Sodium Citrate Increases Expression and Flux of Mg2+ Transport Carriers Mediated by Activation of MEK/ERK/c-Fos Pathway in Renal Tubular Epithelial Cells

 and
and
Abstract
:
1. Introduction
2. Experimental Section
2.1. Materials
2.2. Cell Culture
2.3. RNA Isolation and Quantitative Real-Time Reverse-Transcription Polymerase Chain Reaction (PCR)
2.4. Preparation of Cytoplasmic Extracts and Western Blotting
2.5. Immunoprecipitation
2.6. Luciferase Reporter Assay
2.7. Mg2+ Influx Assay
2.8. Reactive Oxygen Species (ROS) Production
2.9. Chromatin Immunoprecipitation Assay
2.10. Statistics
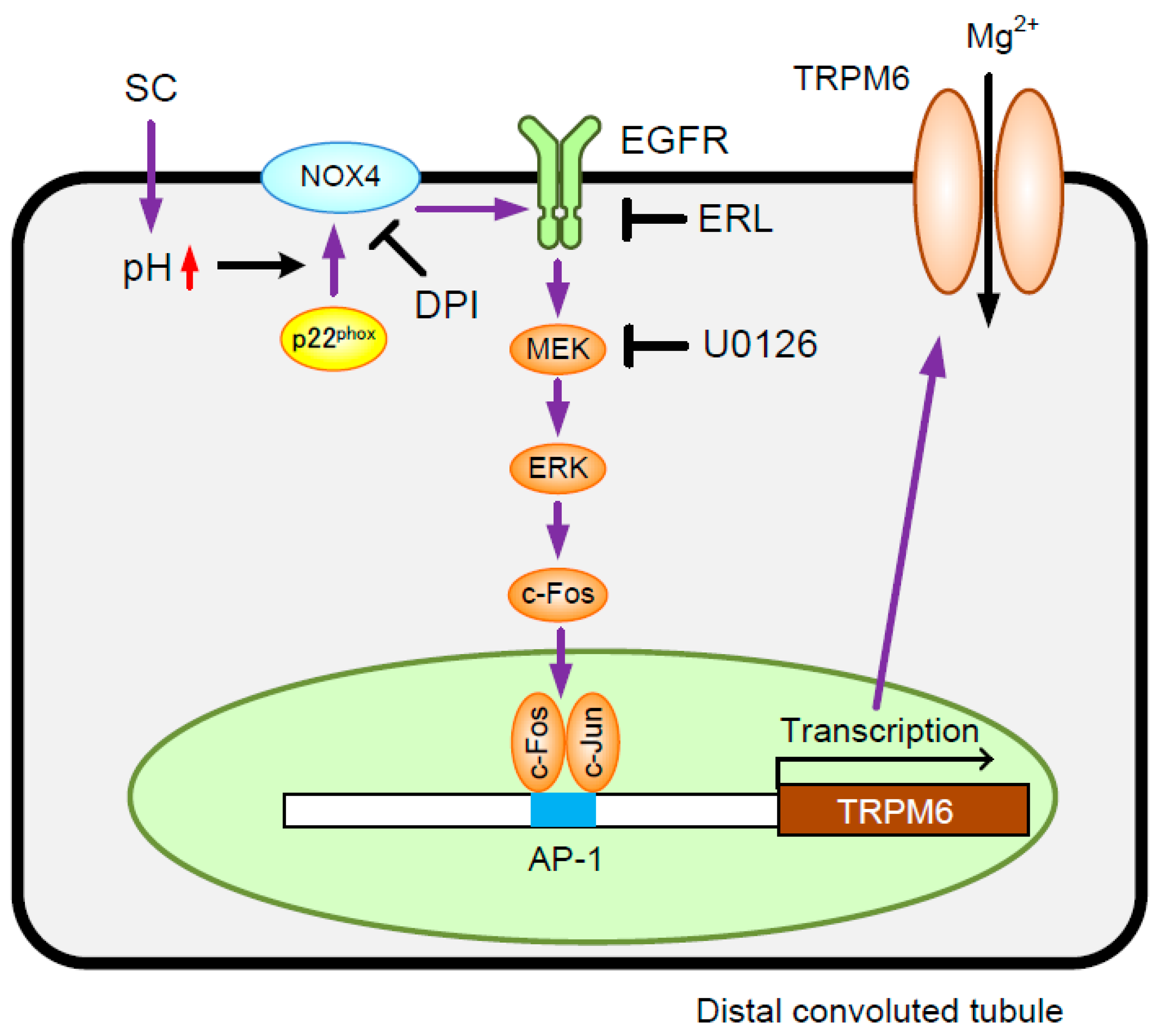
3. Results
3.1. Increase in TRPM6 Expression by Sc and Calcium Citrate (CC)
3.2. Increase in TRPM6 Expression by Alkalization
3.3. Effects of SC and Alkalization on Mg2+ Influx
3.4. Effects of SC and Alkalization on TRPM6 Promoter Activity
3.5. Involvement of NADPH Oxidase in SC-Induced Elevation of TRPM6 Expression
3.6. Association of c-Fos with the TRPM6 Promoter Region
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| CC | calcium citrate |
| ChIP | chromatin immunoprecipitation |
| CitM | Mg2+-citrate transporter |
| CNNM2 | cyclin M2 |
| Ct | threshold cycle |
| DCFDA | 2′,7′-dichlorofluorescin diacetate |
| DCT | distal convoluted tubule |
| DPI | diphenyleneiodonium |
| EGFR | epidermal growth factor receptor |
| EIPA | 5-(N-ethyl-N-isopropyl) amiloride |
| ERK | extracellular signal-regulated kinase |
| HBSS | Hank’s balanced salt solution |
| Mag-fura-2 AM | 2-[5-[2-[(acetyloxy)methoxy]-2-oxoethoxy]-6-[bis[2-[(acetyloxy)methoxy]-2-oxoethyl]amino]-2-benzofuranyl]-(acetyloxy)methyl ester |
| Mg2+ | magnesium ion |
| [Mg2+]i | intracellular free Mg2+ concentration |
| MEK | mitogen-activated protein kinase kinase |
| NaDC | sodium-dicarboxylate co-transporter |
| NADPH | nicotinamide-adenine dinucleotide phosphate |
| NOX | NADPH oxidase |
| PCR | polymerase chain reaction |
| p-ERK1/2 | phosphorylated ERK1/2 |
| PVDF | polyvinylidene fluoride |
| SC | sodium citrate |
| SDS-PAGE | sodium dodecyl sulfate-polyacrylamide gel electrophoresis |
| TAL | thick ascending limb of Henle’s loop |
| TRPM6 | transient receptor potential melastatin 6 |
References
- Takayanagi, K.; Shimizu, T.; Tayama, Y.; Ikari, A.; Anzai, N.; Iwashita, T.; Asakura, J.; Hayashi, K.; Mitarai, T.; Hasegawa, H. Downregulation of transient receptor potential M6 channels as a cause of hypermagnesiuric hypomagnesemia in obese type 2 diabetic rats. Am. J. Physiol. Renal Physiol. 2015, 308, 1386–1397. [Google Scholar] [CrossRef] [PubMed]
- De Leeuw, I.H.; van Gaal, L.; Vanroelen, W. Magnesium and obesity: Effects of treatment on magnesium and other parameters. Magnesium 1987, 6, 40–47. [Google Scholar] [PubMed]
- Dousdampanis, P.; Trigka, K.; Fourtounas, C. Hypomagnesemia, chronic kidney disease and cardiovascular mortality: Pronounced association but unproven causation. Hemodial Int. 2014, 18, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Zieba, A.; Kata, R.; Dudek, D.; Schlegel-Zawadzka, M.; Nowak, G. Serum trace elements in animal models and human depression: Part III. Magnesium. Relationship with copper. Hum. Psychopharmacol. 2000, 15, 631–635. [Google Scholar] [CrossRef] [PubMed]
- Golf, S.W.; Bender, S.; Gruttner, J. On the significance of magnesium in extreme physical stress. Cardiovasc. Drugs Ther. 1998, 12 Suppl. 2, 197–202. [Google Scholar] [CrossRef]
- Schweigel, M.; Martens, H. Magnesium transport in the gastrointestinal tract. Front. Biosci. 2000, 5, D666–D677. [Google Scholar] [CrossRef] [PubMed]
- Graham, L.A.; Caesar, J.J.; Burgen, A.S. Gastrointestinal absorption and excretion of Mg 28 in man. Metabolism 1960, 9, 646–659. [Google Scholar] [PubMed]
- Xi, Q.; Hoenderop, J.G.; Bindels, R.J. Regulation of magnesium reabsorption in DCT. Pflugers Arch. 2009, 458, 89–98. [Google Scholar] [CrossRef] [PubMed]
- Schlingmann, K.P.; Weber, S.; Peters, M.; Niemann Nejsum, L.; Vitzthum, H.; Klingel, K.; Kratz, M.; Haddad, E.; Ristoff, E.; Dinour, D.; et al. Hypomagnesemia with secondary hypocalcemia is caused by mutations in TRPM6, a new member of the TRPM gene family. Nat. Genet. 2002, 31, 166–170. [Google Scholar] [CrossRef] [PubMed]
- Walder, R.Y.; Landau, D.; Meyer, P.; Shalev, H.; Tsolia, M.; Borochowitz, Z.; Boettger, M.B.; Beck, G.E.; Englehardt, R.K.; Carmi, R.; et al. Mutation of TRPM6 causes familial hypomagnesemia with secondary hypocalcemia. Nat. Genet. 2002, 31, 171–174. [Google Scholar] [CrossRef] [PubMed]
- Voets, T.; Nilius, B.; Hoefs, S.; van der Kemp, A.W.; Droogmans, G.; Bindels, R.J.; Hoenderop, J.G. TRPM6 forms the Mg2+ influx channel involved in intestinal and renal Mg2+ absorption. J. Biol. Chem. 2004, 279, 19–25. [Google Scholar] [CrossRef] [PubMed]
- De Baaij, J.H.; Hoenderop, J.G.; Bindels, R.J. Magnesium in man: Implications for health and disease. Physiol. Rev. 2015, 95, 1–46. [Google Scholar] [CrossRef] [PubMed]
- Stuiver, M.; Lainez, S.; Will, C.; Terryn, S.; Gunzel, D.; Debaix, H.; Sommer, K.; Kopplin, K.; Thumfart, J.; Kampik, N.B.; et al. CNNM2, encoding a basolateral protein required for renal Mg2+ handling, is mutated in dominant hypomagnesemia. Am. J. Hum. Genet. 2011, 88, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Goytain, A.; Quamme, G.A. Functional characterization of ACDP2 (ancient conserved domain protein), a divalent metal transporter. Physiol. Genom. 2005, 22, 382–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Groenestege, W.M.; Hoenderop, J.G.; van den Heuvel, L.; Knoers, N.; Bindels, R.J. The epithelial Mg2+ channel transient receptor potential melastatin 6 is regulated by dietary Mg2+ content and estrogens. J. Am. Soc. Nephrol. 2006, 17, 1035–1043. [Google Scholar] [CrossRef] [PubMed]
- Walker, A.F.; Marakis, G.; Christie, S.; Byng, M. Mg citrate found more bioavailable than other Mg preparations in a randomised, double-blind study. Magnes. Res. 2003, 16, 183–191. [Google Scholar] [PubMed]
- Lindberg, J.S.; Zobitz, M.M.; Poindexter, J.R.; Pak, C.Y. Magnesium bioavailability from magnesium citrate and magnesium oxide. J. Am. Coll. Nutr. 1990, 9, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Kuwertz-Broking, E.; Frund, S.; Bulla, M.; Kleta, R.; August, C.; Kisters, K. Familial hypomagnesemia-hypercalciuria in 2 siblings. Clin. Nephrol. 2001, 56, 155–161. [Google Scholar] [PubMed]
- Ikari, A.; Sanada, A.; Okude, C.; Sawada, H.; Yamazaki, Y.; Sugatani, J.; Miwa, M. Up-regulation of TRPM6 transcriptional activity by AP-1 in renal epithelial cells. J. Cell. Physiol. 2010, 222, 481–487. [Google Scholar] [CrossRef] [PubMed]
- Ikari, A.; Okude, C.; Sawada, H.; Yamazaki, Y.; Sugatani, J.; Miwa, M. TRPM6 expression and cell proliferation are up-regulated by phosphorylation of ERK1/2 in renal epithelial cells. Biochem. Biophys. Res. Commun. 2008, 369, 1129–1133. [Google Scholar] [CrossRef] [PubMed]
- Ikari, A.; Okude, C.; Sawada, H.; Takahashi, T.; Sugatani, J.; Miwa, M. Down-regulation of TRPM6-mediated magnesium influx by cyclosporin A. Naunyn Schmiedebergs Arch. Pharmacol. 2008, 377, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Furukawa, C.; Fujii, N.; Manabe, A.; Matsunaga, T.; Endo, S.; Hasegawa, H.; Ito, Y.; Yamaguchi, M.; Yamazaki, Y.; Ikari, A. Up-regulation of transient receptor potential melastatin 6 channel expression by tumor necrosis factor-alpha in the presence of epidermal growth factor receptor tyrosine kinase inhibitor. J. Cell. Physiol. 2017, 232, 2841–2850. [Google Scholar] [CrossRef] [PubMed]
- Ikari, A. Gifu Pharmaceutical University, Gifu, Japan. Unpublished work. 2018. [Google Scholar]
- Ratliff, B.B.; Abdulmahdi, W.; Pawar, R.; Wolin, M.S. Oxidant Mechanisms in Renal Injury and Disease. Antioxid Redox Signal. 2016, 25, 119–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Laurindo, F.R.; Araujo, T.L.; Abrahao, T.B. Nox NADPH oxidases and the endoplasmic reticulum. Antioxid Redox Signal. 2014, 20, 2755–2775. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Andres, O.; Sanchez-Nino, M.D.; Moreno, J.A.; Ruiz-Ortega, M.; Ramos, A.M.; Sanz, A.B.; Ortiz, A. Downregulation of kidney protective factors by inflammation: Role of transcription factors and epigenetic mechanisms. Am. J. Physiol. Renal Physiol. 2016, 311, 1329–1340. [Google Scholar] [CrossRef] [PubMed]
- Baruch, S.B.; Burich, R.L.; Eun, C.K.; King, V.F. Renal metabolism of citrate. Med. Clin. N. Am. 1975, 59, 569–582. [Google Scholar] [CrossRef]
- Unwin, R.J.; Capasso, G.; Shirley, D.G. An overview of divalent cation and citrate handling by the kidney. Nephron. Physiol. 2004, 98, 15–20. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Pajor, A.M. Functional characterization of CitM, the Mg2+-citrate transporter. J. Membr. Biol. 2002, 185, 9–16. [Google Scholar] [CrossRef] [PubMed]
- Stoll, M.L.; Listman, J.A. Nephrolithiasis in a neonate with transient renal wasting of calcium and magnesium. Pediatr. Nephrol. 2002, 17, 386–389. [Google Scholar] [CrossRef] [PubMed]
- Pak, C.Y. Correction of thiazide-induced hypomagnesemia by potassium-magnesium citrate from review of prior trials. Clin. Nephrol. 2000, 54, 271–275. [Google Scholar] [PubMed]
- Nijenhuis, T.; Renkema, K.Y.; Hoenderop, J.G.; Bindels, R.J. Acid-base status determines the renal expression of Ca2+ and Mg2+ transport proteins. J. Am. Soc. Nephrol. 2006, 17, 617–626. [Google Scholar] [CrossRef] [PubMed]
- Dai, L.J.; Friedman, P.A.; Quamme, G.A. Acid-base changes alter Mg2+ uptake in mouse distal convoluted tubule cells. Am. J. Physiol. 1997, 272, 759–766. [Google Scholar] [CrossRef] [PubMed]
- Brown, D.; Wagner, C.A. Molecular mechanisms of acid-base sensing by the kidney. J. Am. Soc. Nephrol. 2012, 23, 774–780. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, F.; Iaccio, A.; Guerra, G.; Montagnani, S.; Ammendola, R. NADPH-oxidase-dependent reactive oxygen species mediate EGFR transactivation by FPRL1 in WKYMVm-stimulated human lung cancer cells. Free Radic. Biol. Med. 2011, 51, 1126–1136. [Google Scholar] [CrossRef] [PubMed]
- Susa, S.; Wakabayashi, I. Extracellular alkalosis activates ERK mitogen-activated protein kinase of vascular smooth muscle cells through NADPH-mediated formation of reactive oxygen species. FEBS Lett. 2003, 554, 399–402. [Google Scholar] [CrossRef] [Green Version]
- Schreck, C.; O’Connor, P.M. NAD(P)H oxidase and renal epithelial ion transport. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2011, 300, R1023–R1029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chabrashvili, T.; Tojo, A.; Onozato, M.L.; Kitiyakara, C.; Quinn, M.T.; Fujita, T.; Welch, W.J.; Wilcox, C.S. Expression and cellular localization of classic NADPH oxidase subunits in the spontaneously hypertensive rat kidney. Hypertension 2002, 39, 269–274. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence |
|---|---|
| Rat TRPM6 | Sense: 5′-CTTCTTGGGATACCAAATCAG-3′ |
| Antisense: 5′-GAAACTTTTCCTAGTGTAGCTG-3′ | |
| Rat TRPM7 | Sense: 5′-AACCAACACTCTGGAAGAGATCA-3′ |
| Antisense: 5′-TCAGTCAAGTTTTCTCCCACAC-3′ | |
| Rat CNNM2 | Sense: 5′-AACACCATCTTCCTCACCAAGT-3′ |
| Antisense: 5′-TCAGCTCTTCCTTAACGAGGTC-3′ | |
| Rat β-actin | Sense: 5′-CCAACCGTGAAAAGATGACC-3′ |
| Antisense: 5′-CCAGAGGCATACAGGGACAG-3′ | |
| Human TRPM6 (hTRPM6) | Sense: 5′-AAAGTTCAATTGGAGTTGACAAGA-3′ |
| Antisense: 5′-AAATTATTCCTTTCAATGGCTGA-3′ | |
| -421S/-616A | Sense: 5′-CTGTGTGCTTTGTGCCACCTC-3′ |
| Antisense: 5′-GAAATGGGGTCTCACTATGTTG-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Takashina, Y.; Manabe, A.; Hasegawa, H.; Matsunaga, T.; Endo, S.; Ikari, A. Sodium Citrate Increases Expression and Flux of Mg2+ Transport Carriers Mediated by Activation of MEK/ERK/c-Fos Pathway in Renal Tubular Epithelial Cells. Nutrients 2018, 10, 1345. https://doi.org/10.3390/nu10101345
Takashina Y, Manabe A, Hasegawa H, Matsunaga T, Endo S, Ikari A. Sodium Citrate Increases Expression and Flux of Mg2+ Transport Carriers Mediated by Activation of MEK/ERK/c-Fos Pathway in Renal Tubular Epithelial Cells. Nutrients. 2018; 10(10):1345. https://doi.org/10.3390/nu10101345
Chicago/Turabian StyleTakashina, Yui, Aya Manabe, Hajime Hasegawa, Toshiyuki Matsunaga, Satoshi Endo, and Akira Ikari. 2018. "Sodium Citrate Increases Expression and Flux of Mg2+ Transport Carriers Mediated by Activation of MEK/ERK/c-Fos Pathway in Renal Tubular Epithelial Cells" Nutrients 10, no. 10: 1345. https://doi.org/10.3390/nu10101345
APA StyleTakashina, Y., Manabe, A., Hasegawa, H., Matsunaga, T., Endo, S., & Ikari, A. (2018). Sodium Citrate Increases Expression and Flux of Mg2+ Transport Carriers Mediated by Activation of MEK/ERK/c-Fos Pathway in Renal Tubular Epithelial Cells. Nutrients, 10(10), 1345. https://doi.org/10.3390/nu10101345