chemotherapy of 5-fluorouracil | [101] |
| In vivo & in vitro | HCT116 and HT29 human colon cancer cells Male BALB/c-nu/nu mice | 10, 20, 30, and 40 μM 40 mg/kg b.w. | Downregulating NF-κB activation Inhibiting AMPK/ULK1-dependent autophagy Potentiating 5-fluorouracil-induced reduction in cells’ proliferation and invasion | [102] |
| Effects on metastasis and invasion | ||||
| In vitro | NPC cells | 5, 10, and 15 μM | Inhibiting cell motility Suppressing invasion into the Matrigel-coated membrane | [72] |
| In vitro | Lewis lung cancer cells | 20 μM | Reducing the capacity to invade through Matrigel | [105] |
| Study Type | Subjects | Dose | Potential Mechanisms | Ref. |
|---|---|---|---|---|
| Liver injuries induced by pollutants, drugs, and alcohol | ||||
| In vivo | Swiss albino rats with CCl4 hepatotoxicity | 8.98 μM | Maintaining cellular ROS levels Increasing the level of GR and GST Decreasing the level of NADH oxidase Increasing the activity of SDH | [112] |
| In vivo | Sprague-Dawley rats with CCl4 hepatotoxicity | 200 mg/kg b.w. | Decreasing the activities of AST and ALT and the level of lipid peroxidase Increasing hepatic GSH content | [114] |
| In vivo | Sprague-Dawley rats with diabetes induced by streptozotocin | 100 mg/kg b.w. | Decreasing hepatic endoplasmic reticulum stress marker protein and the sub-arm of unfolded protein response signaling protein Inhibiting TNF-α, IL1β, phospho-p38 MAPK, and ASK1 in liver tissues | [116] |
| In vivo | CD1 mice with paracetamol hepatotoxicity | 35, 50, and 100 mg/kg b.w. | Attenuating the decrease in oxygen consumption in the membrane potential, ATP synthesis, aconitase activity, and activity of respiratory complexes, I, III, and IV | [117] |
| In vivo | Kunming mice with alcoholic fatty liver | 60 mg/kg b.w. | Suppressing ethanol-induced pathways, including biosynthesis of unsaturated fatty acids, fatty acid biosynthesis, and pentose and glucuronate interconversions Inhibited glyoxylate, dicarboxylate, and pyruvate metabolism | [118] |
| In vivo | Male ICR mice with alcoholic fatty liver | 20 μM | Attenuating hepatocyte necroptosis Increasing hepatic Nrf2 expression | [119] |
| Nonalcoholic fatty liver disease | ||||
| In vivo | Peripheral blood mononuclear cells | 10 μM | Reducing cytoplasmic translocation of HMGB1, protein expression of TLR4, and nuclear translocation of NF-κB Suppressing glypican-3 expression, VEGF, and pro-thrombin in NASH liver | [124] |
| C57BL/6J mice with NASH-hepatocellular carcinoma | 100 mg/kg b.w. | |||
| In vivo & in vitro | C57BL/6J mice with NAFLD | 2 g curcumin/kg of diet | Preventing high-fat diet-induced liver injury, metabolic alterations, and intrahepatic CD4+ cell accumulation Reducing the pro-inflammatory and pro-oxidant effects on liver macrophages. | [122] |
| In vivo | TRPM2 knockout Hooded Wistar rats | 5 μM | Inhibiting the activation of TRPM2 channels Restoring Ca2+ levels Reducing oxidative stress Lowering the risk of NASH | [124] |
| Liver fibrosis | ||||
| In vivo | Sprague-Dawley rats with alcohol-induced hepatic fibrosis | 50 μM | Inhibiting HSCs proliferation Stimulating endoplasmic reticulum stress Suppressing the TGF-β/Smad signaling pathway Reducing the viability of HSCs | [125] |
| In vivo & in vitro | Sprague-Dawley rats with CCl4-induced hepatic fibrosis | 100, 200, and 400 mg/kg b.w. | Reducing extracellular matrix overproduction in HSCs Downregulating CBR type 1 Upregulating CBR type 2 | [127] |
| HSCs isolated from rats | 10, 20, and 30 μM | |||
| In vivo & in vitro | Sprague-Dawley rats with CCl4-induced hepatic fibrosis | 100, 200, and 400 mg/kg b.w. | Disrupted PDGF-R/ERK and mTOR pathways Activating PPAR-γ Reducing the angiogenic properties of HSCs | [128] |
| HSCs isolated from rats | 20 μM | |||
| In vivo & in vitro | Sprague Dawley rats with CCl4-induced hepatic fibrosis | 200 mg/kg b.w. | Upregulating miR-29b expression Downregulating DNA methyltransferase 3b Upregulating PTEN Inhibiting activated HSCs | [129] |
| Rat HSC-T6 cells | 20 μM | |||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, X.-Y.; Meng, X.; Li, S.; Gan, R.-Y.; Li, Y.; Li, H.-B. Bioactivity, Health Benefits, and Related Molecular Mechanisms of Curcumin: Current Progress, Challenges, and Perspectives. Nutrients 2018, 10, 1553. https://doi.org/10.3390/nu10101553
Xu X-Y, Meng X, Li S, Gan R-Y, Li Y, Li H-B. Bioactivity, Health Benefits, and Related Molecular Mechanisms of Curcumin: Current Progress, Challenges, and Perspectives. Nutrients. 2018; 10(10):1553. https://doi.org/10.3390/nu10101553
Chicago/Turabian StyleXu, Xiao-Yu, Xiao Meng, Sha Li, Ren-You Gan, Ya Li, and Hua-Bin Li. 2018. "Bioactivity, Health Benefits, and Related Molecular Mechanisms of Curcumin: Current Progress, Challenges, and Perspectives" Nutrients 10, no. 10: 1553. https://doi.org/10.3390/nu10101553
APA StyleXu, X.-Y., Meng, X., Li, S., Gan, R.-Y., Li, Y., & Li, H.-B. (2018). Bioactivity, Health Benefits, and Related Molecular Mechanisms of Curcumin: Current Progress, Challenges, and Perspectives. Nutrients, 10(10), 1553. https://doi.org/10.3390/nu10101553