Physiological Translocation of Lactic Acid Bacteria during Pregnancy Contributes to the Composition of the Milk Microbiota in Mice

 , ,
, ,
Abstract
1. Introduction
2. Material and Methods
2.1. Bacterial Strains and Media
2.2. Construction of pMG36e::luxAB and pMG36e::luxABCDE
2.3. Transformation of L. lactis MG1614 and L. salivarius PS2
2.4. Bioluminescence Assays
2.5. In Vivo Translocation Model
2.6. Statistics Analysis: Sample Size Calculation
3. Results
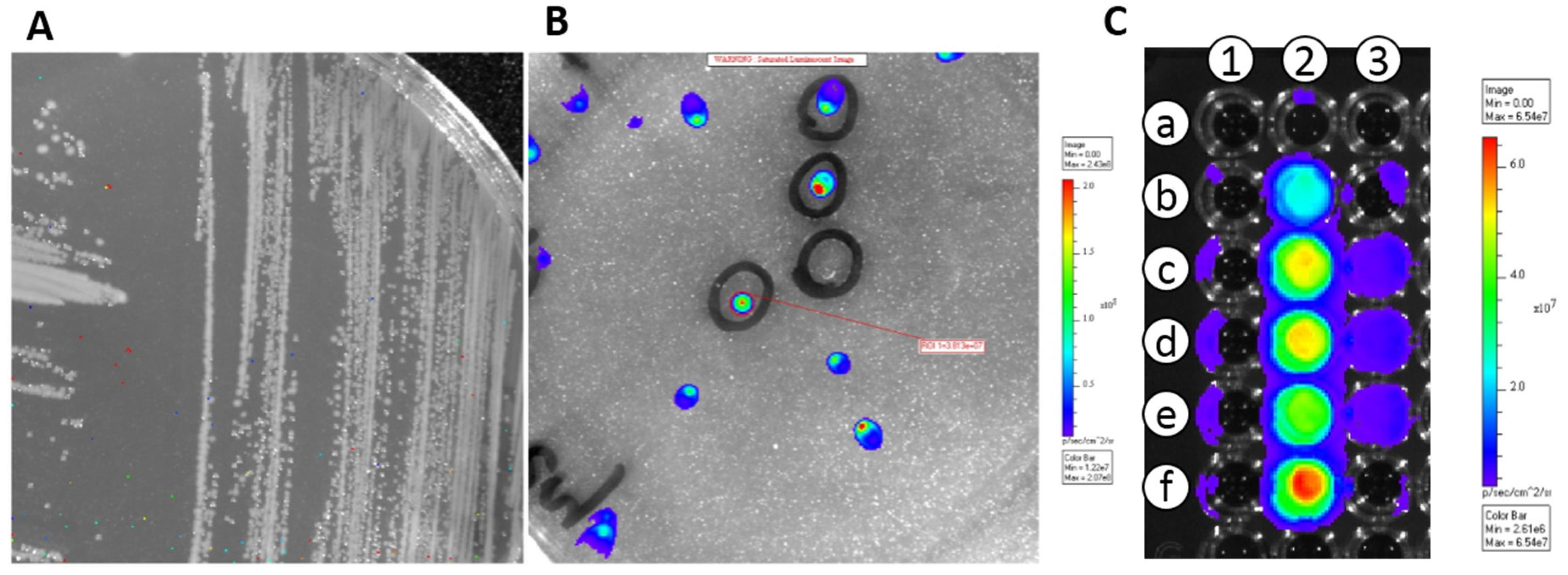
3.1. Transformation of E. coli and LAB Strains with pMG36e::luxAB and pMG36e::luxABCDE
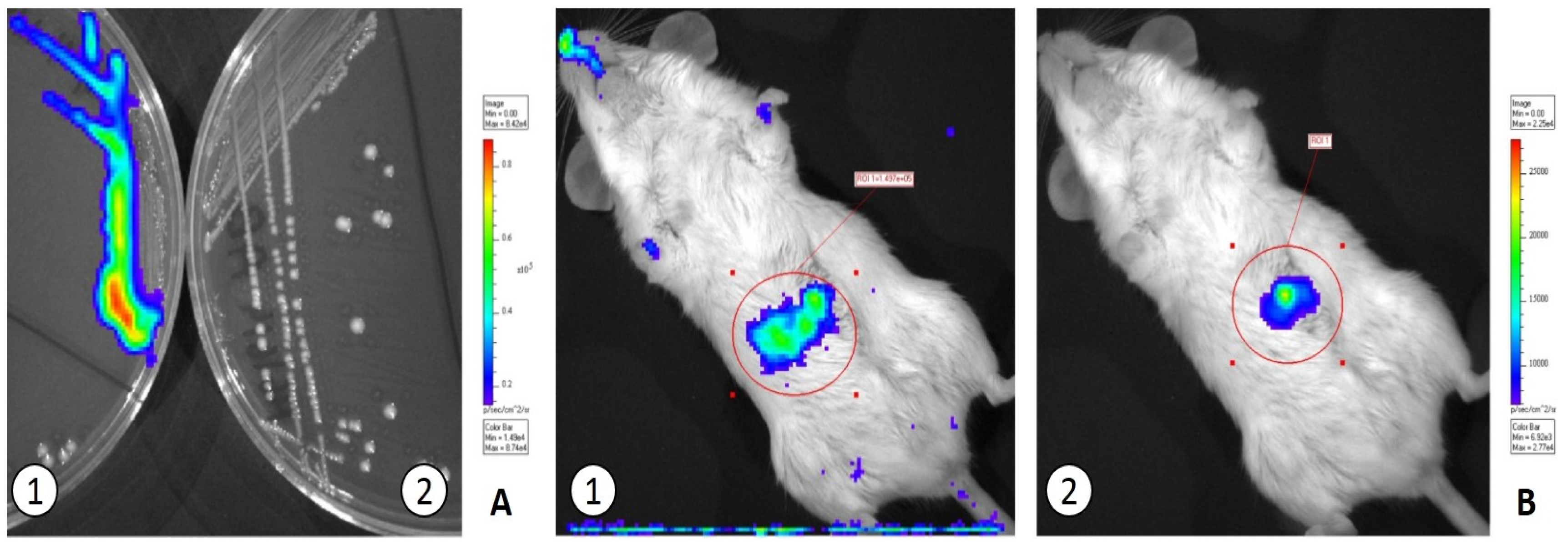
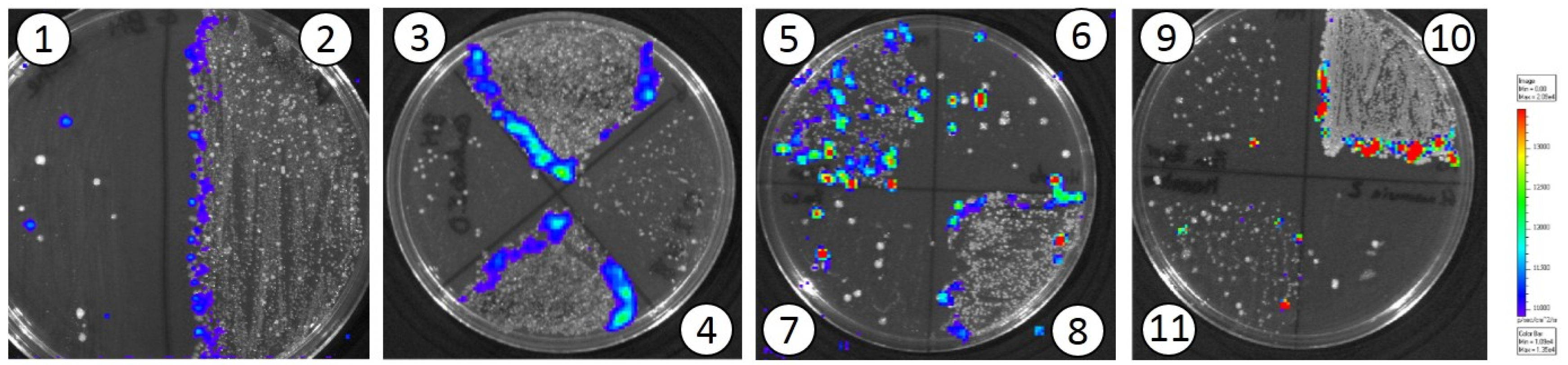
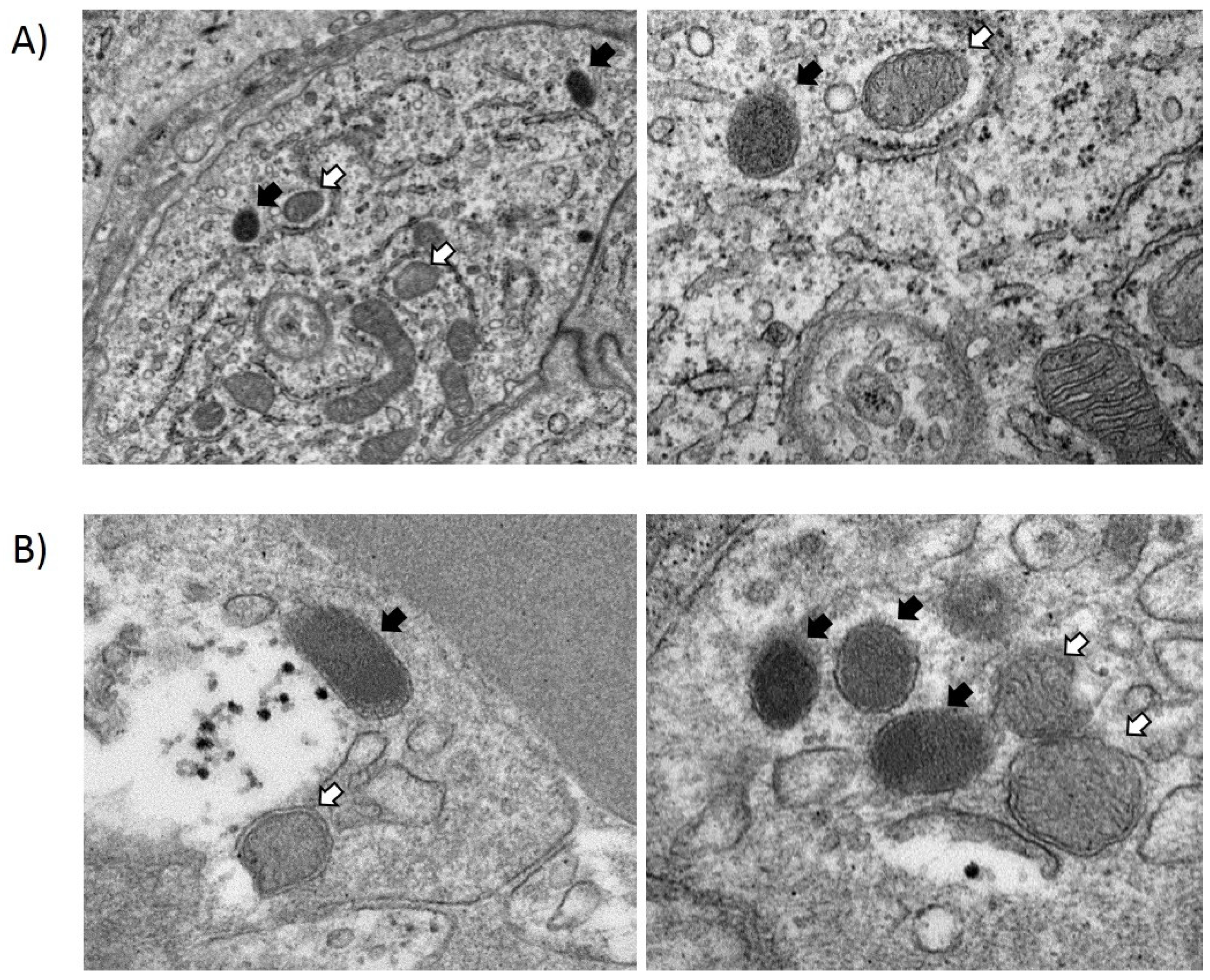
3.2. In Vivo Translocation Model
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Fernandez, L.; Langa, S.; Martin, V.; Maldonado, A.; Jimenez, E.; Martin, R.; Rodriguez, J.M. The Human Milk Microbiota: Origin and Potential Roles in Health and Disease. Pharmacol. Res. 2013, 69, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Jeurink, P.V.; van Bergenhenegouwen, J.; Jimenez, E.; Knippels, L.M.; Fernandez, L.; Garssen, J.; Knol, J.; Rodriguez, J.M.; Martin, R. Human Milk: A Source of More Life than We Imagine. Benef. Microbes 2013, 4, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Martı́n, R.; Langa, S.; Reviriego, C.; Jiménez, E.; Marı́n, M.L.; Olivares, M.; Boza, J.; Jiménez, J.; Fernández, L.; Xaus, J.; et al. The commensal microflora of human milk: New perspectives for food bacteriotherapy and probiotics. Trends Food Sci. Technol. 2004, 15, 121–127. [Google Scholar] [CrossRef]
- Perez, P.F.; Dore, J.; Leclerc, M.; Levenez, F.; Benyacoub, J.; Serrant, P.; Segura-Roggero, I.; Schiffrin, E.J.; Donnet-Hughes, A. Bacterial imprinting of the neonatal immune system: Lessons from maternal cells? Pediatrics 2007, 119, e724–e732. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.; Fernandez, L.; Maldonado, A.; Martin, R.; Olivares, M.; Xaus, J.; Rodriguez, J.M. Oral administration of Lactobacillus strains isolated from breast milk as an alternative for the treatment of infectious mastitis during lactation. Appl. Environ. Microbiol. 2008, 74, 4650–4655. [Google Scholar] [CrossRef] [PubMed]
- Arroyo, R.; Martin, V.; Maldonado, A.; Jimenez, E.; Fernandez, L.; Rodriguez, J.M. Treatment of infectious mastitis during lactation: Antibiotics versus oral administration of lactobacilli isolated from breast milk. Clin. Infect. Dis. 2010, 50, 1551–1558. [Google Scholar] [CrossRef] [PubMed]
- Dunlap, P. Biochemistry and genetics of bacterial bioluminescence. Adv. Biochem. Eng. Biotechnol. 2014, 144, 37–64. [Google Scholar] [PubMed]
- Rocchetta, H.L.; Boylan, C.J.; Foley, J.W.; Iversen, P.W.; LeTourneau, D.L.; McMillian, C.L.; Contag, P.R.; Jenkins, D.E.; Parr, T.R., Jr. Validation of a Noninvasive, Real-Time Imaging Technology using Bioluminescent Escherichia coli in the Neutropenic Mouse Thigh Model of Infection. Antimicrob. Agents Chemother. 2001, 45, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Wiles, S.; Clare, S.; Harker, J.; Huett, A.; Young, D.; Dougan, G.; Frankel, G. Organ specificity, colonization and clearance dynamics in vivo following oral challenges with the murine pathogen citrobacter rodentium. Cell Microbiol. 2004, 6, 963–972. [Google Scholar] [CrossRef] [PubMed]
- Dunn, A.K.; Rader, B.A.; Stabb, E.V.; Mandel, M.J. Regulation of bioluminescence in photobacterium leiognathi strain KNH6. J. Bacteriol. 2015, 197, 3676–3685. [Google Scholar] [CrossRef] [PubMed]
- Diehl, G.E.; Longman, R.S.; Zhang, J.X.; Breart, B.; Galan, C.; Cuesta, A.; Schwab, S.R.; Littman, D.R. Microbiota restricts trafficking of bacteria to mesenteric lymph nodes by CX3CR1Hi cells. Nature 2013, 494, 116–120. [Google Scholar] [CrossRef] [PubMed]
- Cronin, M.; Morrissey, D.; Rajendran, S.; El Mashad, S.M.; van Sinderen, D.; O’Sullivan, G.C.; Tangney, M. Orally administered bifidobacteria as vehicles for delivery of agents to systemic tumors. Mol. Ther. 2010, 18, 1397–1407. [Google Scholar] [CrossRef] [PubMed]
- Gasson, M.J. Plasmid complements of Streptococcus lactis NCDO 712 and other lactic streptococci after protoplast-induced curing. J. Bacteriol. 1983, 154, 1–9. [Google Scholar] [PubMed]
- Vazquez-Fresno, R.; Llorach, R.; Marinic, J.; Tulipani, S.; Garcia-Aloy, M.; Espinosa-Martos, I.; Jimenez, E.; Rodriguez, J.M.; Andres-Lacueva, C. Urinary metabolomic fingerprinting after consumption of a probiotic strain in women with mastitis. Pharmacol. Res. 2014, 87, 160–165. [Google Scholar] [CrossRef] [PubMed]
- Espinosa-Martos, I.; Jimenez, E.; de Andres, J.; Rodriguez-Alcala, L.M.; Tavarez, S.; Manzano, S.; Fernandez, L.; Alonso, E.; Fontecha, J.; Rodriguez, J.M. Milk and blood biomarkers associated to the clinical efficacy of a probiotic for the treatment of infectious mastitis. Benef. Microbes 2016, 7, 305–318. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, L.; Cardenas, N.; Arroyo, R.; Manzano, S.; Jimenez, E.; Martin, V.; Rodriguez, J.M. Prevention of Infectious Mastitis by Oral Administration of Lactobacillus salivarius PS2 during Late Pregnancy. Clin. Infect. Dis. 2016, 62, 568–573. [Google Scholar] [CrossRef] [PubMed]
- Francis, K.P.; Joh, D.; Bellinger-Kawahara, C.; Hawkinson, M.J.; Purchio, T.F.; Contag, P.R. Monitoring Bioluminescent Staphylococcus aureus Infections in Living Mice using a Novel luxABCDE Construct. Infect. Immun. 2000, 68, 3594–3600. [Google Scholar] [CrossRef] [PubMed]
- Van de Guchte, M.; van der Vossen, J.M.; Kok, J.; Venema, G. Construction of a lactococcal expression vector: Expression of hen egg white lysozyme in Lactococcus. lactis subsp. Lactis Appl. Environ. Microbiol. 1989, 55, 224–228. [Google Scholar] [PubMed]
- Holo, H.; Nes, I.F. High-frequency transformation, by electroporation, of Lactococcus. lactis Subsp. cremoris grown with glycine in osmotically stabilized media. Appl. Environ. Microbiol. 1989, 55, 3119–3123. [Google Scholar] [PubMed]
- Mason, C.K.; Collins, M.A.; Thompson, K. Modified electroporation protocol for lactobacilli isolated from the chicken crop facilitates transformation and the use of a genetic tool. J. Microbiol. Methods 2005, 60, 353–363. [Google Scholar] [CrossRef] [PubMed]
- Alvarez-Martin, P.; O’Connell-Motherway, M.; van Sinderen, D.; Mayo, B. Functional analysis of the pBC1 replicon from Bifidobacterium catenulatum L48. Appl. Microbiol. Biotechnol. 2007, 76, 1395–1402. [Google Scholar] [CrossRef] [PubMed]
- Jiménez, E.; Arroyo, R.; Cárdenas, N.; Marín, M.; Serrano, P.; Fernández, L.; Rodríguez, J.M. Mammary candidiasis: A medical condition without scientific evidence? PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [PubMed]
- Faul, F.; Erdfelder, E.; Lang, A.G.; Buchner, A. G*Power 3: A flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behav. Res. Methods 2007, 39, 175–191. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.; Marin, M.L.; Martin, R.; Odriozola, J.M.; Olivares, M.; Xaus, J.; Fernandez, L.; Rodriguez, J.M. Is meconium from healthy newborns actually sterile? Res. Microbiol. 2008, 159, 187–193. [Google Scholar] [CrossRef] [PubMed]
- Greer, L.F., ІІІ; Szalay, A.A. Imaging of light emission from the expression of luciferases in living cells and organisms: A review. Luminescence 2002, 17, 43–74. [Google Scholar] [CrossRef] [PubMed]
- Gahan, C.G. The bacterial lux reporter system: Applications in bacterial localisation studies. Curr. Gene Ther. 2012, 12, 12–19. [Google Scholar] [CrossRef] [PubMed]
- Mediano, P.; Fernandez, L.; Jimenez, E.; Arroyo, R.; Espinosa-Martos, I.; Rodriguez, J.M.; Marin, M. Microbial diversity in milk of women with mastitis: Potential role of coagulase-negative staphylococci, viridans group streptococci, and corynebacteria. J. Hum. Lact. 2017, 33, 309–318. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Heilig, H.G.; Zoetendal, E.G.; Jimenez, E.; Fernandez, L.; Smidt, H.; Rodriguez, J.M. Cultivation-independent assessment of the bacterial diversity of breast milk among healthy women. Res. Microbiol. 2007, 158, 31–37. [Google Scholar] [CrossRef] [PubMed]
- Hunt, K.M.; Foster, J.A.; Forney, L.J.; Schutte, U.M.; Beck, D.L.; Abdo, Z.; Fox, L.K.; Williams, J.E.; McGuire, M.K.; McGuire, M.A. Characterization of the diversity and temporal stability of bacterial communities in human milk. PLoS ONE 2011, 6. [Google Scholar] [CrossRef] [PubMed]
- Cabrera-Rubio, R.; Collado, M.C.; Laitinen, K.; Salminen, S.; Isolauri, E.; Mira, A. The human milk microbiome changes over lactation and is shaped by maternal weight and mode of delivery. Am. J. Clin. Nutr. 2012, 96, 544–551. [Google Scholar] [CrossRef] [PubMed]
- Jost, T.; Lacroix, C.; Braegger, C.P.; Rochat, F.; Chassard, C. Vertical mother-neonate transfer of maternal gut bacteria via breastfeeding. Environ. Microbiol. 2014, 16, 2891–2904. [Google Scholar] [CrossRef] [PubMed]
- Jost, T.; Lacroix, C.; Braegger, C.; Chassard, C. Assessment of bacterial diversity in breast milk using culture-dependent and culture-independent approaches. Br. J. Nutr. 2013, 110, 1253–1262. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.; de Andres, J.; Manrique, M.; Pareja-Tobes, P.; Tobes, R.; Martinez-Blanch, J.F.; Codoner, F.M.; Ramon, D.; Fernandez, L.; Rodriguez, J.M. Metagenomic analysis of milk of healthy and mastitis-suffering women. J. Hum. Lact. 2015, 31, 406–415. [Google Scholar] [CrossRef] [PubMed]
- Albesharat, R.; Ehrmann, M.A.; Korakli, M.; Yazaji, S.; Vogel, R.F. Phenotypic and genotypic analyses of lactic acid bacteria in local fermented food, breast milk and faeces of mothers and their babies. Syst. Appl. Microbiol. 2011, 34, 148–155. [Google Scholar] [CrossRef] [PubMed]
- Martin, V.; Maldonado-Barragan, A.; Moles, L.; Rodriguez-Banos, M.; Campo, R.D.; Fernandez, L.; Rodriguez, J.M.; Jimenez, E. Sharing of Bacterial Strains between Breast Milk and Infant Feces. J. Hum. Lact. 2012, 28, 36–44. [Google Scholar] [CrossRef] [PubMed]
- Milani, C.; Mancabelli, L.; Lugli, G.A.; Duranti, S.; Turroni, F.; Ferrario, C.; Mangifesta, M.; Viappiani, A.; Ferretti, P.; Gorfer, V.; et al. Exploring vertical transmission of bifidobacteria from mother to child. Appl. Environ. Microbiol. 2015, 81, 7078–7087. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.M. The origin of human milk bacteria: Is there a bacterial entero-mammary pathway during late pregnancy and lactation? Adv. Nutr. 2014, 5, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Salter, S.J.; Cox, M.J.; Turek, E.M.; Calus, S.T.; Cookson, W.O.; Moffatt, M.F.; Turner, P.; Parkhill, J.; Loman, N.J.; Walker, A.W. Reagent and laboratory contamination can critically impact sequence-based microbiome analyses. BMC Biol. 2014, 12. [Google Scholar] [CrossRef] [PubMed]
- Makino, H.; Kushiro, A.; Ishikawa, E.; Muylaert, D.; Kubota, H.; Sakai, T.; Oishi, K.; Martin, R.; Ben Amor, K.; Oozeer, R.; et al. Transmission of intestinal bifidobacterium longum subsp. longum strains from mother to infant, determined by multilocus sequencing typing and amplified fragment length polymorphism. Appl. Environ. Microbiol. 2011, 77, 6788–6793. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Mikami, K.; Nishino, R.; Matsuoka, T.; Kimura, M.; Koga, Y. Comparative analysis of the properties of bifidobacterial isolates from fecal samples of mother-infant Pairs. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 653–660. [Google Scholar] [CrossRef] [PubMed]
- Kulagina, E.V.; Shkoporov, A.N.; Kafarskaia, L.I.; Khokhlova, E.V.; Volodin, N.N.; Donskikh, E.E.; Korshunova, O.V.; Efimov, B.A. Molecular genetic study of species and strain variability in bifidobacteria population in intestinal microflora of breastfed infants and their mothers. Bull. Exp. Biol. Med. 2010, 150, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Abrahamsson, T.R.; Sinkiewicz, G.; Jakobsson, T.; Fredrikson, M.; Bjorksten, B. Probiotic lactobacilli in breast milk and infant stool in relation to oral intake during the first year of life. J. Pediatr. Gastroenterol. Nutr. 2009, 49, 349–354. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Jimenez, E.; Olivares, M.; Marin, M.L.; Fernandez, L.; Xaus, J.; Rodriguez, J.M. Lactobacillus salivarius cect 5713, a potential probiotic strain isolated from infant feces and breast milk of a mother-child Pair. Int. J. Food Microbiol. 2006, 112, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Martin, R.; Langa, S.; Reviriego, C.; Jiminez, E.; Marin, M.L.; Xaus, J.; Fernandez, L.; Rodriguez, J.M. Human Milk is a Source of Lactic Acid Bacteria for the Infant Gut. J. Pediatr. 2003, 143, 754–758. [Google Scholar] [CrossRef] [PubMed]
- Mira, A.; Rodríguez, J.M. The origin of human milk bacteria. In Prebiotics and Probiotics in Human Milk; McGuire, M., McGuire, M., Bode, L., Eds.; Academic Press: London, UK, 2017; pp. 349–364. [Google Scholar]
- Begier, E.M.; Barrett, N.L.; Mshar, P.A.; Johnson, D.G.; Hadler, J.L. Connecticut bioterrorism field epidemiology response team. Gram-positive rod surveillance for early anthrax detection. Emerg. Infect. Dis. 2005, 11, 1483–1486. [Google Scholar] [CrossRef] [PubMed]
- Jimenez, E.; Fernandez, L.; Marin, M.L.; Martin, R.; Odriozola, J.M.; Nueno-Palop, C.; Narbad, A.; Olivares, M.; Xaus, J.; Rodriguez, J.M. Isolation of commensal bacteria from umbilical cord blood of healthy neonates born by cesarean section. Curr. Microbiol. 2005, 51, 270–274. [Google Scholar] [CrossRef] [PubMed]
- Vankerckhoven, V.; Van Autgaerden, T.; Huys, G.; Vancanneyt, M.; Swings, J.; Goossens, H. Establishment of the PROSAFE collection of probiotic and human lactic acid bacteria. Microb. Ecol. Health. Dis. 2004, 26, 131–136. [Google Scholar] [CrossRef]
- Ouwehand, A.C.; Isolauri, E.; He, F.; Hashimoto, H.; Benno, Y.; Salminen, S. Differences in Bifidobacterium Flora Composition in Allergic and Healthy Infants. J. Allergy Clin. Immunol. 2001, 108, 144–145. [Google Scholar] [CrossRef] [PubMed]
- Dasanayake, A.P.; Li, Y.; Wiener, H.; Ruby, J.D.; Lee, M.J. Salivary actinomyces naeslundii genospecies 2 and lactobacillus casei levels predict pregnancy outcomes. J. Periodontol. 2005, 76, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Vazquez-Torres, A.; Jones-Carson, J.; Baumler, A.J.; Falkow, S.; Valdivia, R.; Brown, W.; Le, M.; Berggren, R.; Parks, W.T.; Fang, F.C. Extraintestinal dissemination of salmonella by cd18-expressing phagocytes. Nature 1999, 401, 804–808. [Google Scholar] [PubMed]
- Rescigno, M.; Urbano, M.; Valzasina, B.; Francolini, M.; Rotta, G.; Bonasio, R.; Granucci, F.; Kraehenbuhl, J.P.; Ricciardi-Castagnoli, P. Dendritic cells express tight junction proteins and penetrate gut epithelial monolayers to sample bacteria. Nat. Immunol. 2001, 2, 361–367. [Google Scholar] [CrossRef] [PubMed]
- Macpherson, A.J.; Uhr, T. Induction of protective IgA by intestinal dendritic cells carrying commensal bacteria. Science 2004, 303, 1662–1665. [Google Scholar] [CrossRef] [PubMed]
- Roitt, I.M.; Delves, P.J. Essential Immunology, 10th ed.; Blackwell Science: Oxford, UK, 2001. [Google Scholar]
- Bertotto, A.; Gerli, R.; Castellucci, G.; Scalise, F.; Vaccaro, R. Human milk lymphocytes bearing the gamma/delta T-cell receptor are mostly delta TCS1-positive cells. Immunology 1991, 74, 360–361. [Google Scholar] [PubMed]
- Newburg, D.S. Innate Immunity and Human Milk. J. Nutr. 2005, 135, 1308–1312. [Google Scholar] [PubMed]
- Berg, R.D.; Garlington, A.W. Translocation of certain indigenous bacteria from the gastrointestinal tract to the mesenteric lymph nodes and other organs in a gnotobiotic mouse model. Infect. Immun. 1979, 23, 403–411. [Google Scholar] [PubMed]
- Berg, R.D. Bacterial translocation from the gastrointestinal tract. Adv. Exp. Med. Biol. 1999, 473, 11–30. [Google Scholar] [PubMed]
- Balzan, S.; de Almeida Quadros, C.; de Cleva, R.; Zilberstein, B.; Cecconello, I. Bacterial Translocation: Overview of mechanisms and clinical impact. J. Gastroenterol. Hepatol. 2007, 22, 464–471. [Google Scholar] [CrossRef] [PubMed]
- Lichtman, S.M. Bacterial [Correction of Baterial] Translocation in Humans. J. Pediatr. Gastroenterol. Nutr. 2001, 33, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Sedman, P.C.; Macfie, J.; Sagar, P.; Mitchell, C.J.; May, J.; Mancey-Jones, B.; Johnstone, D. The Prevalence of Gut Translocation in Humans. Gastroenterology 1994, 107, 643–649. [Google Scholar] [CrossRef]
- Berg, R.D. Bacterial Translocation from the Gastrointestinal Tract. Trends Microbiol. 1995, 3, 149–154. [Google Scholar] [CrossRef]
- Moore, F.A.; Moore, E.E.; Poggetti, R.S.; Read, R.A. Postinjury shock and early bacteremia. A lethal combination. Arch. Surg. 1992, 127, 897–898. [Google Scholar] [CrossRef]
- Rodriguez, A.V.; Baigori, M.D.; Alvarez, S.; Castro, G.R.; Oliver, G. Phosphatidylinositol-specific phospholipase C activity in lactobacillus rhamnosus with capacity to translocate. FEMS Microbiol. Lett. 2001, 204, 33–38. [Google Scholar] [CrossRef] [PubMed]
- Liong, M.T. Safety of probiotics: Translocation and infection. Nutr. Rev. 2008, 66, 192–202. [Google Scholar] [CrossRef] [PubMed]
- Yamazaki, S.; Machii, K.; Tsuyuki, S.; Momose, H.; Kawashima, T.; Ueda, K. Immunological responses to monoassociated Bifidobacterium longum and their relation to prevention of bacterial invasion. Immunology 1985, 56, 43–50. [Google Scholar] [PubMed]
- Bengmark, S.; Jeppsson, B. Gastrointestinal surface protection and mucosa reconditioning. J. Parenter. Enter. Nutr. 1995, 19, 410–415. [Google Scholar] [CrossRef] [PubMed]
- MacFie, J. Current status of bacterial translocation as a cause of surgical sepsis. Br. Med. Bull. 2004, 71, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Danino, T.; Prindle, A.; Kwong, G.A.; Skalak, M.; Li, H.; Allen, K.; Hasty, J.; Bhatia, S.N. Programmable probiotics for detection of cancer in urine. Sci. Transl. Med. 2015, 7. [Google Scholar] [CrossRef] [PubMed]
- Straka, M. Pregnancy and Periodontal Tissues. Neuro Endocrinol. Lett. 2011, 32, 34–38. [Google Scholar] [PubMed]
- Hammond, K.A. Adaptation of the maternal intestine during lactation. J. Mammary Gland Biol. Neoplasia 1997, 2, 243–252. [Google Scholar] [CrossRef] [PubMed]
- Beischer, N.A.; Mackay, E.V.; Colditz, P.B. Obstetrics and the Newborn; WB Saunders Co.: Philadelphia, PA, USA, 1997. [Google Scholar]
- Li, X.; Fu, G.F.; Fan, Y.R.; Liu, W.H.; Liu, X.J.; Wang, J.J.; Xu, G.X. Bifidobacterium adolescentis as a delivery system of endostatin for cancer gene therapy: Selective inhibitor of angiogenesis and hypoxic tumor growth. Cancer Gene Ther. 2003, 10, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.A.; Shabahang, S.; Timiryasova, T.M.; Zhang, Q.; Beltz, R.; Gentschev, I.; Goebel, W.; Szalay, A.A. Visualization of tumors and metastases in live animals with bacteria and vaccinia virus encoding light-emitting proteins. Nat. Biotechnol. 2004, 22, 313–320. [Google Scholar] [CrossRef] [PubMed]
- Morrissey, D.; O’Sullivan, G.C.; Tangney, M. Tumour targeting with systemically administered bacteria. Curr. Gene Ther. 2010, 10, 3–14. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yang, M.; Li, X.M.; Jiang, P.; Baranov, E.; Li, S.; Xu, M.; Penman, S.; Hoffman, R.M. Tumor-targeting bacterial therapy with amino acid auxotrophs of GFP-expressing Salmonella Typhimurium. Proc. Natl. Acad. Sci. USA 2005, 102, 755–760. [Google Scholar] [CrossRef] [PubMed]
- Zhao, M.; Yang, M.; Ma, H.; Li, X.; Tan, X.; Li, S.; Yang, Z.; Hoffman, R.M. Targeted therapy with a Salmonella Typhimurium leucine-arginine auxotroph cures orthotopic human breast tumors in nude mice. Cancer Res. 2006, 66, 7647–7652. [Google Scholar] [CrossRef] [PubMed]
- Nagakura, C.; Hayashi, K.; Zhao, M.; Yamauchi, K.; Yamamoto, N.; Tsuchiya, H.; Tomita, K.; Bouvet, M.; Hoffman, R.M. Efficacy of a genetically-modified Salmonella Typhimurium in an orthotopic human pancreatic cancer in nude mice. Anticancer. Res. 2009, 29, 1873–1878. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Sequence (5′-3′) | Reference or Source |
|---|---|---|
| XAF1 | CCC CGA GCT CAT GAA GCA AGA GGA GGA CTC TCT ATG | Modified from [17] |
| XBR1 | GGC CCC GGG TTA GGT ATA TTC CAT GTG GTA C | Modified from [17] |
| XAF1 | CCC CGA GCT CAT GAA GCA AGA GGA GGA CTC TCT ATG | Modified from [17] |
| XER2 | GGC GGC GTC GAC TTA ACT ATC AAA CGC TTC GGT TA | Modified from [17] |
| lux1280 | ACG CCG CAG GAA TGT ATT GA | This study |
| lux1732 | TAT GGC GAC AGG ATG ATG AG | This study |
| lux4807 | GTC AAT GAA CGC CGA ATG AG | This study |
| lux5068 | GTC ACT ACT GTC AGG CAC AC | This study |
| Sample | Mice | Growth Medium | |
|---|---|---|---|
| MRS | MRS-Ema | ||
| Feces | 1 | na | 5.92 |
| 2 | 7.96 | 4.72 | |
| 3 | 7.54 | 4.26 | |
| 4 | 7.96 | 3.48 | |
| Small intestine | 1 | 4.80 | 3.00 |
| 2 | 4.64 | nd | |
| 3 | 4.96 | nd | |
| 4 | na | na | |
| Large intestine | 1 | 6.65 | 5.54 |
| 2 | na | na | |
| 3 | 6.48 | 2.70 | |
| 4 | 6.23 | 4.56 | |
| Spleen | 1 | nd | nd |
| 2 | nd | nd | |
| 3 | nd | nd | |
| 4 | 3.65 | 3.90 | |
| Mammary gland tissue | 1 | nd | nd |
| 2 | nd | nd | |
| 3 | 4.98 | 3.30 | |
| 4 | 4.98 | 3.00 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Andrés, J.; Jiménez, E.; Chico-Calero, I.; Fresno, M.; Fernández, L.; Rodríguez, J.M. Physiological Translocation of Lactic Acid Bacteria during Pregnancy Contributes to the Composition of the Milk Microbiota in Mice. Nutrients 2018, 10, 14. https://doi.org/10.3390/nu10010014
De Andrés J, Jiménez E, Chico-Calero I, Fresno M, Fernández L, Rodríguez JM. Physiological Translocation of Lactic Acid Bacteria during Pregnancy Contributes to the Composition of the Milk Microbiota in Mice. Nutrients. 2018; 10(1):14. https://doi.org/10.3390/nu10010014
Chicago/Turabian StyleDe Andrés, Javier, Esther Jiménez, Isabel Chico-Calero, Manuel Fresno, Leónides Fernández, and Juan Miguel Rodríguez. 2018. "Physiological Translocation of Lactic Acid Bacteria during Pregnancy Contributes to the Composition of the Milk Microbiota in Mice" Nutrients 10, no. 1: 14. https://doi.org/10.3390/nu10010014
APA StyleDe Andrés, J., Jiménez, E., Chico-Calero, I., Fresno, M., Fernández, L., & Rodríguez, J. M. (2018). Physiological Translocation of Lactic Acid Bacteria during Pregnancy Contributes to the Composition of the Milk Microbiota in Mice. Nutrients, 10(1), 14. https://doi.org/10.3390/nu10010014