Maternal Protein Restriction Alters the Renal Ptger1 DNA Methylation State in SHRSP Offspring

 ,
, 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement and Animal Conditions
2.2. Animal Experiment 1: Water-Drinking Offspring
2.3. Animal Experiment 2: Salt-Loading Offspring
2.4. Blood Pressure Measurement
2.5. Biochemical Analysis
2.6. Quantitative Real-Time PCR
2.7. Analysis of DNA Methylation Profile
2.8. Statistical Analysis
3. Results
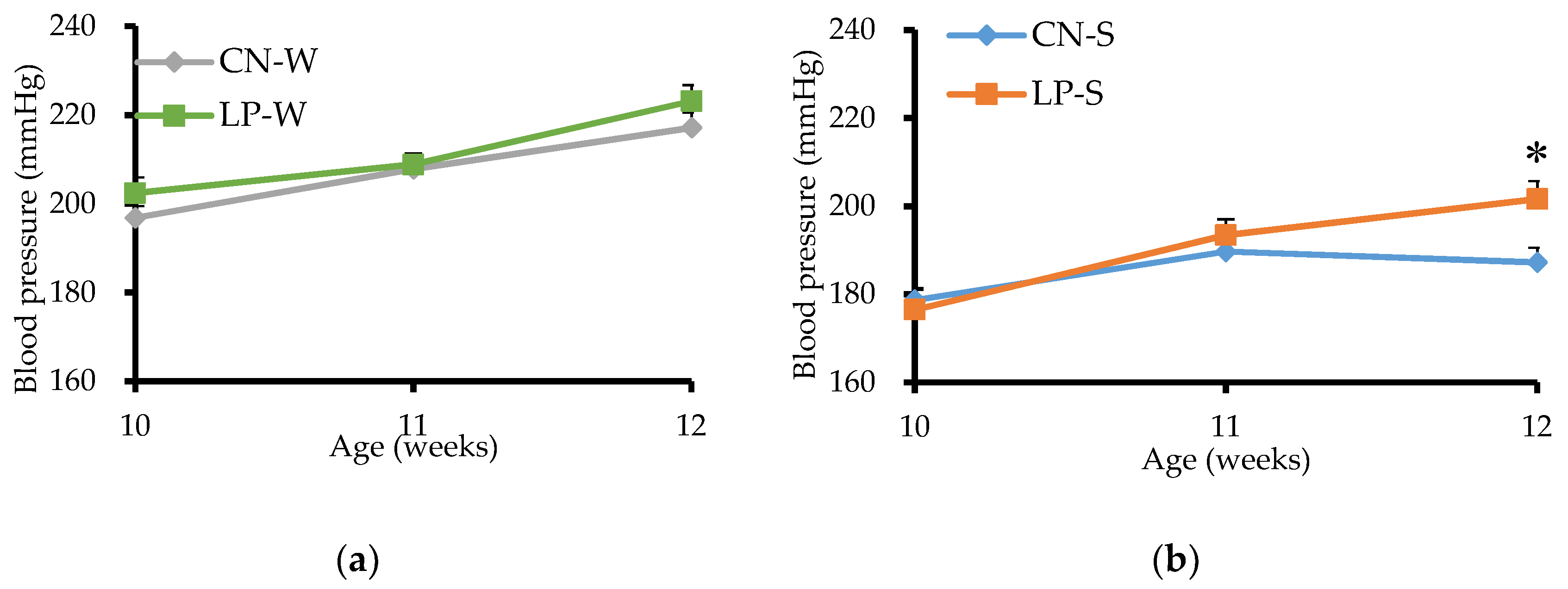
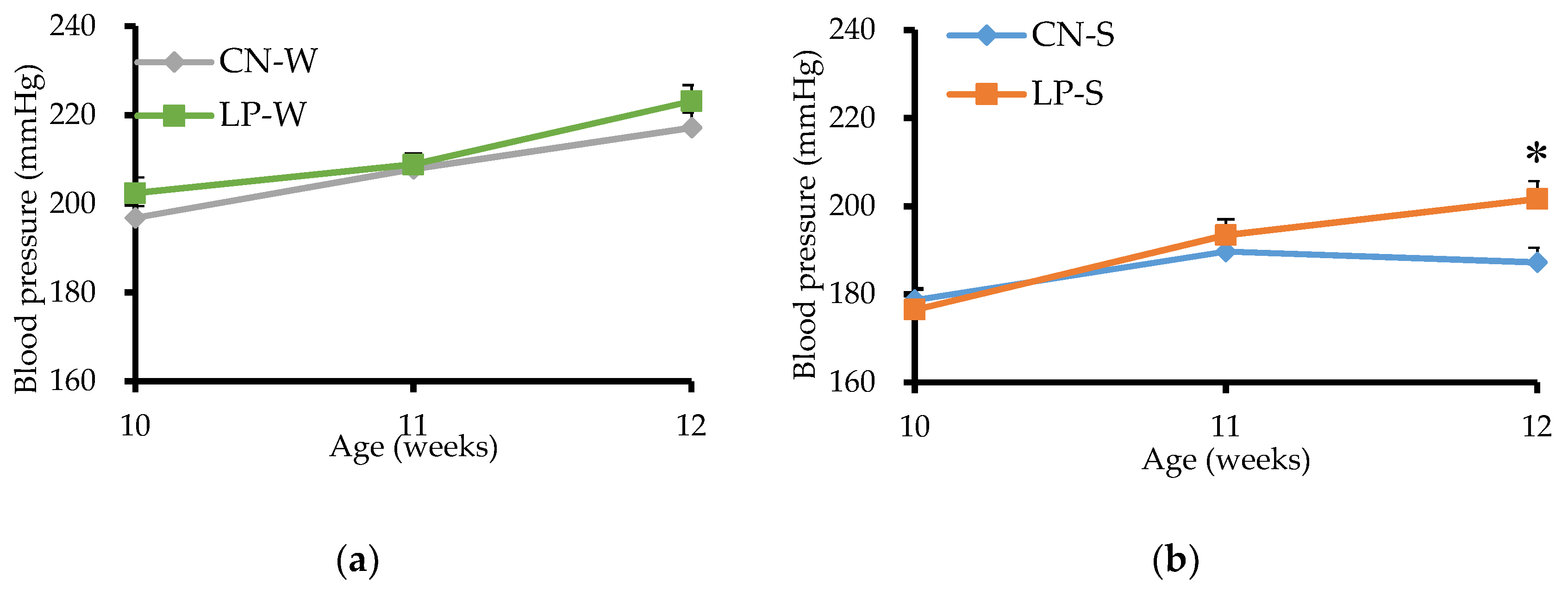
3.1. Blood Pressure of Offspring from 10 to 12 Weeks after Birth
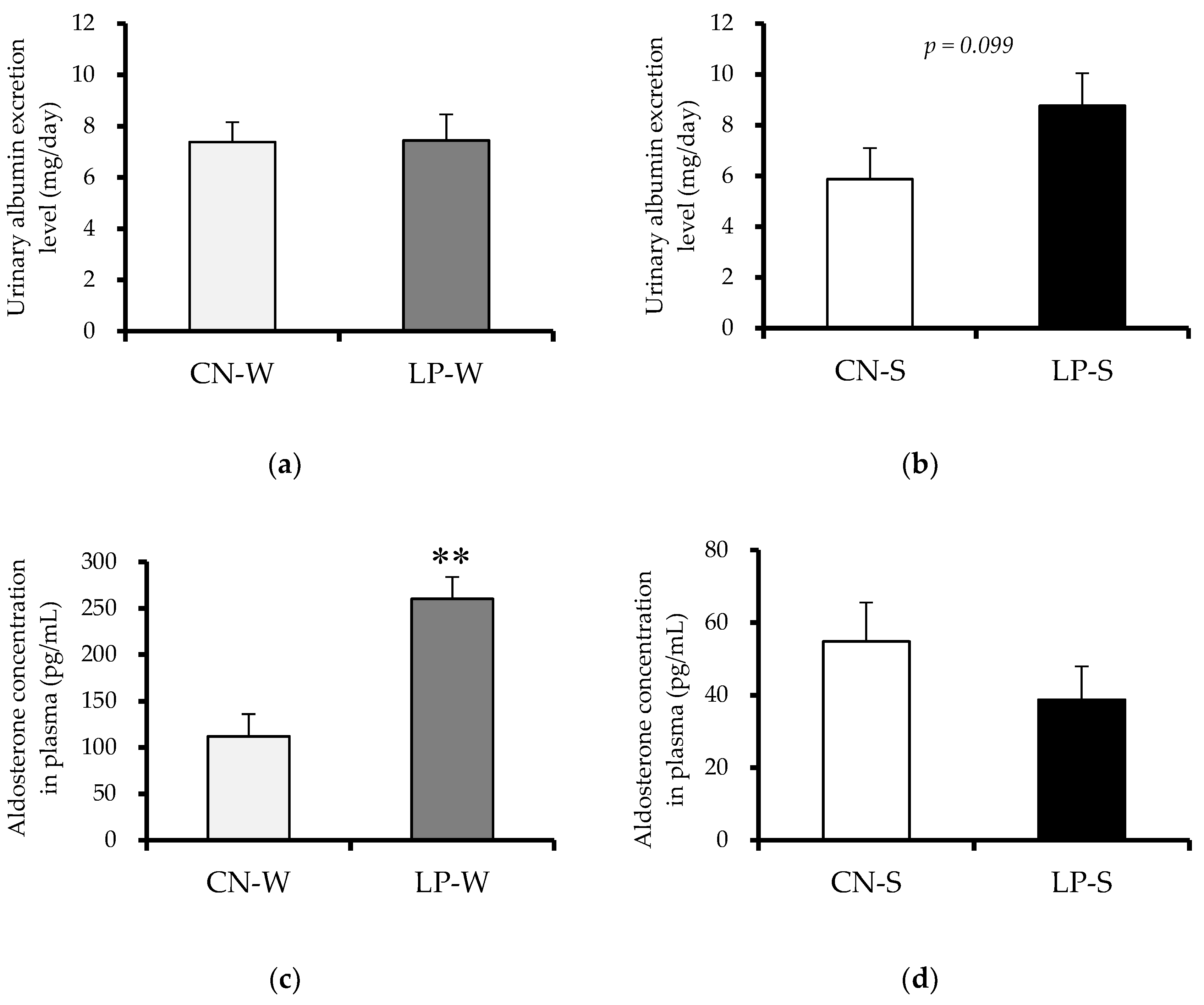
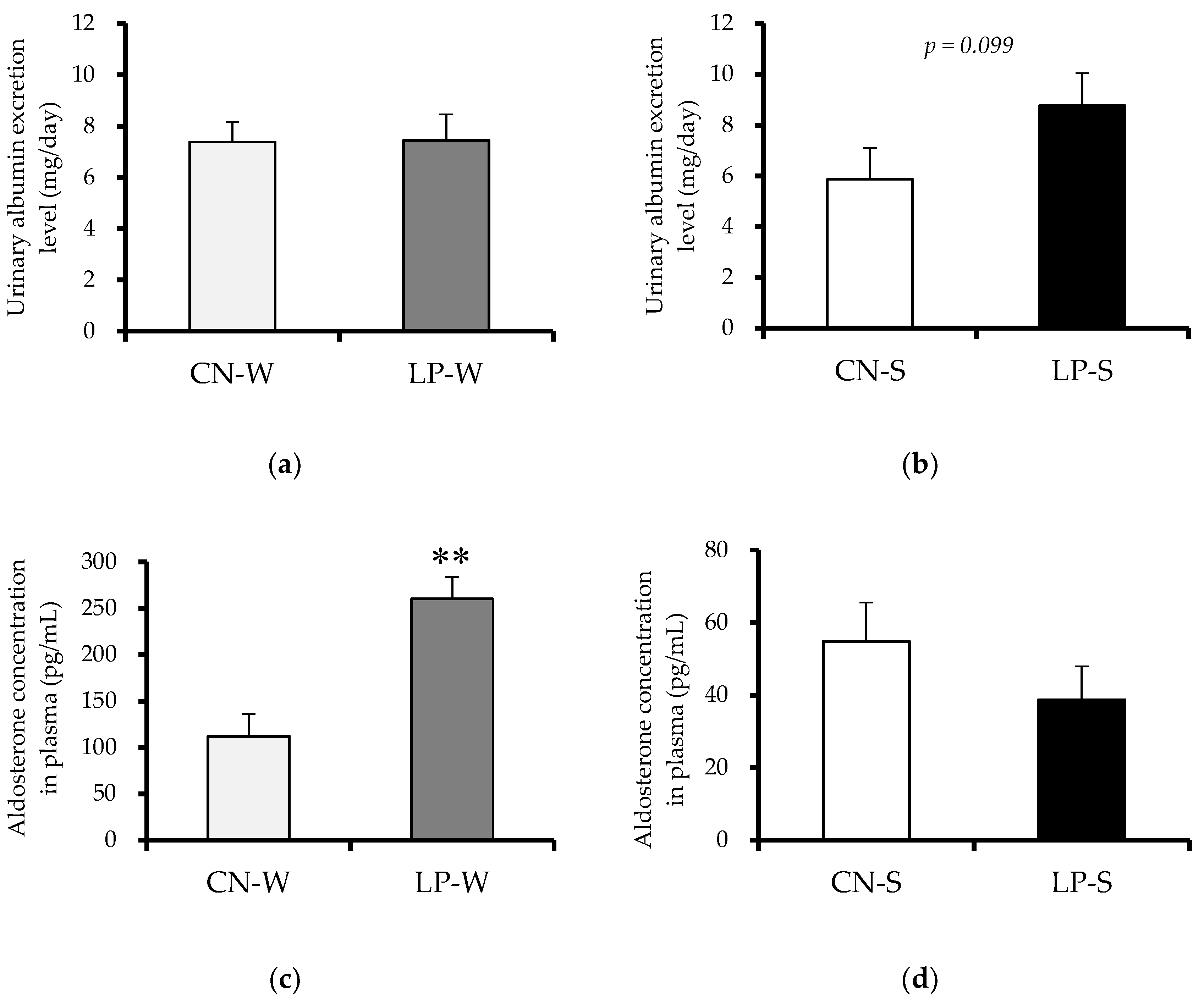
3.2. Offspring Organ Weight, and Urinary Albumin Excretion, and Plasma Aldosterone Levels at 12 Weeks after Birth
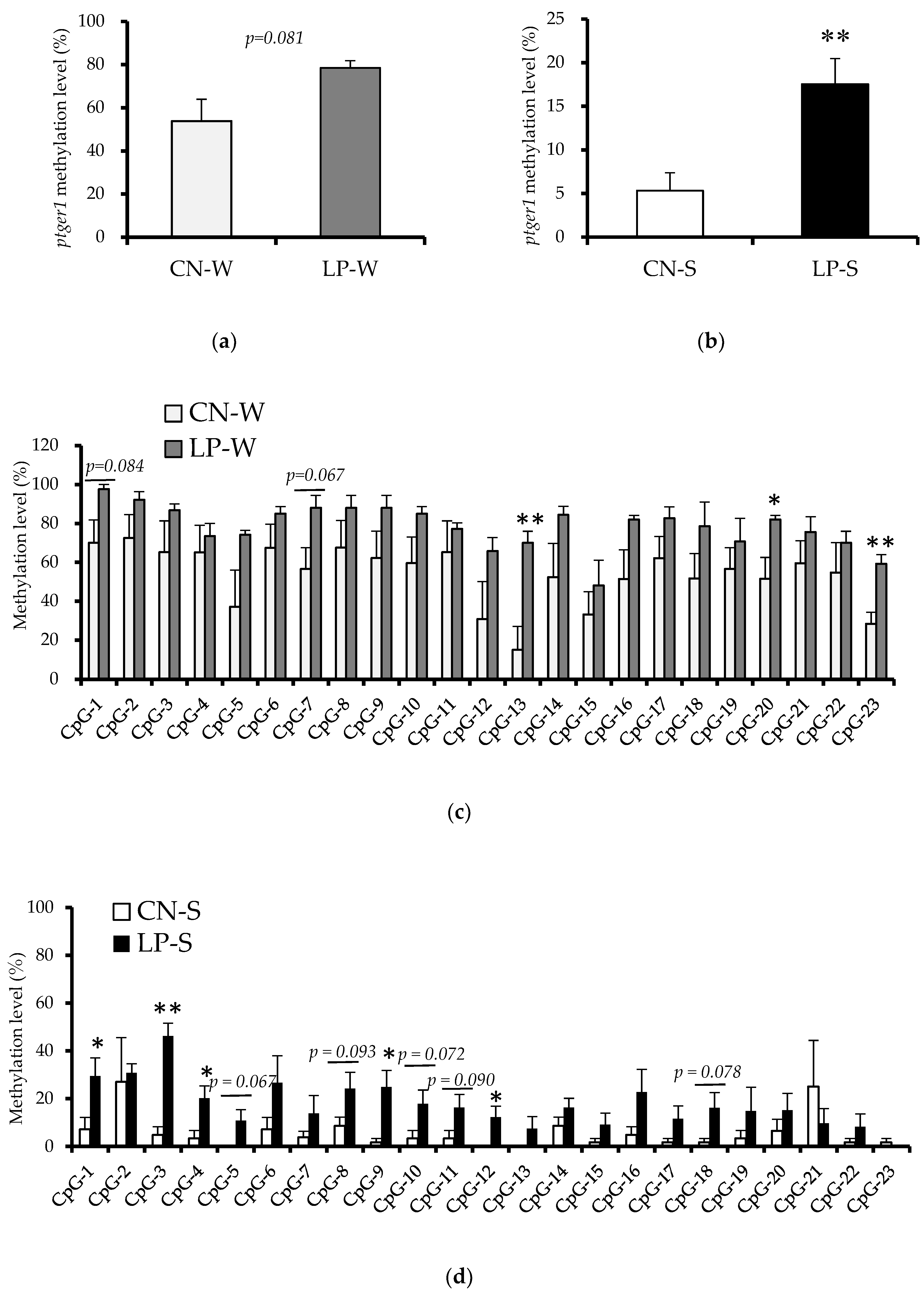
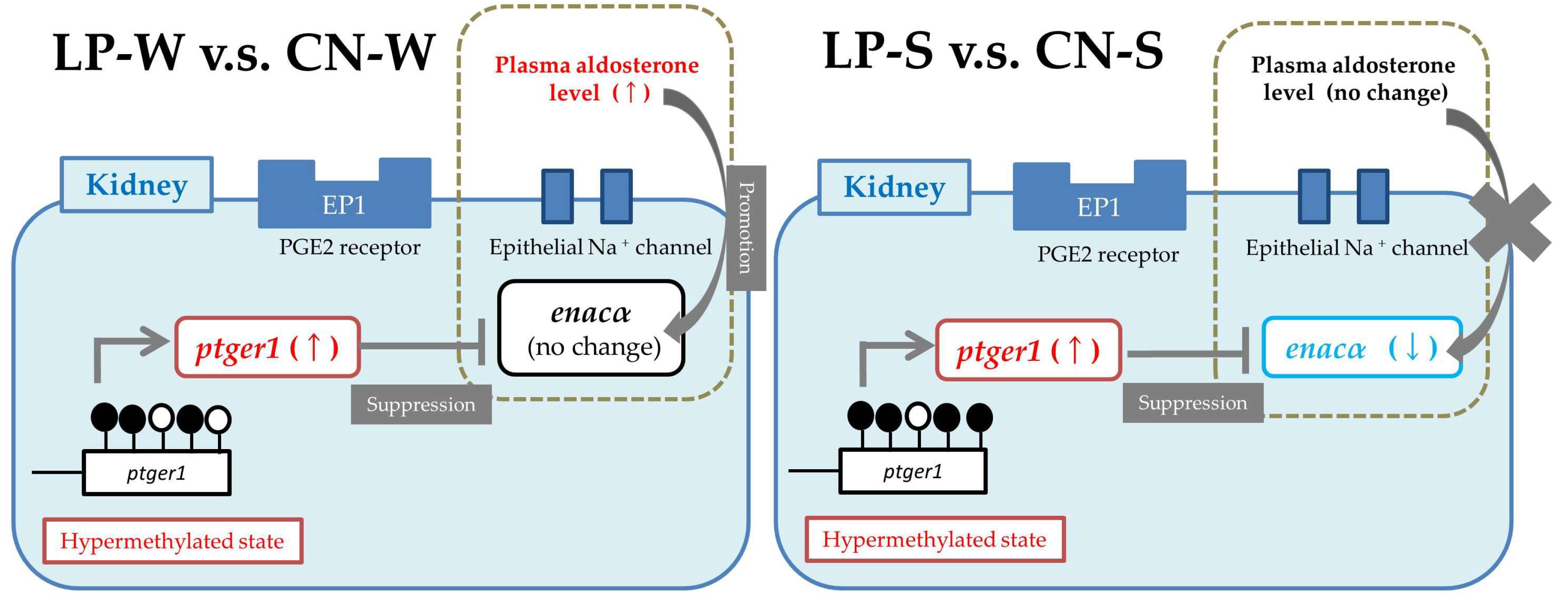
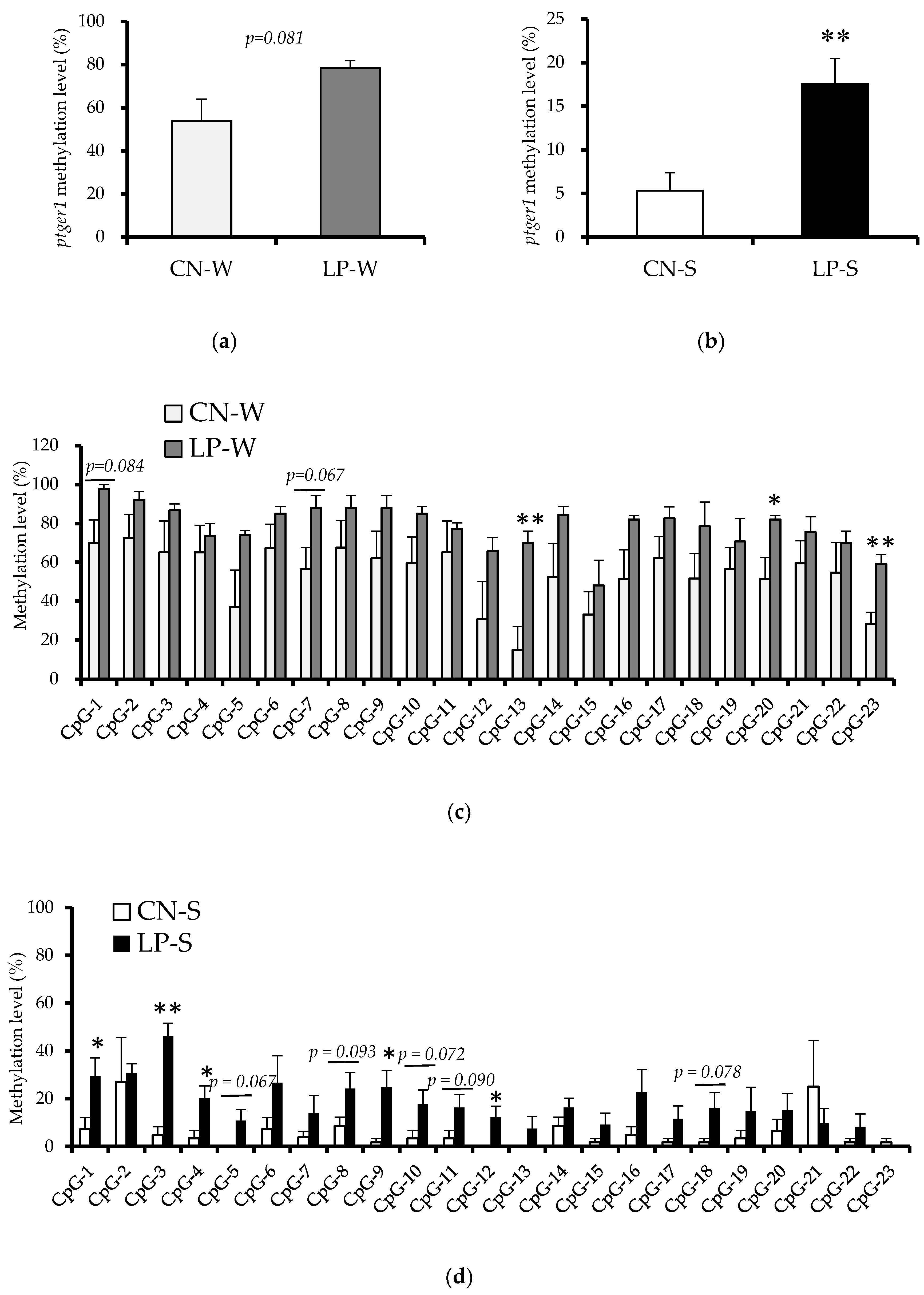
3.3. DNA Methylation Status in the Ptger1 Gene Region in Offspring Kidney Samples
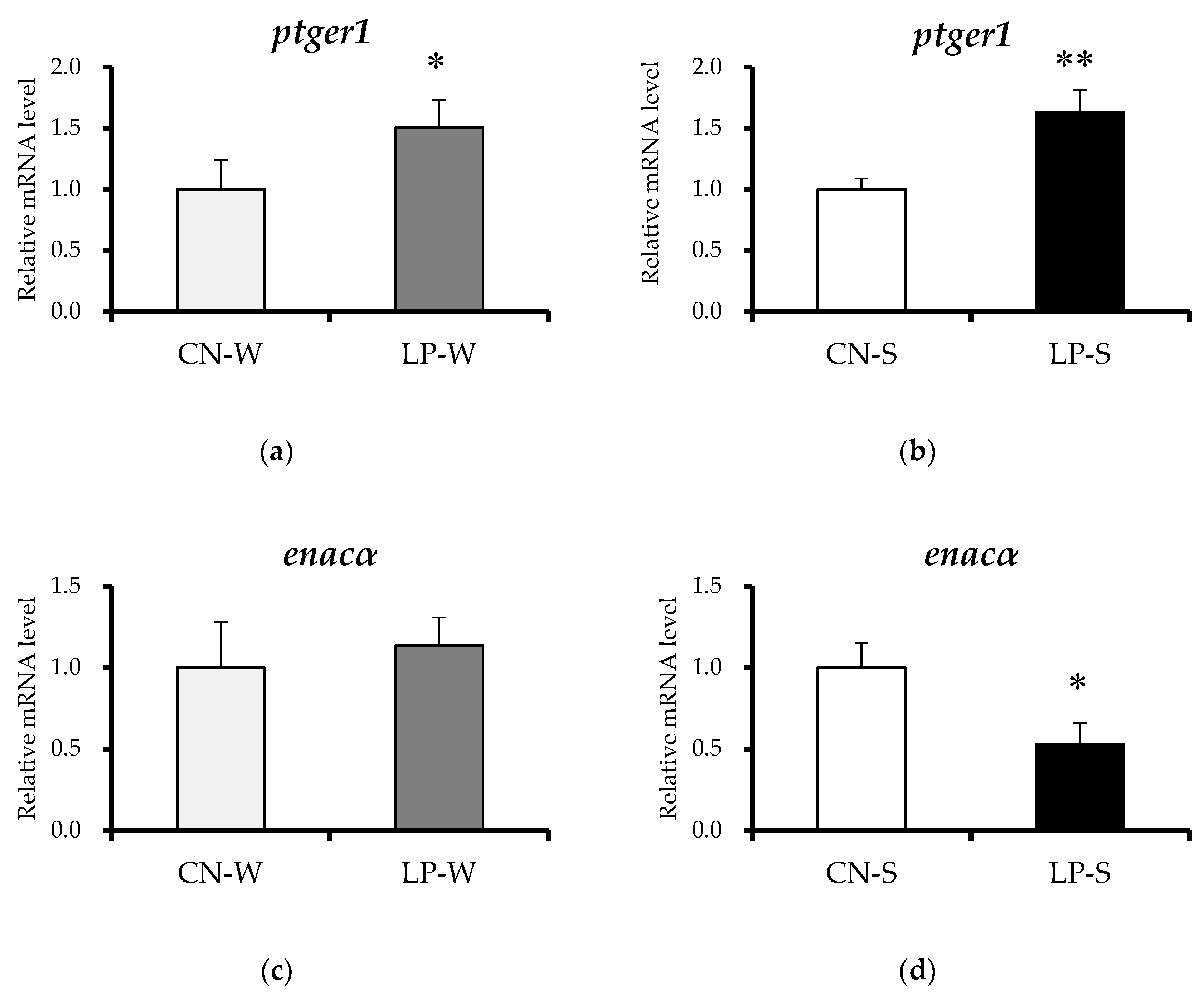
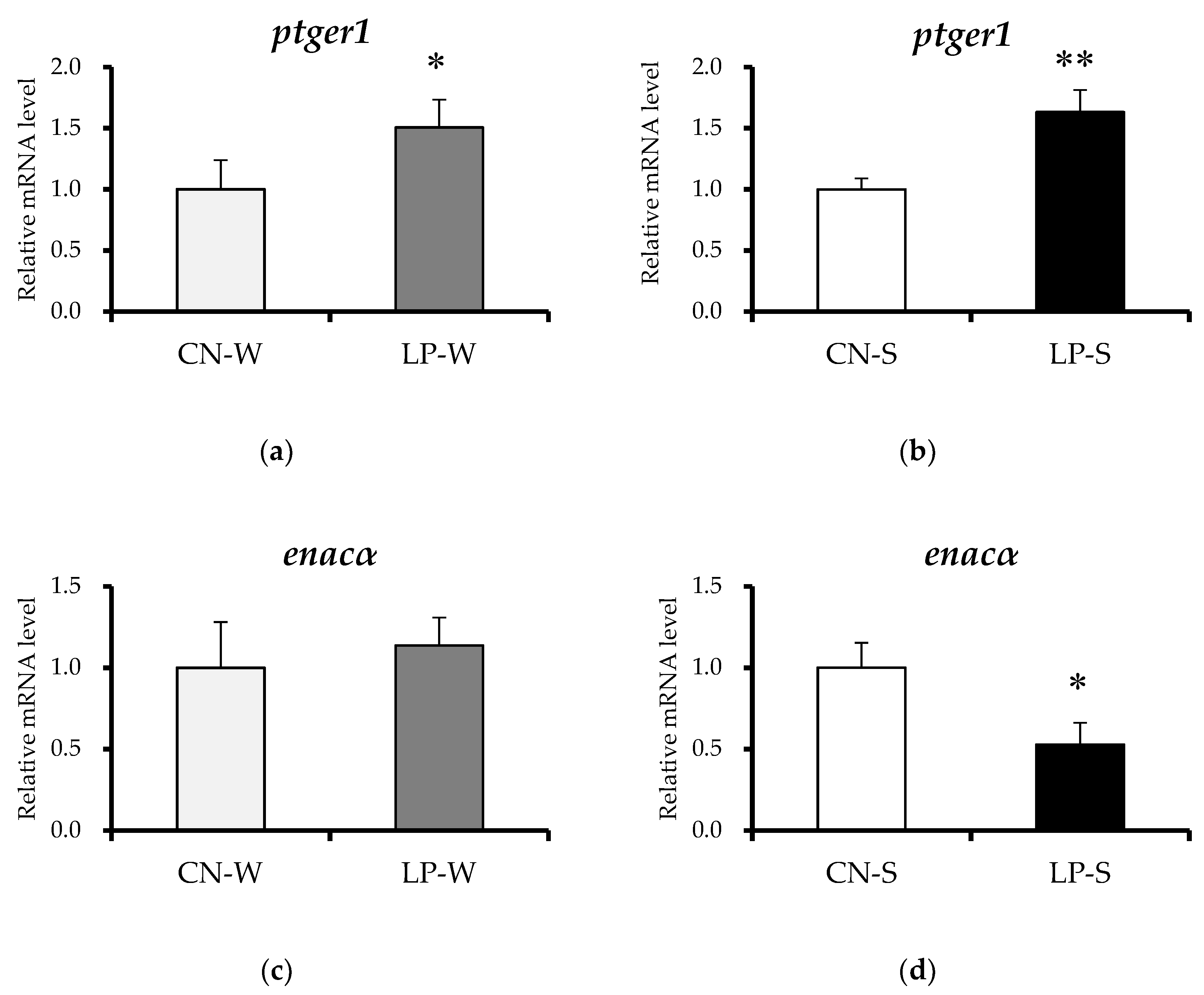
3.4. Offspring Renal Expression Of Ptger1-Related Genes
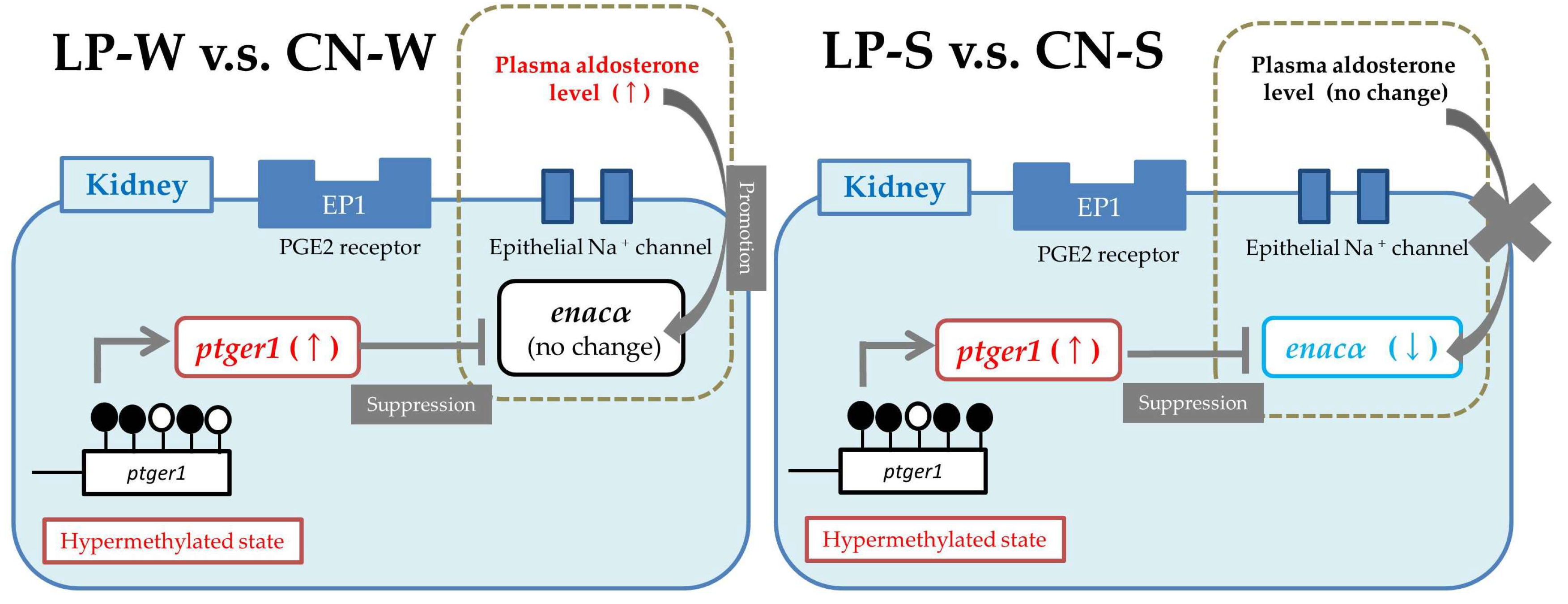
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Osmond, C.; Barker, D.J.; Winter, P.D.; Fall, C.H.; Simmonds, S.J. Early growth and death from cardiovascular disease in women. BMJ 1993, 307, 1519–1524. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J.; Martyn, C.N.; Osmond, C.; Hales, C.N.; Fall, C.H. Growth in utero and serum cholesterol concentrations in adult life. BMJ 1993, 307, 1524–1527. [Google Scholar] [CrossRef] [PubMed]
- Barker, D.J. Fetal origins of coronary heart disease. BMJ 1995, 311, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Roseboom, T.J.; van der Meulen, J.H.; Ravelli, A.C.; Osmond, C.; Barker, D.J.; Bleker, O.P. Effects of prenatal exposure to the Dutch famine on adult disease in later life: An overview. Mol. Cell. Endocrinol. 2001, 185, 93–98. [Google Scholar] [CrossRef]
- Barker, D.J. Fetal nutrition and cardiovascular disease in later life. Br. Med. Bull. 1997, 53, 96–108. [Google Scholar] [CrossRef] [PubMed]
- Remacle, C.; Bieswal, F.; Bol, V.; Reusens, B. Developmental programming of adult obesity and cardiovascular disease in rodents by maternal nutrition imbalance. Am. J. Clin. Nutr. 2011, 94, 1846S–1852S. [Google Scholar] [CrossRef] [PubMed]
- Van Straten, E.M.E.; Bloks, V.W.; van Dijk, T.H.; Baller, J.F.W.; Huijkman, N.C.A.; Kuipers, I.; Verkade, H.J.; Plösch, T. Sex-dependent programming of glucose and fatty acid metabolism in mouse offspring by maternal protein restriction. Gend. Med. 2012, 9, 166–179.e13. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Xiao, X.; Zhang, Q.; Yu, M. DNA methylation: The pivotal interaction between early-life nutrition and glucose metabolism in later life. Br. J. Nutr. 2014, 112, 1850–1857. [Google Scholar] [CrossRef] [PubMed]
- Burdge, G.C.; Hanson, M.A.; Slater-Jefferies, J.L.; Lillycrop, K.A. Epigenetic regulation of transcription: A mechanism for inducing variations in phenotype (fetal programming) by differences in nutrition during early life? Br. J. Nutr. 2007, 97, 1036–1046. [Google Scholar] [CrossRef] [PubMed]
- Hanson, M.; Godfrey, K.M.; Lillycrop, K.A.; Burdge, G.C.; Gluckman, P.D. Developmental plasticity and developmental origins of non-communicable disease: Theoretical considerations and epigenetic mechanisms. Prog. Biophys. Mol. Biol. 2011, 106, 272–280. [Google Scholar] [CrossRef] [PubMed]
- Ganu, R.S.; Harris, R.A.; Collins, K.; Aagaard, K.M. Early origins of adult disease: Approaches for investigating the programmable epigenome in humans, nonhuman primates, and rodents. ILAR J. 2012, 53, 306–321. [Google Scholar] [CrossRef] [PubMed]
- Otani, L.; Shirasaka, N.; Yoshizumi, H.; Murakami, T. The effects of maternal mild protein restriction on stroke incidence and blood pressure in stroke-prone spontaneously hypertensive rats (SHRSP). Biosci. Biotechnol. Biochem. 2004, 68, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Otani, L.; Sugimoto, N.; Kaji, M.; Murai, M.; Chang, S.-J.; Kato, H.; Murakami, T. Role of the renin-angiotensin-aldosterone system in the enhancement of salt sensitivity caused by prenatal protein restriction in stroke-prone spontaneously hypertensive rats. J. Nutr. Biochem. 2012, 23, 892–899. [Google Scholar] [CrossRef] [PubMed]
- Otani, L.; Yasumatsu, T.; Murakami, M.; Hayashi, A.; Kimoto, K.; Murakami, T. Reduction of salt sensitivity in stroke-prone spontaneously hypertensive rats administered an AT1 receptor antagonist during suckling. Am. J. Hypertens. 2006, 19, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Suganami, T.; Mori, K.; Tanaka, I.; Mukoyama, M.; Sugawara, A.; Makino, H.; Muro, S.; Yahata, K.; Ohuchida, S.; Maruyama, T.; et al. Role of prostaglandin E receptor EP1 subtype in the development of renal injury in genetically hypertensive rats. Hypertension 2003, 42, 1183–1190. [Google Scholar] [CrossRef] [PubMed]
- Narumiya, S.; Sugimoto, Y.; Ushikubi, F. Prostanoid receptors: Structures, properties, and functions. Physiol. Rev. 1999, 79, 1193–1226. [Google Scholar] [CrossRef] [PubMed]
- Breyer, M.D.; Jacobson, H.R.; Breyer, R.M. Functional and molecular aspects of renal prostaglandin receptors. J. Am. Soc. Nephrol. 1996, 7, 8–17. [Google Scholar] [PubMed]
- Altmann, S.; Murani, E.; Schwerin, M.; Metges, C.C.; Wimmers, K.; Ponsuksili, S. Dietary protein restriction and excess of pregnant German Landrace sows induce changes in hepatic gene expression and promoter methylation of key metabolic genes in the offspring. J. Nutr. Biochem. 2013, 24, 484–495. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Gu, P.; Liu, K.; Huang, P. Maternal protein restriction in rats leads to reduced PGC-1α expression via altered DNA methylation in skeletal muscle. Mol. Med. Rep. 2013, 7, 306–312. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, T.; Yamashita, S.; Ushijima, T.; Takumi, S.; Sano, T.; Michikawa, T.; Nohara, K. Genome-wide analysis of DNA methylation changes induced by gestational arsenic exposure in liver tumors. Cancer Sci. 2013, 104, 1575–1585. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Han, H.; De Carvalho, D.D.; Lay, F.D.; Jones, P.A.; Liang, G. Gene body methylation can alter gene expression and is a therapeutic target in cancer. Cancer Cell 2014, 26, 577–590. [Google Scholar] [CrossRef] [PubMed]
- Guyton, A.C.; Coleman, T.G.; Cowley, A.V; Scheel, K.W.; Manning, R.D.; Norman, R.A. Arterial pressure regulation. Overriding dominance of the kidneys in long-term regulation and in hypertension. Am. J. Med. 1972, 52, 584–594. [Google Scholar] [CrossRef]
- Gu, X.; Gu, D.; He, J.; Rao, D.C.; Hixson, J.E.; Chen, J.; Li, J.; Huang, J.; Wu, X.; Rice, T.K.; et al. Resequencing Epithelial Sodium Channel Genes Identifies Rare Variants Associated with Blood Pressure Salt-Sensitivity: The GenSalt Study. Am. J. Hypertens. 2018, 31, 205–211. [Google Scholar] [CrossRef] [PubMed]
- Canessa, C.M.; Schild, L.; Buell, G.; Thorens, B.; Gautschi, I.; Horisberger, J.D.; Rossier, B.C. Amiloride-sensitive epithelial Na+ channel is made of three homologous subunits. Nature 1994, 367, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Pácha, J.; Frindt, G.; Antonian, L.; Silver, R.B.; Palmer, L.G. Regulation of Na channels of the rat cortical collecting tubule by aldosterone. J. Gen. Physiol. 1993, 102, 25–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Xia, X.; Reisenauer, M.R.; Rieg, T.; Lang, F.; Kuhl, D.; Vallon, V.; Kone, B.C. Aldosterone-induced Sgk1 relieves Dot1a-Af9-mediated transcriptional repression of epithelial Na+ channel alpha. J. Clin. Investig. 2007, 117, 773–783. [Google Scholar] [CrossRef] [PubMed]
- González, A.A.; Céspedes, C.; Villanueva, S.; Michea, L.; Vio, C.P. E Prostanoid-1 receptor regulates renal medullary alphaENaC in rats infused with angiotensin II. Biochem. Biophys. Res. Commun. 2009, 389, 372–377. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component (%) | CN (20% Casein) | LP (9% Casein) |
|---|---|---|
| Casein | 20.0 | 9.0 |
| Corn starch | 66.8 | 77.8 |
| DL-Methionine | 0.2 | 0.2 |
| Soy bean oil | 5.0 | 5.0 |
| Vitamin mixture * | 1.0 | 1.0 |
| Mineral mixture * | 4.0 | 4.0 |
| Cellulose powder | 3.0 | 3.0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miyoshi, M.; Sato, M.; Saito, K.; Otani, L.; Shirahige, K.; Miura, F.; Ito, T.; Jia, H.; Kato, H. Maternal Protein Restriction Alters the Renal Ptger1 DNA Methylation State in SHRSP Offspring. Nutrients 2018, 10, 1436. https://doi.org/10.3390/nu10101436
Miyoshi M, Sato M, Saito K, Otani L, Shirahige K, Miura F, Ito T, Jia H, Kato H. Maternal Protein Restriction Alters the Renal Ptger1 DNA Methylation State in SHRSP Offspring. Nutrients. 2018; 10(10):1436. https://doi.org/10.3390/nu10101436
Chicago/Turabian StyleMiyoshi, Moe, Masayuki Sato, Kenji Saito, Lila Otani, Katsuhiko Shirahige, Fumihito Miura, Takashi Ito, Huijuan Jia, and Hisanori Kato. 2018. "Maternal Protein Restriction Alters the Renal Ptger1 DNA Methylation State in SHRSP Offspring" Nutrients 10, no. 10: 1436. https://doi.org/10.3390/nu10101436
APA StyleMiyoshi, M., Sato, M., Saito, K., Otani, L., Shirahige, K., Miura, F., Ito, T., Jia, H., & Kato, H. (2018). Maternal Protein Restriction Alters the Renal Ptger1 DNA Methylation State in SHRSP Offspring. Nutrients, 10(10), 1436. https://doi.org/10.3390/nu10101436