The Amelioration of Olfactory Acuity upon Sexual Maturation Might Affect Food Preferences

Abstract
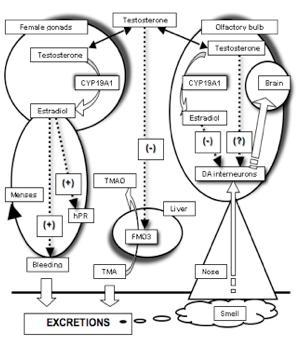
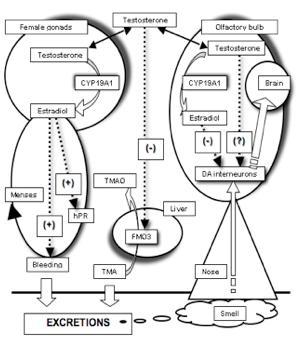
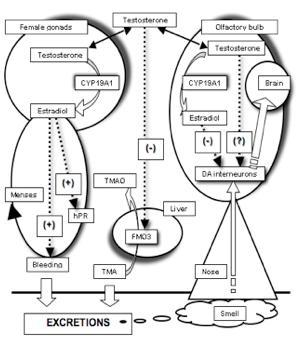
:
1. Introduction
2. Sex Differences in Olfaction
2.1. Olfaction and Menstrual Cycle in Women
2.2. Trimethylamine: An Odor That Might Unveil the Question of Olfactory Sensitivity Changes in Cycling Women
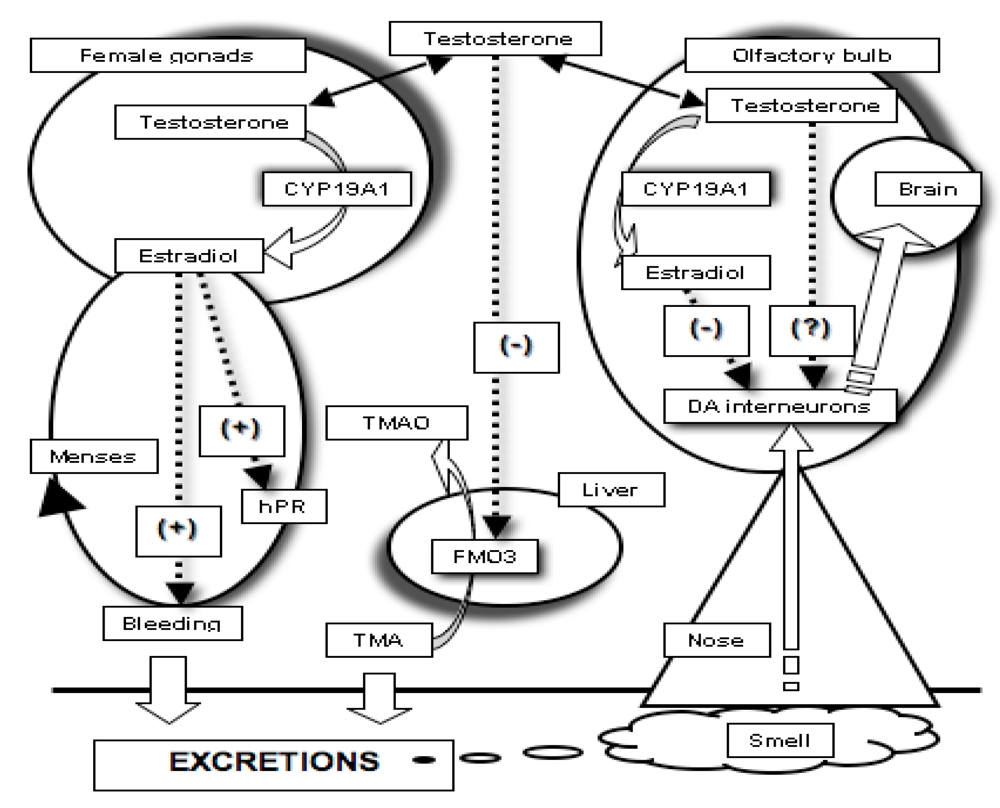
2.3. The Need for a Model Reconciling Several Physiological Aspects under the Control of Testosterone
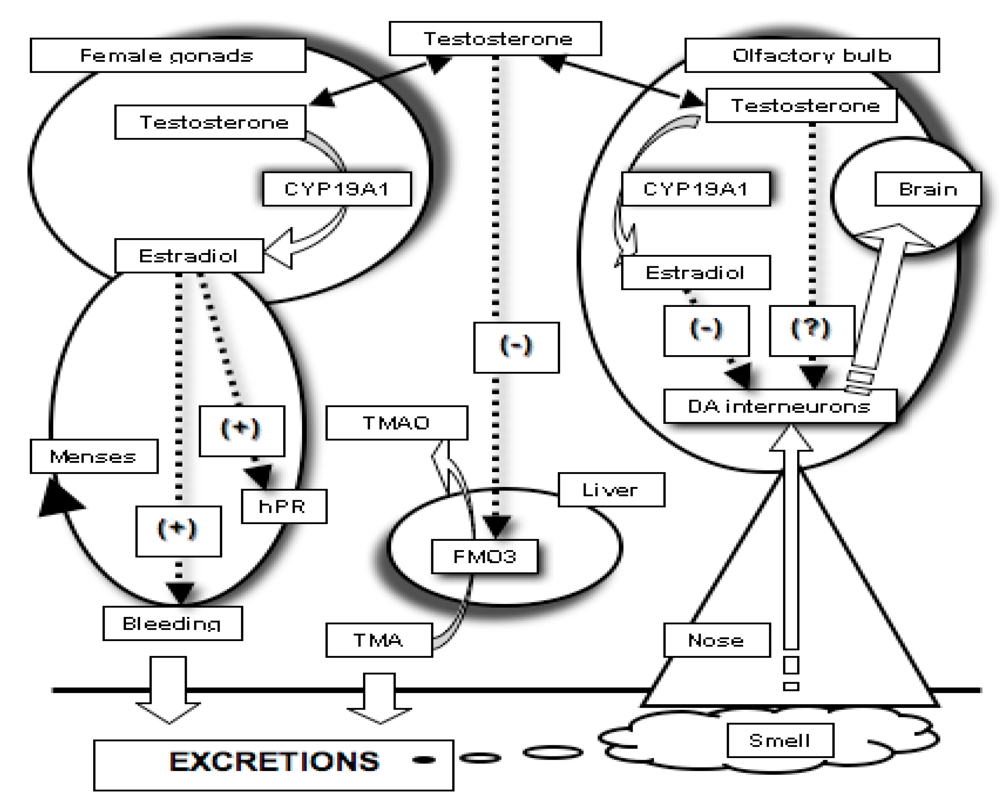
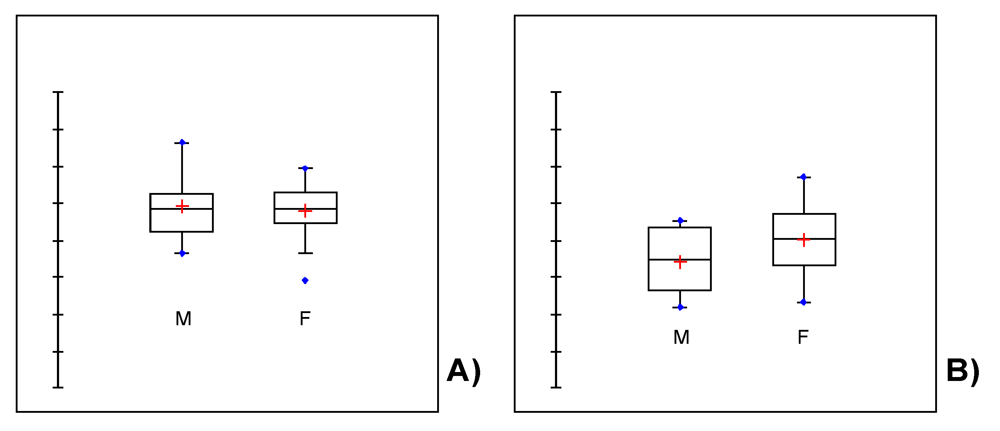
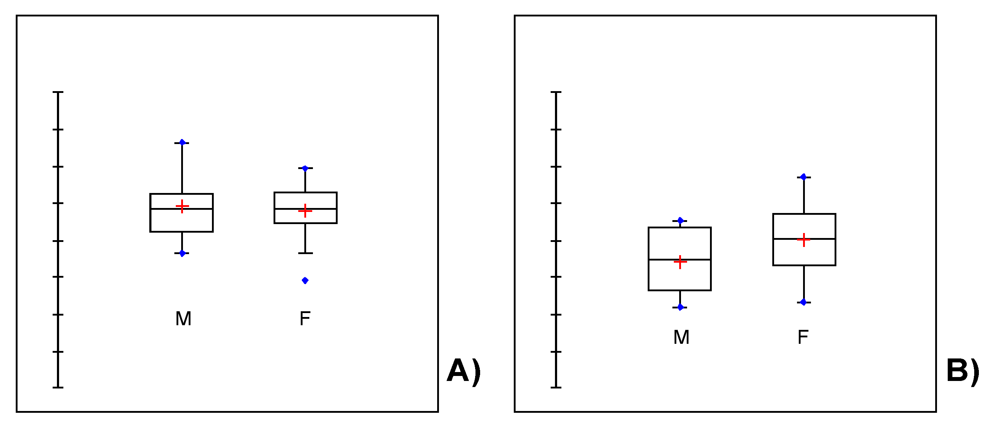
3. Covariation between Sex-Dependent Olfaction and Alimentary Habits

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Sex | Italy | North-West | North-East | North | Centre | South | Islands |
|---|---|---|---|---|---|---|---|---|
| 2007 | M | 59.61 | 56.05 | 49.45 | 53.29 | 62.26 | 66.96 | 64.97 |
| 2007 | F | 60.64 | 57.00 | 51.18 | 54.58 | 62.16 | 67.55 | 67.53 |
| 2007 | M+F | 60.14 | 56.54 | 50.33 | 53.95 | 62.21 | 67.27 | 66.29 |
| Extrapolated from ISTAT (2008). Statistical databases. Available: http://www.istat.it/sanita/Health/ | ||||||||
4. Conclusions
Acknowledgements
References and Notes
- Bignetti, E.; Cavaggioni, A.; Tirindelli, R. The chemistry of chemoreception in vertebrate sensory system. In Sensory Transduction; Borsellino, A., Cervetto, L., Torre, V., Eds.; Plenum Press: New York, NY, USA, 1990; pp. 67–97. [Google Scholar]
- Venstrom, D.; Amoore, J.E. Olfactory threshold in relation to age, sex or smoking. J. Food Sci. 1968, 33, 264–265. [Google Scholar]
- Koelega, H.; Koster, E.P. Some experiments on sex difference in odor perception. Ann. N.Y. Acad. Sci. 1976, 237, 234–246. [Google Scholar]
- Doty, R.L.; Shaman, P.; Michael, D. Development of the university of Pensylvania smell identification test: a standardized microencapsulated test of olfactory function. Physiol. Behav. 1984, 32, 489–502. [Google Scholar]
- Navarrete-Palacios, N.; Hudson, R.; Reyes-Guerrero, G.; Guevara-Guzman, R. Lower olfactory threshold during the ovulatory phase of the menstrual cycle. Biol. Psychol. 2003, 63, 269–279. [Google Scholar]
- Laska, M.; Wieser, A.; Salazar, L.T.H. Sex-specific differences in olfactory sensitivity for putative human pheromones in nonhuman primates. J. Comp. Physiol. 2006, 120, 106–112. [Google Scholar]
- Olsen, S.O. Understanding the relationship between age and seafood consumption: the mediating role of attitude, health involvement and convenience. Food Qual. Prefer. 2003, 14, 199–209. [Google Scholar]
- Bensafi, M.; Brown, W.M.; Tsutsui, T.; Mainland, J.D.; Johnson, B.N.; Bremmer, E.A.; Young, N.; Mauss, I.; Ray, B.; Gross, J.; Richards, J.; Stappen, I.; Levenson, B.; Sobel, N. Sex-steroid derived compounds induce sex-specific effects on autonomic nervous system function in humans. Behav. Neurosci. 2003, 117, 1125–1134. [Google Scholar]
- Bensafi, M.; Brown, W.M.; Khan, R.; Levenson, B.; Sobel, N. Sniffing human sex-steroid compounds modulates mood, memory and autonomic nervous system function in specific behavioral contexts. Behav. Brain Res. 2004, 152, 11–22. [Google Scholar]
- Solbu, E.G.; Jellestad, F.K.; Straetkvern, K.O. Children’s sensitivity to odor of trimethylamine. J. Chem. Ecol. 1989, 16, 1829–1840. [Google Scholar]
- Öberg, C.; Larsson, M.; Bäckman, L. Differential sex effects in olfactory functioning: the role of verbal processing. J. Int. Neuropsych. Soc. 2002, 8, 691–698. [Google Scholar]
- Brand, G.; Millot, J.-L. Sex differences in human olfaction: between evidence and enigma. Q. J. Exp. Psychol. B 2001, 54, 259–270. [Google Scholar]
- Navarrete-Palacios, N.; Hudson, R.; Reyes-Guerrero, G.; Guevara-Guzman, R. Correlation between cytology of the nasal epithelium and the menstrual cycle. Arch. Otolaryngol. 2003, 129, 460–463. [Google Scholar]
- Doty, R.L.; Snyder, P.J.; Huggins, G.R.; Lowry, L.D. Endocrine, cardiovascular and psychological correlates of olfactory sensitivity changes during the human menstrual cycle. J. Comp. Physiol. Psyc. 1981, 95, 45–60. [Google Scholar]
- Pietras, R.J.; Moulton, D.G. Hormonal influences on odor detection in rats: changes associated with the estrous cycle pseudopregnancy, ovariectomy, and administration of testosterone propionate. Physiol. Behav. 1974, 12, 475–491. [Google Scholar] [CrossRef] [PubMed]
- Lamprecht, F.; Matta, R.J.; Little, B.; Zahn, T. Plasma Dopamine-beta-hydroxylase (DBH) Activity During the Menstrual Cycle. Psychosom. Med. 1974, 36, 304–310. [Google Scholar]
- Duchamp-Viret, P.; Coronas, V.; Delaleu, J.-C.; Moyse, E.; Duchamp, A. Dopaminergic modulation of mitral cell activity in the frog olfactory bulb: a combined radioligand binding-electrophysiological study. Neuroscience 1997, 79, 203–216. [Google Scholar]
- Davila, N.; Blakemore, L.J.; Trombley, P.Q. Dopamine modulates synaptic transmission between rat olfactory bulb neurons in culture. J. Neurophysiol. 2003, 90, 395–404. [Google Scholar]
- Davison, I.G.; Boyd, J.D.; Delaney, K.R. Dopamine inhibits Mitral/Tufted→Granule cell synapses in the frog olfactory bulb. J. Neurosci. 2004, 24, 8057–8067. [Google Scholar]
- Pignatelli, A.; Kobayashi, O.H.; Belluzzi, O. Functional properties of dopaminergic neurons in the mouse olfactory bulb. J. Physiol. 2005, 564, 501–514. [Google Scholar]
- Weltzien, F.-A.; Pasqualini, C.; Sebert, M.-E.; Vidal, B.; LeBelle, N.; Kah, O.; Vernier, P.; Dufour, S. Androgen-dependent stimulation of brain dopaminergic system in the female european eel (Anguilla anguilla). Endocrinology 2006, 147, 2964–2973. [Google Scholar]
- Sharma, U.; Dunphy, G.; Ely, D. Testosterone increased blood pressure and decreaded renal tyrosine hydroxylase activity in SHR/Y and Wistar-Kyoto rats. Clin. Exp. Hypertens. 2002, 24, 141–153. [Google Scholar]
- Hafner, H.; Behrens, S.; De Vry, J.; Gattaz, W.F. An animal model for the effects of estradiol on dopamine-mediated behavior: implications for sex differences in schizophrenia. Psychiat. Res. 1991, 38, 125–134. [Google Scholar]
- Kastner, P.; Krust, A.; Turcotte, B.; Stropp, U.; Tora, L.; Gronemeyer, H.; Chambon, P. Two distinct estrogen-regulated promoters generate transcrpts encoding the two functionally different human progesterone receptor forms A and B. EMBO J. 1990, 9, 1603–1614. [Google Scholar] [PubMed]
- Hampton, J.M. C1-C4 Mono, Di-, and Trialkylamine. Spacecraft Water Exposure Guidelines for Selected Contaminants. BEST 2007, 2, 96–153. [Google Scholar]
- Amoore, J.E.; Forrester, L.J. Specific anosmia to trimethylamine: the fishy primary odor. J. Chem. Ecol. 1975, 2, 49–56. [Google Scholar]
- Shewan, J.M.; Hobbs, G.; Hodgkiss, W. The Pseudomonas and Achromobacter groups of bacteria in the spoilage of marine white fish. J. Appl. Microbiol. 1960, 23, 463–468. [Google Scholar]
- Jorgensen, B.R.; Huss, H.H. Growth and activity of Shewanella putrefaciens isolated from spoiling fish. Int. J. Food Microbiol. 1989, 9, 51–62. [Google Scholar]
- Ohashi, E.; Takao, Y.; Fujita, T.; Shimizu, Y.; Egashi, M. Semiconductive Trimethylamine Gas Sensor for Detecting Fish Freshness. J. Food Sci. 1991, 56, 1275–1278. [Google Scholar]
- Caraway, E.A.; Parker, D.; Ruby, M.; Green, G.; Spears, J.; Olsen, M.; Rhoades, M.; Buser, Z. Identification of malodorous compounds from a fish meal plants. In Intl. Symposium on air quality and waste management for agriculture, Bromfield, CO, USA,, 2007; American Society of Agricultural and Biological Engineers (ASABE); 701P0907cd.
- Tver, D.F.; Russell, P. Nutrition and Health Encyclopedia; Van Nostrand Reinhold Press: New York, NY, USA, 1981. [Google Scholar]
- Spiegel, C.A. Bacterial Vaginosis. Clin. Microbiol. Rev. 1991, 4, 485–502. [Google Scholar]
- Humbert, J.R.; Hammond, K.B.; Hathaway, W.E.; Marcoux, J.G.; O’Brien, D. Trimethylaminuria: the fish-odor sindrome. Lancet 1970, 2, 770–771. [Google Scholar]
- Wolrath, H.; Forsum, U.; Larsson, P.G.; Boren, H. Analysis of bacterial vaginosis-related amines in vaginal fluid by gas chromatography and mass spectrometry. J. Clin. Microbiol. 2001, 39, 4026–4031. [Google Scholar]
- Wolrath, H.; Boren, H.; Hallen, A.; Forsum, U. Trimethylamine content in vaginal secretion and its relation to bacterial vaginosis. APMIS 2002, 110, 819–824. [Google Scholar]
- Wolrath, H.; Stahlbom, B.; Hallen, A.; Forsum, U. Trimethylamine and trimethylamine oxide levels in normal women and women with bacterial vaginosis reflect a local metabolism in vaginal secretion as compared to urine. APMIS 2005, 113, 513–516. [Google Scholar]
- Dolphin, C.T.; Janmohamed, A.; Smith, R.L.; Shephard, E.A.; Phillips, L.R. Missense mutation in flavin-containing mono-oxygenase 3 gene, FMO3, underlies fish-odor syndrome. Nat. Genet. 1997, 17, 491–494. [Google Scholar]
- Shimizu, M.; Cashman, J.R.; Yamazaki, H. Transient trimethylaminuria related to menstruation. BMC Med. Genet. 2007, 8, 2–4. [Google Scholar]
- Klaus, K. Beitrag Zur Biochemie der Menstruation. Biochem. Z. 1927, 185, 3–10. [Google Scholar]
- Hukkanen, J.; Dempsey, D.; Jacob, P.; Benowitz, N.L. Effect of pregnancy on a measure of FMO3 activity. Brit. J. Clin. Pharmacol. 2005, 60, 224–226. [Google Scholar]
- El-Alfy, A.T.; Schlenk, D. Effect of 17β-Estradiol and Testosterone on the expression of Flavin-containing Monooxigenase and the toxicity of Aldicarb to Japanese Medaka, Oryzias latipes. Toxicol. Sci. 2002, 68, 381–388. [Google Scholar]
- Keller, M.; Douhard, Q.; Baum, M.J.; Bakker, J. Destruction of the main olfactory epithelium reduces female sexual behaviour and olfactory investigation in female mice. Chem. Senses 2006, 31, 315–323. [Google Scholar]
- Ningdong, K.; Baum, M.J.; James, C. A direct main olfactory bulb projection to the “vomeronasal” amigdala in female mice selectively responds to volatile pheromones from males. Eur. J. Neurosci. 2009, 29, 624–634. [Google Scholar]
- Judd, H.L.; Yen, S.S.C. Serum androstenone and testosterone levels during the menstrual cycle. J. Clin. Endocrinol. Metab. 1973, 36, 475–481. [Google Scholar]
- Epstein, M.T.; McNeilly, A.S.; Murray, M.A.F.; Hockaday, T.D.R. Plasma testosterone and prolactin in the menstrual cycle. Clin. Endocrinol. 1975, 4, 531–535. [Google Scholar]
- Lundstròm, J.N.; McClintock, M.K.; Olsson, M.J. Effects of reproductive state on olfactory sensitivity suggest odor specificity. Biol. Psychol. 2005, 3, 244–247. [Google Scholar]
- Nordin, S.; Brämerson, A.; Lidén, E.; Bende, M. The Scandinavian Odor-Identification Test: development, reliability, validity and normative data. Acta Oto-laryngol. 1998, 118, 226–234. [Google Scholar] [CrossRef]
- Eibenstein, A.; Fioretti, A.B.; Lena, C.; Rosati, N.; Ottaviano, I.; Fusetti, M. Olfactory screening test: experience in 102 italian subjects. Acta Otorhinolaryngol. Ital. 2005, 25, 18–22. [Google Scholar]
- Giusti, A.M.; Bignetti, E.; Cannella, C. Exploring new frontiers in food total quality assessment: from chemical to neurochemical properties. Food Bioprocess Technol. 2007, 2, 1–13. [Google Scholar]
- Dalton, P.; Doolitle, N.; Breslin, P.A.S. Gender-specificd induction of enhanced sensitivity to odors. Nature Neurosci. 2002, 5, 199–200. [Google Scholar]
© 2009 by the authors; licensee Molecular Diversity Preservation International, Basel, Switzerland This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Bignetti, E.; Sinesio, F.; Aiello, G.L.; Cannella, C. The Amelioration of Olfactory Acuity upon Sexual Maturation Might Affect Food Preferences. Nutrients 2009, 1, 3-17. https://doi.org/10.3390/nu1010003
Bignetti E, Sinesio F, Aiello GL, Cannella C. The Amelioration of Olfactory Acuity upon Sexual Maturation Might Affect Food Preferences. Nutrients. 2009; 1(1):3-17. https://doi.org/10.3390/nu1010003
Chicago/Turabian StyleBignetti, Enrico, Fiorella Sinesio, Gaetano L. Aiello, and Carlo Cannella. 2009. "The Amelioration of Olfactory Acuity upon Sexual Maturation Might Affect Food Preferences" Nutrients 1, no. 1: 3-17. https://doi.org/10.3390/nu1010003
APA StyleBignetti, E., Sinesio, F., Aiello, G. L., & Cannella, C. (2009). The Amelioration of Olfactory Acuity upon Sexual Maturation Might Affect Food Preferences. Nutrients, 1(1), 3-17. https://doi.org/10.3390/nu1010003