Consuming Genistein Improves Survival Rates in the Absence of Laxative in ΔF508-CF Female Mice

{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
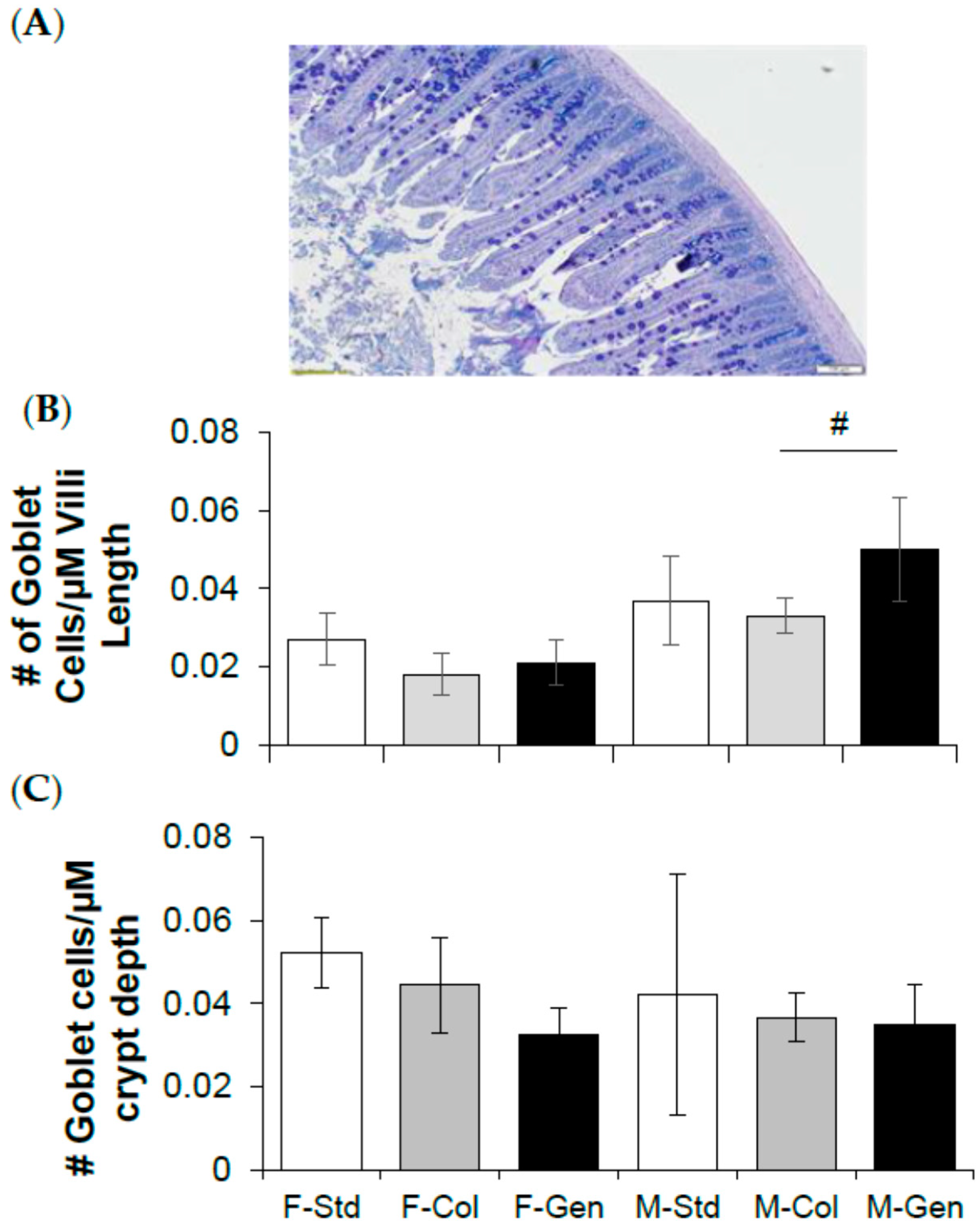
2.1. Histology
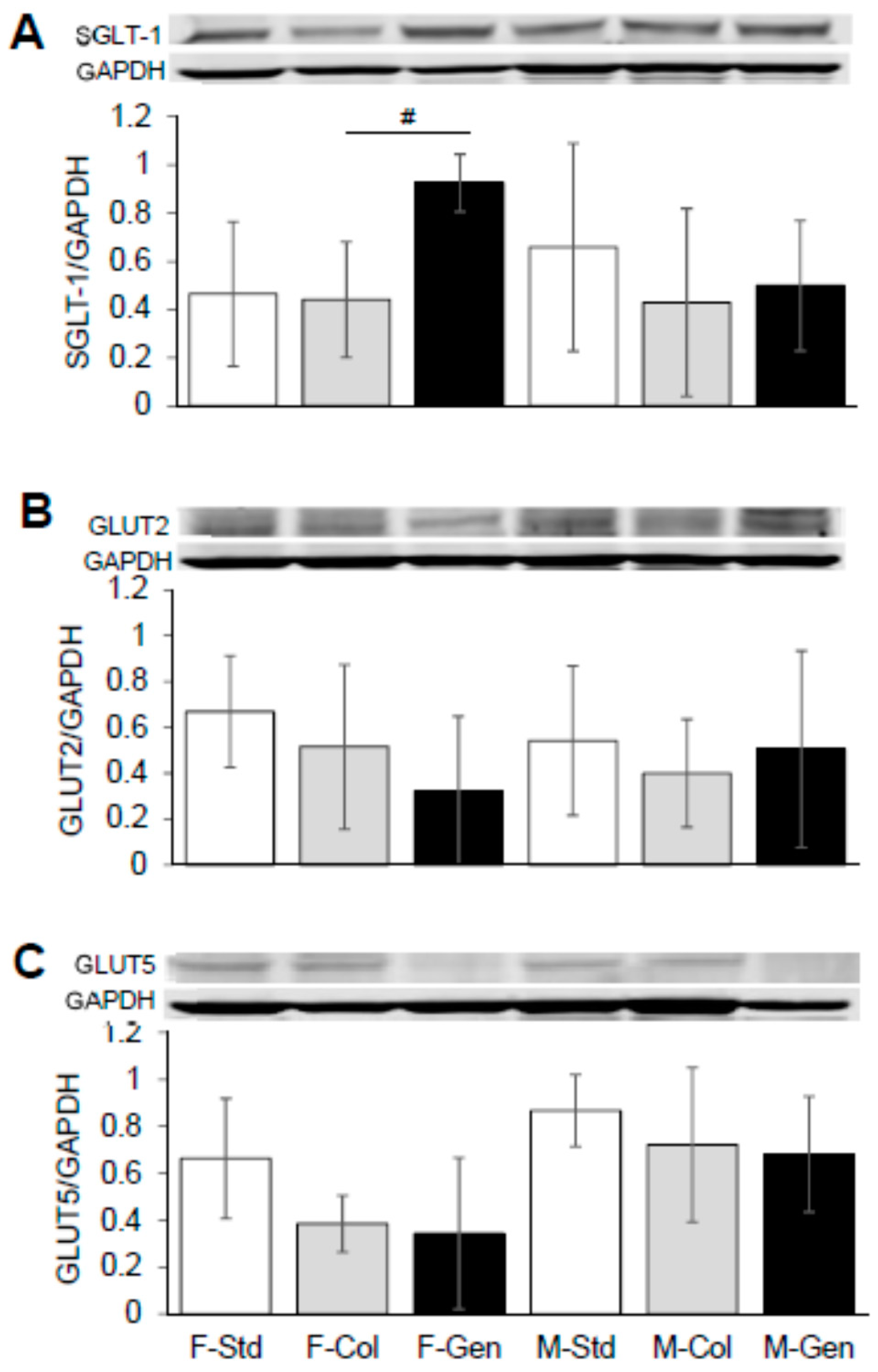
2.2. Western Blot Analysis
2.3. Statistics
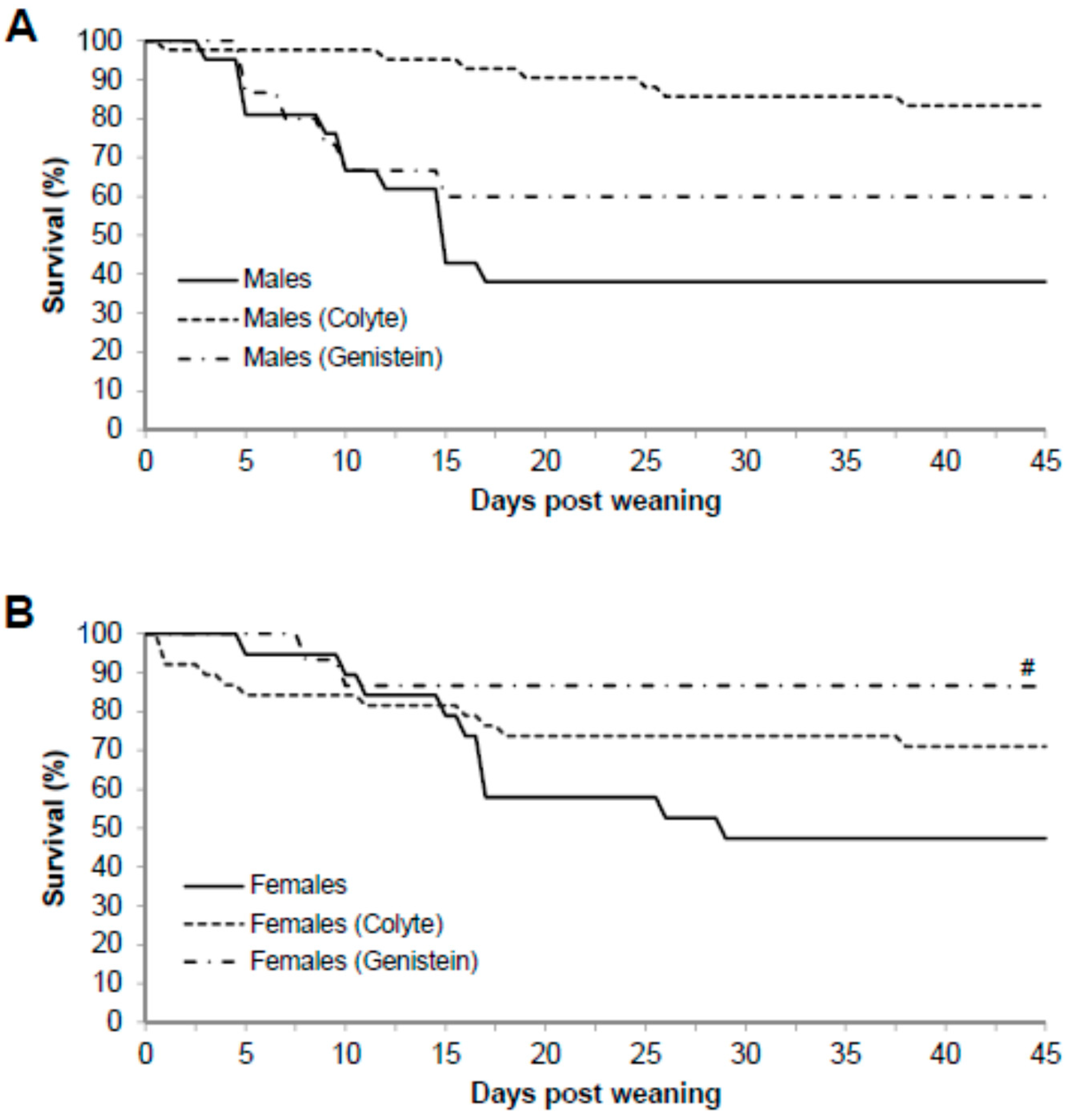
3. Results
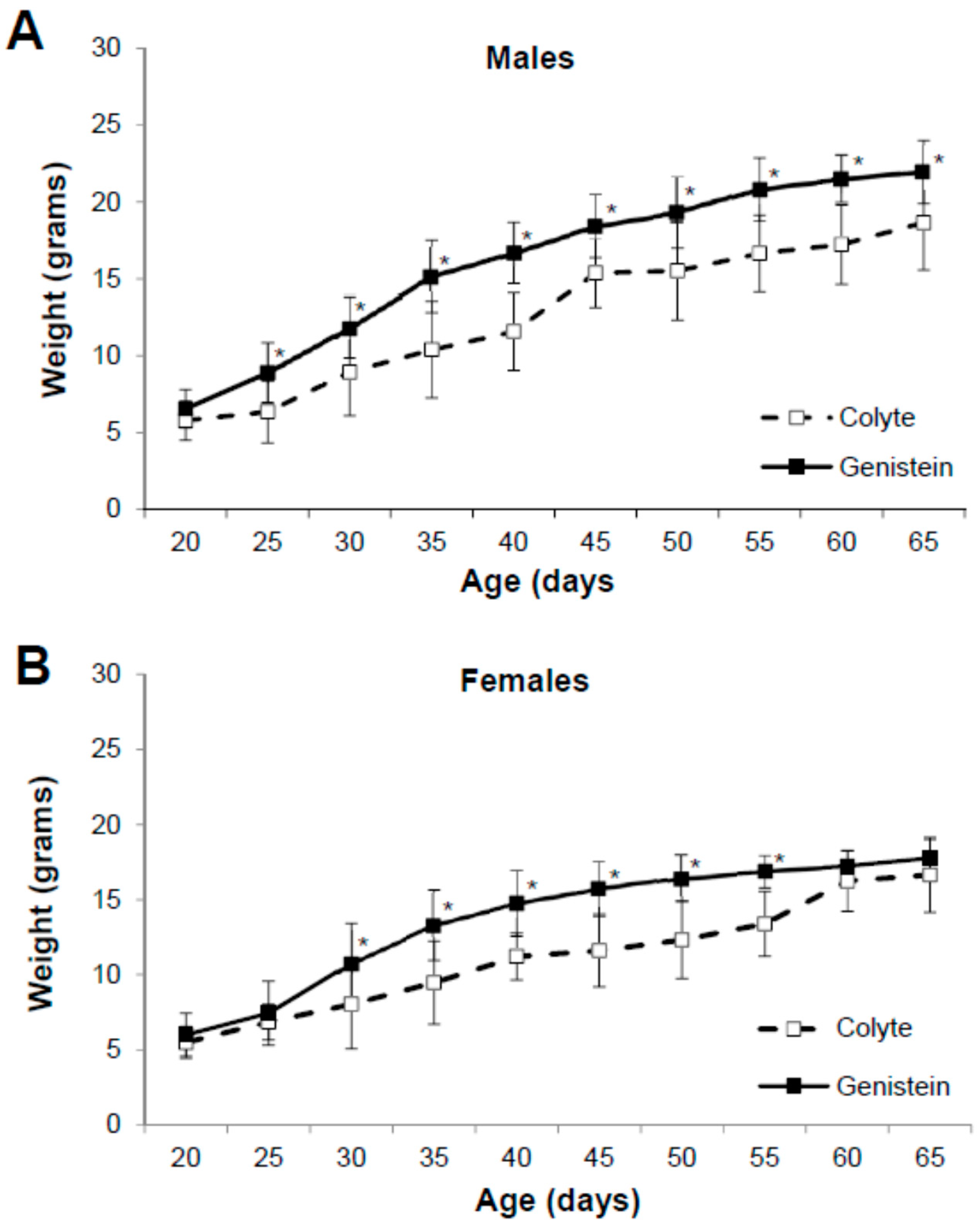
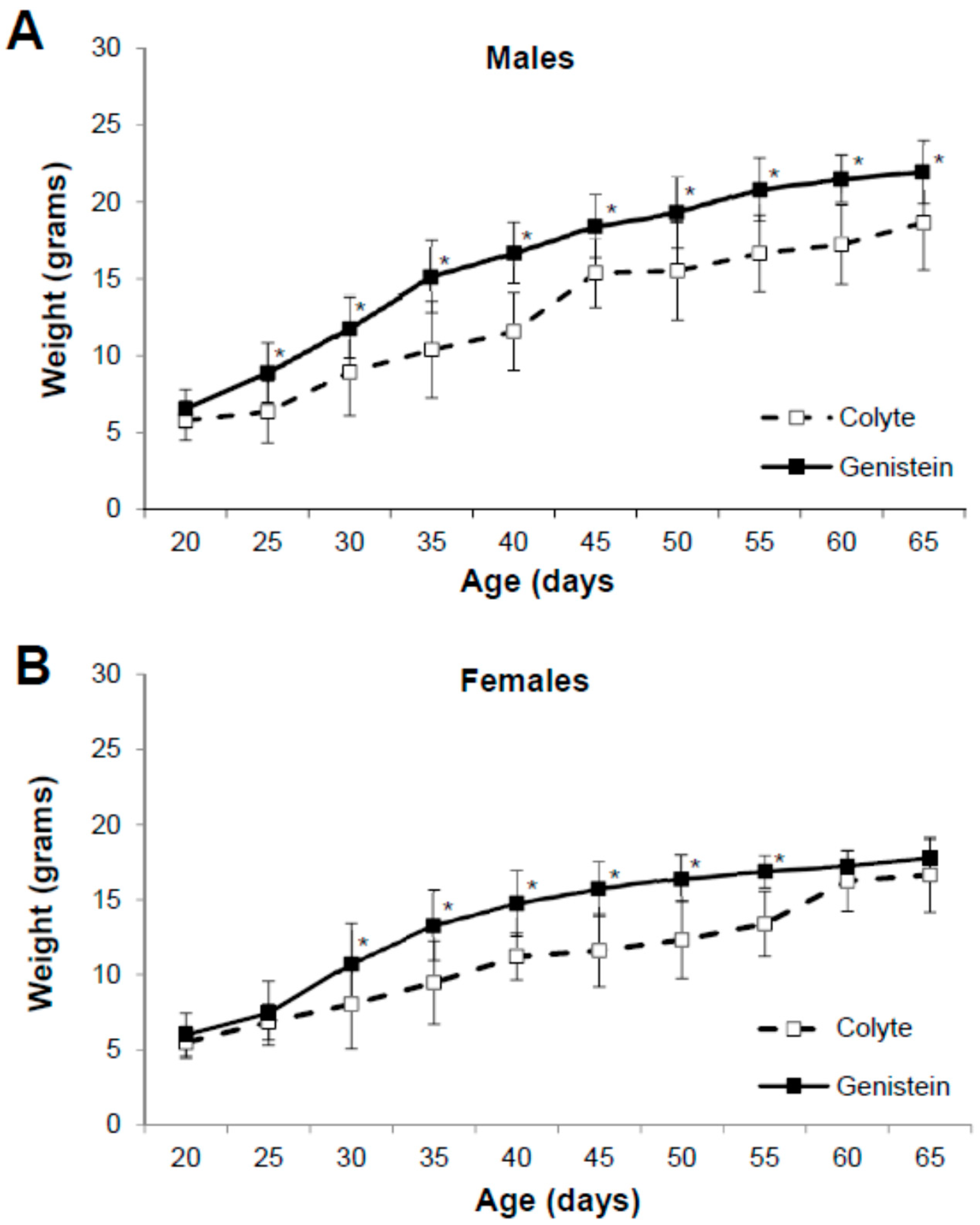
3.1. Physical Characteristics of the Mice
3.2. Jejunum Morphology
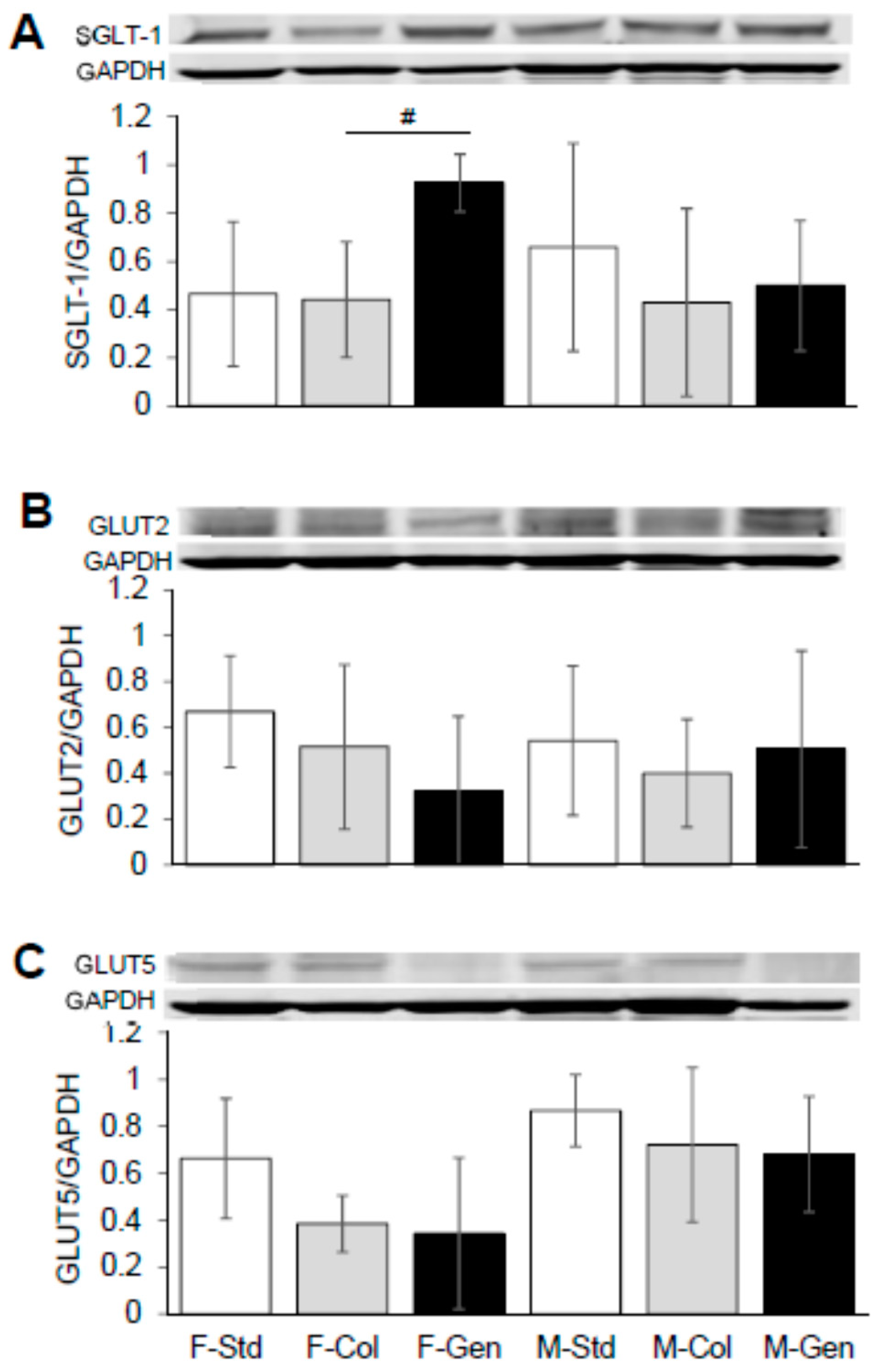
3.3. Expression of Key Proteins Involved in Absorption across Jejunum
4. Discussion
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Rommens, J.M.; Iannuzzi, M.C.; Kerem, B.; Drumm, M.L.; Melmer, G.; Dean, M.; Rozmahel, R.; Cole, J.L.; Kennedy, D.; Hidaka, N.; et al. Identification of the cystic fibrosis gene: Chromosome walking and jumping. Science 1989, 245, 1059–1095. [Google Scholar] [CrossRef] [PubMed]
- Drumm, M.L.; Wilkinson, D.J.; Smit, L.S.; Worrell, R.T.; Strong, T.V.; Frizzell, R.A.; Dawson, D.C.; Collins, F.S. Chloride conductance expressed by DF508 and other mutant CFTR’s in xenopus oocytes. Science 1991, 254, 1797–1799. [Google Scholar] [CrossRef] [PubMed]
- Cutting, G.R. Cystic fibrosis genetics: From molecular understanding to clinical application. Nat. Rev. Genet. 2015, 16, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Jennings, M.T.; Flume, P.A. Cystic fibrosis: Translating molecular mechanisms into effective therapies. Ann. Am. Thorac. Soc. 2018, 15, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Van Der Doef, H.P.; Kokke, F.T.; Van der Ent, C.K.; Houwen, R.H. Intestinal obstruction syndromes in cystic fibrosis: Meconium ileus, distal intestinal obstruction syndrome, and constipation. Curr. Gastroenterol. Rep. 2011, 13, 265–270. [Google Scholar] [CrossRef] [PubMed]
- Abraham, J.M.; Taylor, C.J. Cystic fibrosis and disroderes of the large intestine: DIOS, constipation, and colorectal cancer. J. Cyst. Fibros. 2017, 16 (Suppl. S2), S40–S49. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.L.; Harline, M.C. CFTR is required for cAMP inhibition of intestinal Na+ absorption in a cystic fibrosis mouse model. Am. J. Physiol. 1996, 270, G259–G267. [Google Scholar] [CrossRef] [PubMed]
- Grubb, B.R. Ion transport across the jejunum in normal and cystic fibrosis mice. Am. J. Physiol. 1995, 268, G505–G513. [Google Scholar] [CrossRef] [PubMed]
- De Lisle, R.C.; Mueller, R.; Boyd, M. Impaired mucosal barrier function in the small intestine of the cystsic fibrosis mouse. J. Pediatr. Gastroenterol. Nutr. 2011, 53, 371–379. [Google Scholar] [CrossRef] [PubMed]
- Snouwaert, J.N.; Brigman, K.K.; Latour, A.M.; Malouf, N.N.; Boucher, R.C.; Smithies, O.; Koller, B.H. An animal model for cystic fibrosis made by gene targetting. Science 1992, 257, 1083–1088. [Google Scholar] [CrossRef] [PubMed]
- De Lisle, R.C. Decreased expression of enterocyte nutrient assimilation genes and proteins in the small intestine of cystic fibrois mouse. J. Pediatr. Gastroenterol. Nutr. 2016, 62, 627–634. [Google Scholar] [CrossRef] [PubMed]
- Clarke, L.L.; Gawenis, L.R.; Franklin, C.L.; Harline, M.C. Increased survival of CFTR knockout mice with an oral osmotic laxative. Lab. Anim. Sci. 1996, 46, 612–618. [Google Scholar] [PubMed]
- Grubb, B.R.; Gabriel, S. Intestinal physiology and pathology in gene-targeted mouse models of cystic fibrosis. Am. J. Physiol. 1997, 273, G258–G266. [Google Scholar] [CrossRef] [PubMed]
- Zaheer, K.; Humayoun Akhtar, M. An updated review of dietary isoflavones: Nutrition, processing, bioavailability and impacts on human health. Crit. Rev. Food Sci. Nutr. 2017, 57, 1280–1293. [Google Scholar] [CrossRef] [PubMed]
- Leuner, O.; Havlik, J.; Hummelova, J.; Prokudina, E.; Novy, P.; Kokoska, L. Distribution of isoflavones and coumestrol in negelected tropical and subtropical legumes. J. Sci. Food Agric. 2013, 93, 575–579. [Google Scholar] [CrossRef] [PubMed]
- Murphy, P.A. Phytoestrogen content of processed soybean products. Food Technol. 1982, 36, 60–64. [Google Scholar]
- Messina, M.J. Emerging evidence on the role of soy in reducing prostate cancer risk. Nutr. Rev. 2003, 61, 117–131. [Google Scholar] [CrossRef] [PubMed]
- Kalsaiselvan, V.; Kalaivani, M.; Vijayakumar, A.; Sureshkumar, K.; Ventkateskumar, K. Current knowledge and future direction of research on soy isoflavones as therapeutic agents. Pharmacogn. Rev. 2010, 4, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Kirk, E.A.; Sutherland, P.; Wang, S.A.; Chait, A.; LeBoeuf, R.C. Dietary isoflavones reduce plasma cholesterol and atherosclerosis in C57BL/6 mice but not LDL receptor-deficient mice. J. Nutr. 1998, 128, 954–959. [Google Scholar] [CrossRef] [PubMed]
- Carlson, S.; Peng, N.; Prasain, J.; Wyss, J. Effects of botanical dietary supplements on cardiovascular, cognitive and metabolic function in males and females. Gend. Med. 2008, 5, S76–S90. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Liu, Y.; Wang, T.; Zhao, L.; Feng, W. Does genistein lower plasma lipids and homocysteine levels in postmenopausal women? A meta-analysis. Climacteric 2016, 19, 440–447. [Google Scholar] [CrossRef] [PubMed]
- Si, H.; Liu, D. Genistein, a soy phytoestrogen, upregulates the expression of human endothelial nitric oxide synthase and lowers blood pressure in spontaneously hypertesive rats. J. Nutr. 2008, 138, 297–304. [Google Scholar] [CrossRef] [PubMed]
- Dey, I.; Shah, K.; Bradbury, N.A. Natural compounds as therapeutic agents n the treatment of cystic fibrosis. J. Genet. Syndr. Gene Ther. 2016, 7, 284–294. [Google Scholar] [CrossRef] [PubMed]
- Hwang, T.-C.; Wang, F.; Zeltwanger, S.; Yang, I.; Reenstra, W. Genistein potentiates wild-type and ÐF508-CFTR channel activity. Am. J. Physiol. 1997, 273, C988–C998. [Google Scholar] [CrossRef] [PubMed]
- Al-Nakkash, L.; Hwang, T.-C. Activation of wild-type and ΔF508-CFTR by phosphodiesterase inhibitors through cAMP-dependent and -independent mechanisms. Pfluegers Arch. 1999, 437, 553–561. [Google Scholar] [CrossRef]
- Ai, T.; Bompadre, S.G.; Wang, X.; Hu, S.; Li, M.; Hwang, T.C. Capsaicin potentiates wild type and mutant cystic fibrosis transmembrane conductance regulator chloride-channel currents. Mol. Pharmacol. 2004, 65, 1415–1426. [Google Scholar] [CrossRef] [PubMed]
- Arora, K.; Yarlaggada, S.; Zhang, W.; Moon, C.; Bouquet, E.; Srinivasan, S.; Li, C.; Stokes, D.C.; Naren, A.P. Personalize dmedicine in cystic fibrosis:genisetin supplementation as a treatment for patients with rare S1045Y-CFTR mutation. Am. J. Physiol. 2016, 311, L364–L374. [Google Scholar]
- Dekkers, J.F.; Van Mourik, P.; Vonk, A.M.; Kruisselbrink, E.; Berkers, G.; de Winter-de Groot, K.M.; Janssens, H.M.; Bronsveld, I.; van der Ent, C.K.; de Jonge, H.R.; et al. Potentiator synergy in rectal organoids carrying S1251N, G551D, or F508del CFTR mutations. J. Cyst. Fibros. 2016, 15, 568–578. [Google Scholar] [CrossRef] [PubMed]
- Al-Nakkash, L.; Hu, S.; Li, M.; Hwang, T.-C. A common mechanism for cystic fibrosis transmembrane conductance regulator protein activation by genistein and benzimidazolone analogs. J. Pharmacol. Exp. Ther. 2001, 296, 464–472. [Google Scholar] [PubMed]
- Wang, F.; Zeltwanger, S.; Yang, I.; Nairn, A.; Hwang, T.-C. Actions of genistein on CFTR channel gating. J. Gen. Physiol. 1998, 111, 477–490. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wang, H.; Murphy, P.; Cook, L.; Hendrich, S. Daidzein is a more bioavailable soymilk isoflavone than is genistein in adult women. J. Nutr. 1994, 124, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Barnes, S.; Sfakianos, J.; Coward, L.; Kirk, M. Soy isoflavonoids and cancer prevention. Underlying biochemical and pharmacological issues. Adv. Exp. Med. Biol. 1996, 401, 87–100. [Google Scholar] [PubMed]
- Hsieh, C.Y.; Santell, R.; Haslam, S.; Helferich, W. Estrogenic effects of genistein on the growth of estrogen receptor-positive human breast cancer (MCF-7) cells in vitro and in vivo. Cancer Res. 1998, 58, 3833–3838. [Google Scholar] [PubMed]
- Al-Nakkash, L.; Clarke, L.L.; Rottinghaus, G.E.; Chen, Y.J.; Cooper, K.; Rubin, L.J. Dietary genistein stimulates anion secretion across female murine intestine. J. Nutr. 2006, 136, 2785–2790. [Google Scholar] [CrossRef] [PubMed]
- Cassidy, A.; Brown, J.E.; Hawdon, A.; Faughnan, M.S.; King, L.J.; Millward, J.; Zimmer-Nechemias, L.; Wolfe, B.; Setchell, K.D. Factors affecting the bioavailability of soy isoflavones in humans after ingestion of physiologically relevent levels from different soys foods. J. Nutr. 2006, 136, 45–51. [Google Scholar] [CrossRef] [PubMed]
- Zeiher, B.G.; Eichwald, E.; Zabner, J.; Smith, J.J.; Puga, A.P.; McCray, P.B.; Capecchi, M.R.; Welsh, M.J.; Thomas, K.R. A mouse model for the delta F508 allele of cystic fibrosis. J. Clin. Investig. 1995, 96, 2051–2064. [Google Scholar] [CrossRef] [PubMed]
- Koller, B.H.; Kim, H.S.; Latour, A.M.; Brigman, K.; Boucher, R.C.; Scambler, P.; Wainwright, B.; Smithies, O. Toward an animal model of cystic fibrosis: Targeted interruption of exon 10 in the cystic fibrosis transmebrane regulator gene in embryonic stem cells. Proc. Natl. Acad. Sci. USA 1991, 88, 10730–10734. [Google Scholar] [CrossRef] [PubMed]
- Minguez, M.; Lopez-Higueras, A.; Judez, J. Use of polyethylene glycol in functional constipation and fecal impaction. Rev. Esp. Enferm. Dig. 2016, 108, 790–806. [Google Scholar] [CrossRef] [PubMed]
- Home Office HoC. Code of Practice for the Housing and Care of Animals Bred, Supplied or Used for Scientific Purposes. Available online: https://www.gov.uk/government/publications/code-of-practice-for-the-housing-and-care-of-animals-bred-supplied-or-used-for-scientific-purposes (accessed on 1 June 2015).
- Al-Nakkash, L.; Batia, L.; Bhakta, M.; Peterson, A.; Hale, N.; Skinner, R.; Sears, S.; Jensen, J. Stimulation of murine intestinal secretion by daily genistein injections: Gender-dependent differences. Cell. Physiol. Biochem. 2011, 28, 239–250. [Google Scholar] [CrossRef] [PubMed]
- Leung, L.; Kang, J.; Rayyan, E.; Bhakta, A.; Barrett, B.; Larsen, D.; Jelinek, R.; Willey, J.; Cochran, S.; Broderick, T.B.; et al. Decreased basal chloride secretion and altered CFTR, villin and GLUT5 protein expression in jejunum from ob/ob mice. Diabetes Metab. Syndr. Obes. Targets Ther. 2014, 7, 321–330. [Google Scholar]
- Catmull, S.; Masood, F.; Schacht, S.; Dolan, R.; Stegman, D.; Leung, L.; Al-Nakkash, L. Dietary genistein rescues reduced basal chloride secretion in diabetic jejunum via sex-dependent mechanisms. Cell. Physiol. Biochem. 2016, 40, 335–346. [Google Scholar] [CrossRef] [PubMed]
- Tuo, B.; Wen, G.; Song, P.; Xu, J.; Liu, X.; Seidler, U.; Dong, H. Genistein stimulates duodenal HCO3− secretion through PI3K pathway in mice. Eur. J. Pharmacol. 2011, 651, 159–167. [Google Scholar] [CrossRef] [PubMed]
- Baker, M.J.; Hamilton, K.L. Genistein stimulates electrogenic Cl− secretion in the mouse jejunum. Am. J. Physiol. 2004, 287, C1636–C1645. [Google Scholar] [CrossRef] [PubMed]
- Shin, D.H.; Lee, M.J.; Jiao, H.Y.; Choi, S.; Kim, M.W.; Park, C.G.; Na, J.; Kim, S.W.; Park, I.K.; So, I.; et al. Regulatory roles of endogenous mitogen-activated protein kinases and tyrosine kinases in the pacemaker activity of colonic interstitial cells of Cajal. Pharmacology 2015, 96, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Shahi, P.K.; Choi, S.; Zuo, D.C.; Kim, M.Y.; Park, C.G.; Kim, Y.D.; Lee, J.; Park, K.J.; So, I.; Jun, J.Y. The possible roles of hyperpolarization-activated cyclic nucleotide channels in regulating pacemaker activity in colonic interstitial cells of Cajal. J. Gastroenterol. 2014, 49, 1001–1010. [Google Scholar] [CrossRef] [PubMed]
- Mall, M.; Wissner, A.; Seydewitz, H.; Huber, M.; Kuehr, J.; Brandis, M.; Greger, R.; Kunzelmann, K. Effect of genistein on native epithelial tissue from normal individuals and CF patients and on ion channels expressed in Xenopus oocytes. Br. J. Pharm. 2000, 130, 1884–1892. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Liu, J.; Loizidou, A.; Bugeja, L.A.; Warner, R.; Hawley, B.R.; Cai, Z.; Toye, A.M.; Sheppard, D.N.; Li, H. CFTR potentiators partially restore channel function to A561E-CFTR, a cystic fibrosis mutatnt with a similar mechanism of dysfunction as F508-del-CFTR. Br. J. Pharm. 2014, 171, 4490–4503. [Google Scholar] [CrossRef] [PubMed]
- Hendrich, S.; Lee, K.W.; Xu, X.; Wang, H.J.; Murphy, P.A. Defining food components as new nutrients. J. Nutr. 1994, 124, 1789S–1792S. [Google Scholar] [CrossRef] [PubMed]
- Santell, R.C.; Chang, Y.C.; Nair, M.G.; Helferich, W.G. Dietary genistein exerts estrogenic effects upon the uterus, mammary gland and the hypothalamic/pituitary axis in rats. J. Nutr. 1997, 127, 263–269. [Google Scholar] [CrossRef] [PubMed]
- Bhandari, A.; Crawford, S.E.; Huang, L.; Reenstra, W.W. Effects of oral genistein in mice. Pediatr. Pathol. Mol. Med. 2003, 22, 131–141. [Google Scholar] [CrossRef] [PubMed]
- Hirasaka, K.; Saito, S.; Yamaguchi, S.; Miyazaki, R.; Wang, Y.; Haruna, M.; Taniyama, S.; Higashitani, A.; Terao, J.; Nikawa, T.; et al. Dietary supplementation with isoflavones prevents muscle wasting in tumor-bearing mice. J. Nutr. Sci. Vitaminol. 2016, 62, 178–184. [Google Scholar] [CrossRef] [PubMed]
- Heinke, B.; Ribeiro, R.; Diener, M. Involvement of calmodulin and protein kinase C in the regulation of K+ transport by carbachol across the rat distal colon. Eur. J. Pharmacol. 1999, 377, 75–80. [Google Scholar] [CrossRef]
- Millar, G.A.; Hardin, J.A.; Johnson, L.R.; Gall, D.G. The role of PI 3-kinase in EGF-stimulated jejunal glucose transport. Can. J. Phys. Pharm. 2002, 80, 77–84. [Google Scholar] [CrossRef]
- Zhai, X.; Lin, M.; Zhang, F.; Hu, Y.; Xu, Y.; Li, Y.; Liu, K.; Ma, X.; Tian, X.; Yao, J. Dietary flavonoid genistein induces Nrf2 and phase II detoxification gene expression via ERKs and PKC pathwyas and protects against oxidative stress in Caco-2 cells. Mol. Nutr. Food Res. 2013, 57, 249–259. [Google Scholar] [CrossRef] [PubMed]
- Wegner, C.C.; Zhou, X.; Ding, Z.-M.; Kuo, M.T.; Carson, D.D. Tyrosine kinase inhibition decreases muc-1 expression in mouse epithelial cells. J. Cell. Physiol. 1997, 170, 200–208. [Google Scholar] [CrossRef]
- Kamboh, A.A.; Zhu, W.-Y. Individual and combined effects of genistein and hesperidin on immunity and intestinal morphology in lipopolysacharide-challenged broiler chikens. Poult. Sci. 2014, 93, 2175–2183. [Google Scholar] [CrossRef] [PubMed]
- Davis, P.B. The gender gap in cystic fibrosis survival. J. Gend. Specif. Med. 1999, 2, 47–51. [Google Scholar] [PubMed]
- Kulich, M.; Rosenfeld, M.; Goss, C.H.; Wilmott, R. Improved survival among young patients with cystic fibrosis. J. Pediatr. 2003, 142, 631–636. [Google Scholar] [CrossRef] [PubMed]


© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lord, R.; Fairbourn, N.; Mylavarapu, C.; Dbeis, A.; Bowman, T.; Chandrashekar, A.; Banayat, T.; Hodges, C.A.; Al-Nakkash, L. Consuming Genistein Improves Survival Rates in the Absence of Laxative in ΔF508-CF Female Mice. Nutrients 2018, 10, 1418. https://doi.org/10.3390/nu10101418
Lord R, Fairbourn N, Mylavarapu C, Dbeis A, Bowman T, Chandrashekar A, Banayat T, Hodges CA, Al-Nakkash L. Consuming Genistein Improves Survival Rates in the Absence of Laxative in ΔF508-CF Female Mice. Nutrients. 2018; 10(10):1418. https://doi.org/10.3390/nu10101418
Chicago/Turabian StyleLord, Ryan, Nathan Fairbourn, Charisma Mylavarapu, Ammer Dbeis, Taylor Bowman, Archana Chandrashekar, Tatum Banayat, Craig A. Hodges, and Layla Al-Nakkash. 2018. "Consuming Genistein Improves Survival Rates in the Absence of Laxative in ΔF508-CF Female Mice" Nutrients 10, no. 10: 1418. https://doi.org/10.3390/nu10101418