
Organismic-Scale Remote Sensing of Canopy Foliar Traits in Lowland Tropical Forests


Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area
2.2. Foliar Sampling and Analysis
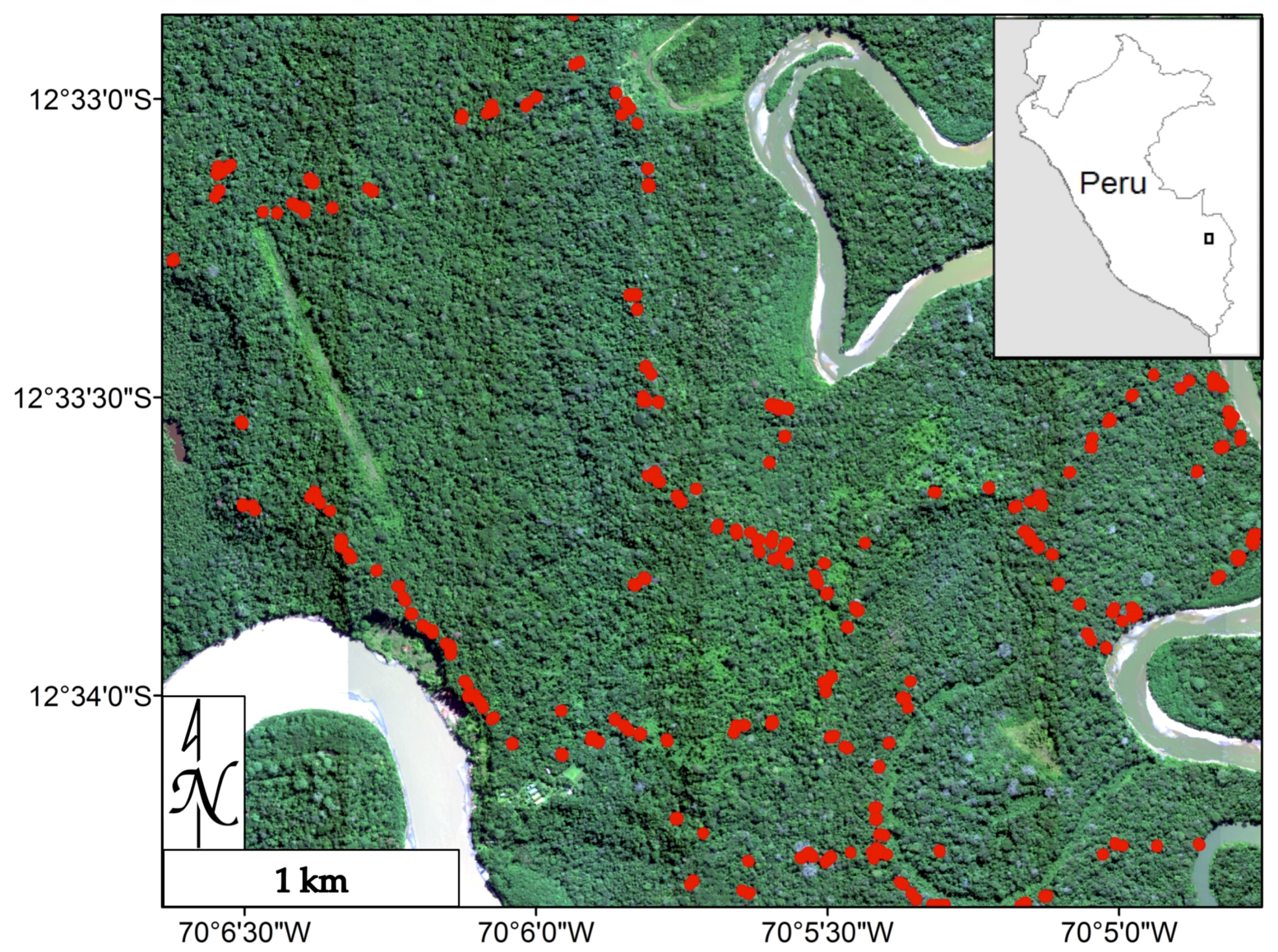
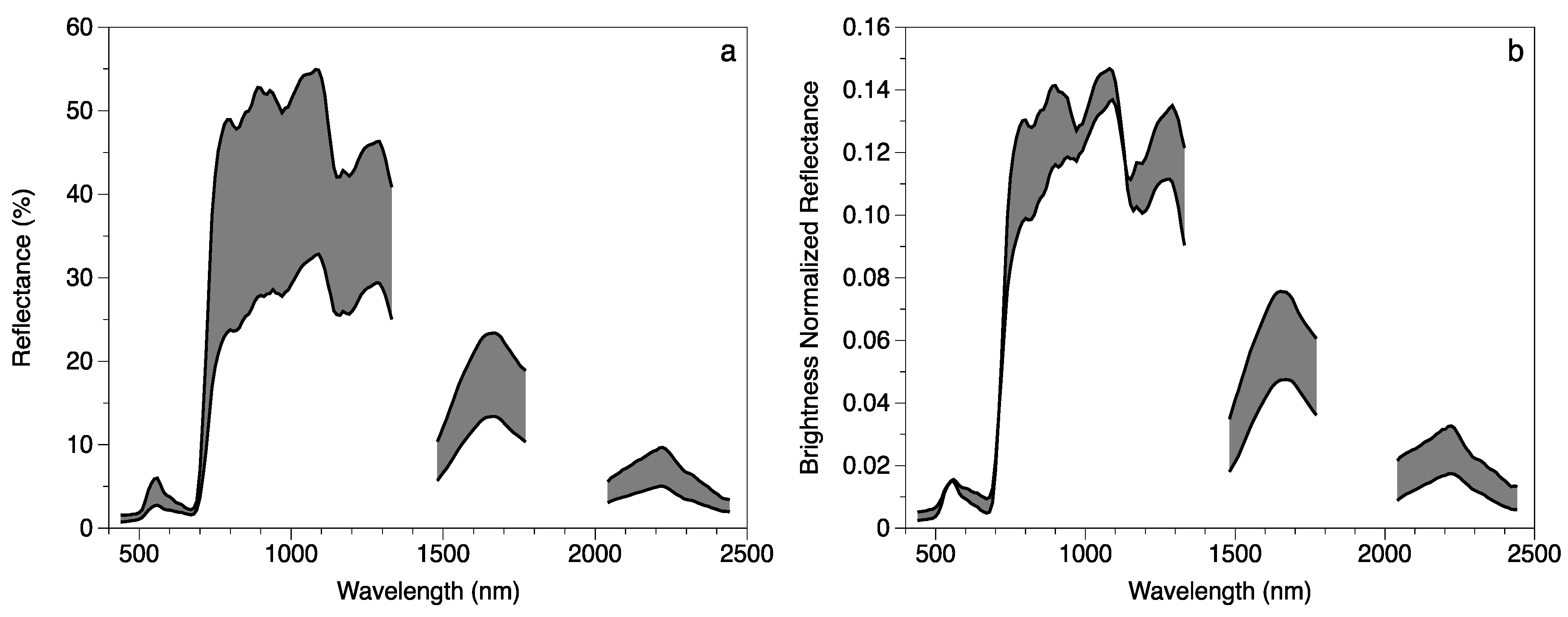
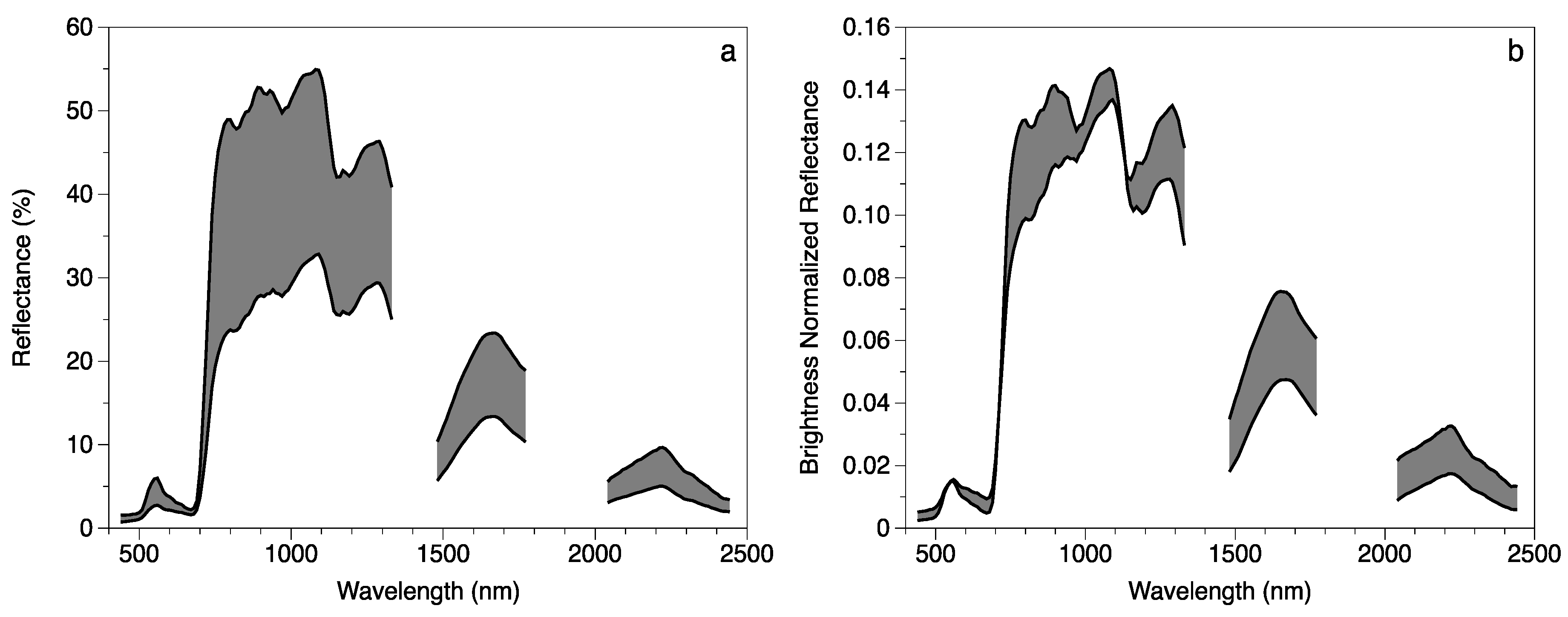
2.3. Remotely Sensed Data
2.4. Model Development
2.5. Model Application and Assessment

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Foliar Trait | Mean | Std. Dev. |
|---|---|---|
| LMA (g·m−2) | 104.4 | 26.6 |
| N (%) | 2.38 | 0.68 |
| P (%) | 0.12 | 0.05 |
| Ca (%) | 0.91 | 0.83 |
| K (%) | 0.64 | 0.31 |
| Mg (%) | 0.22 | 0.13 |
3. Results
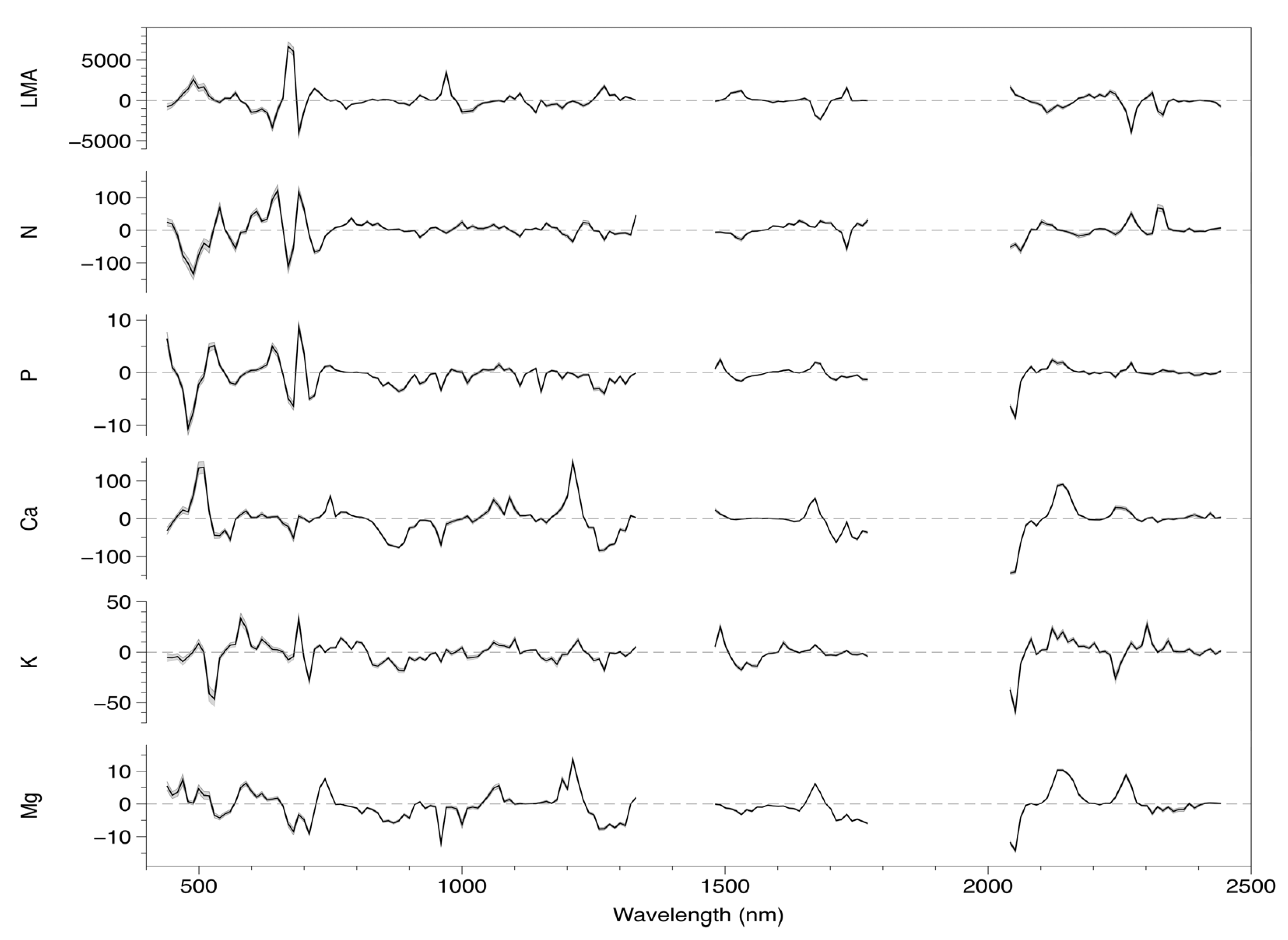
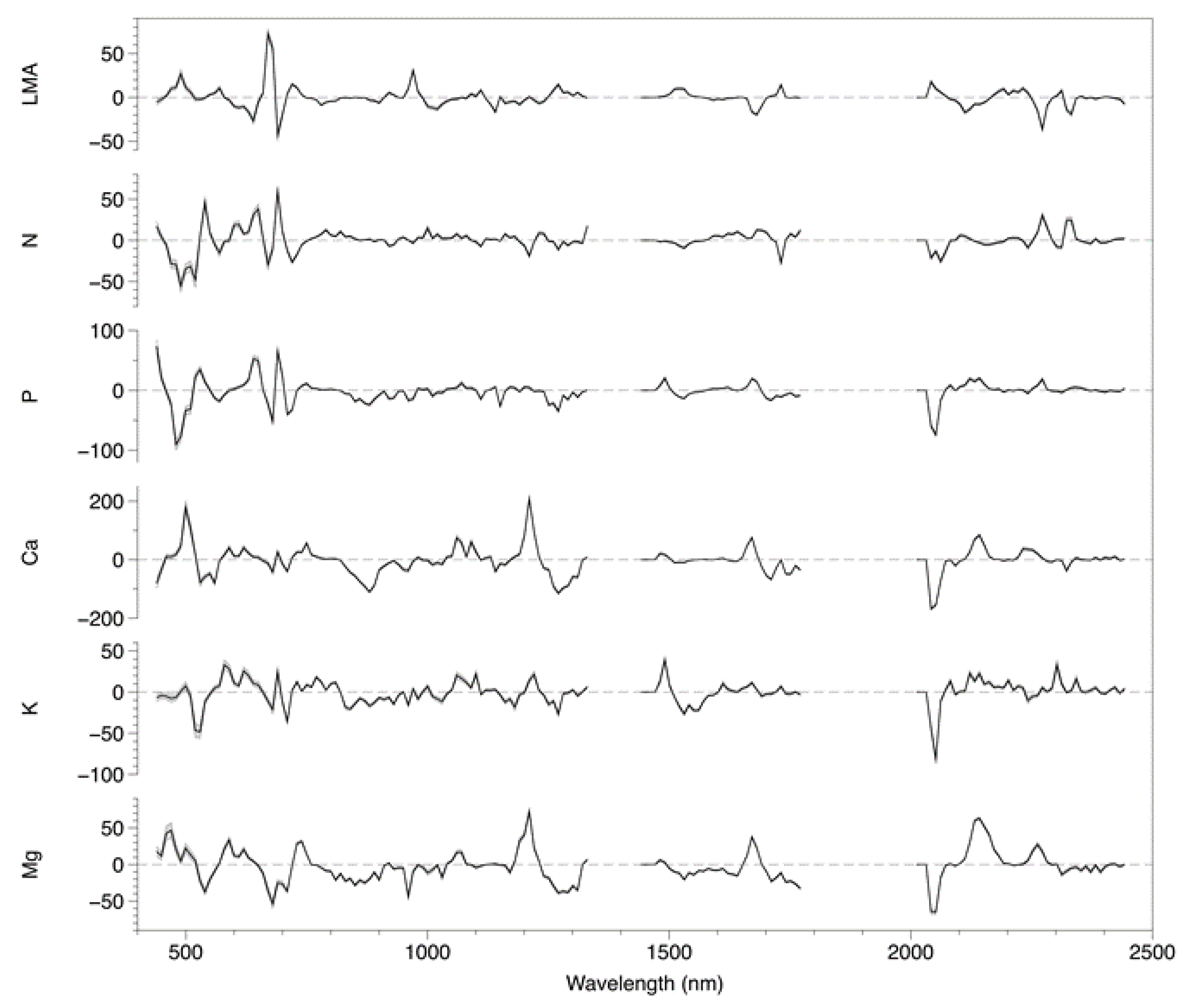
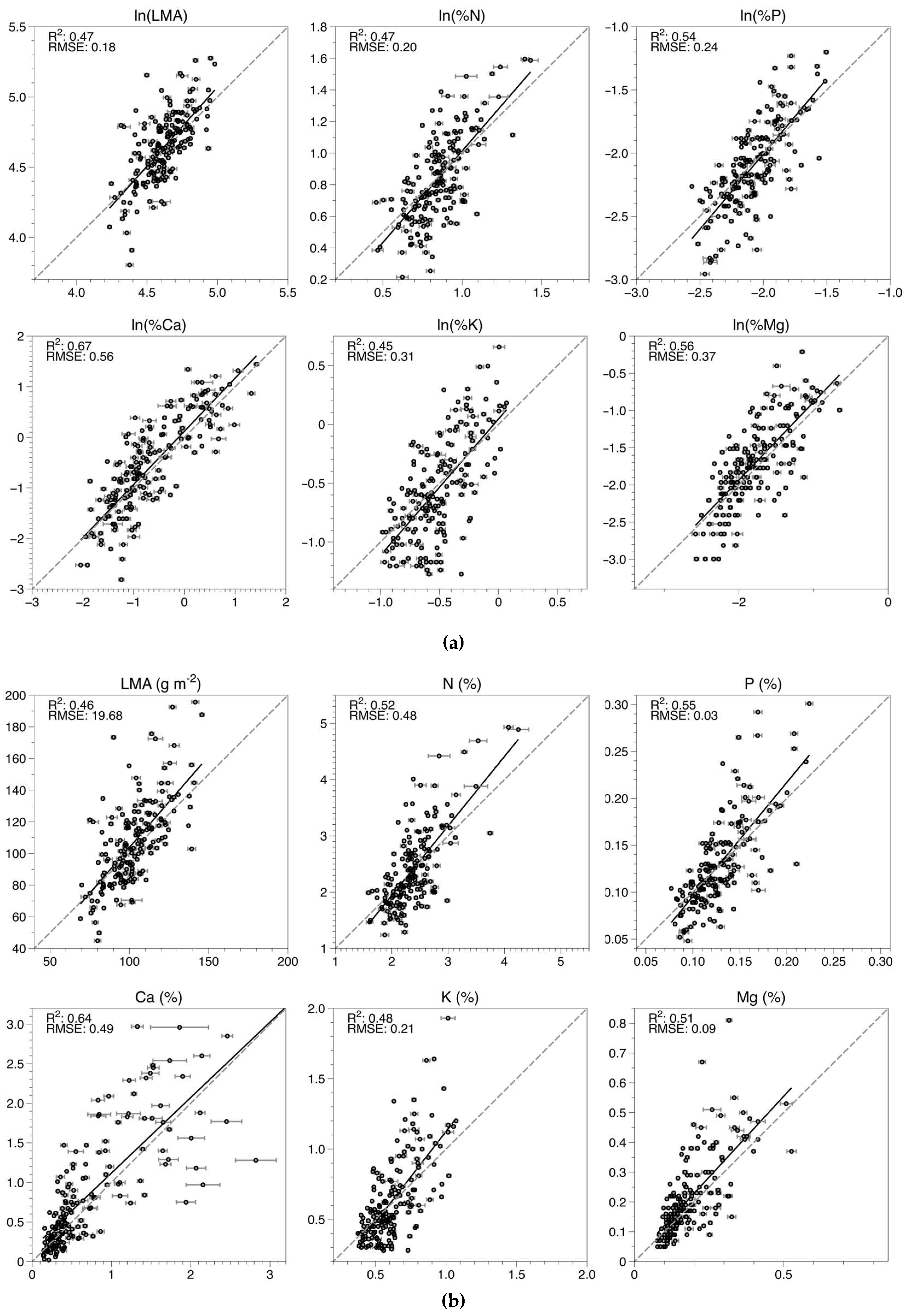
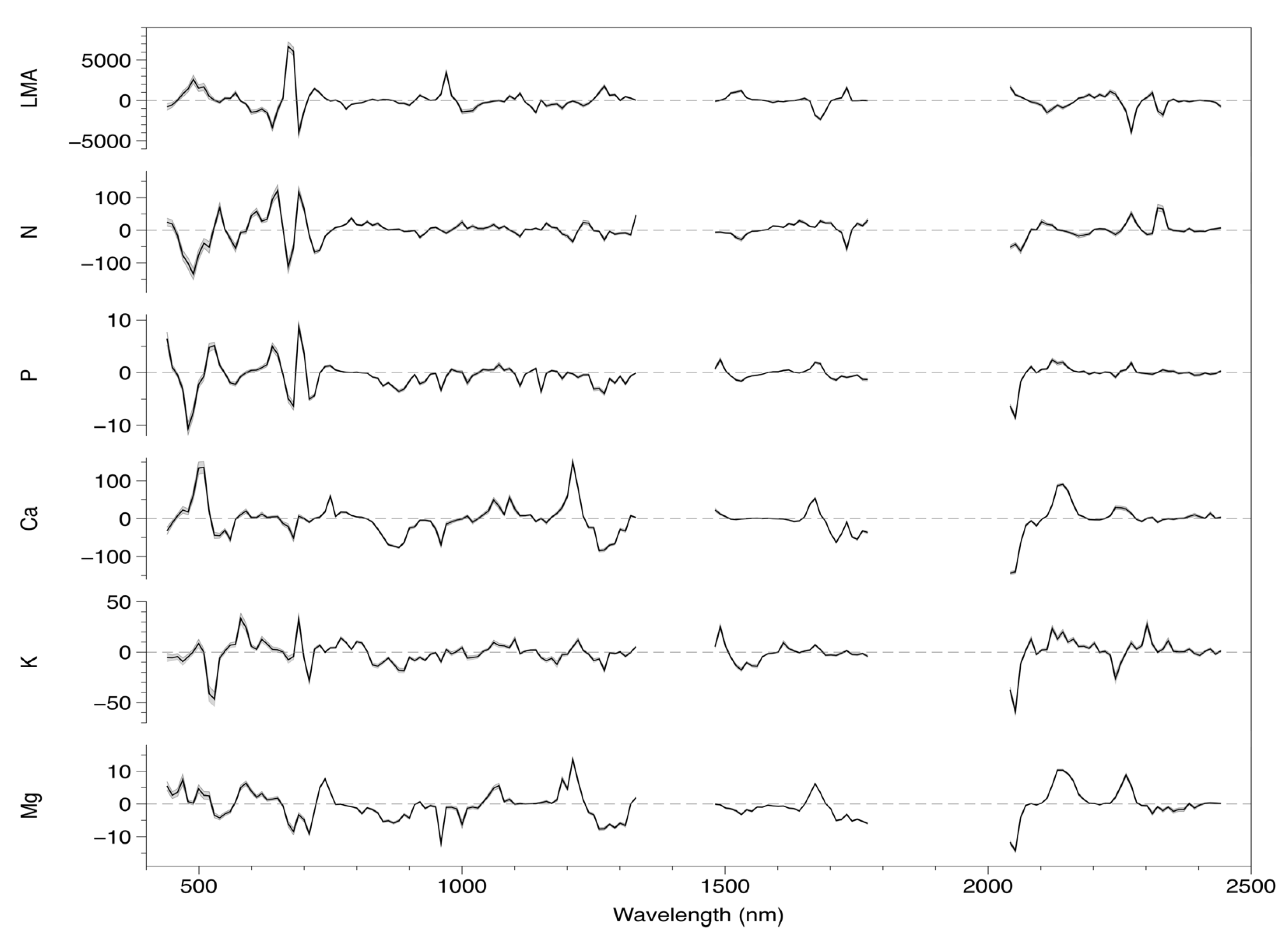
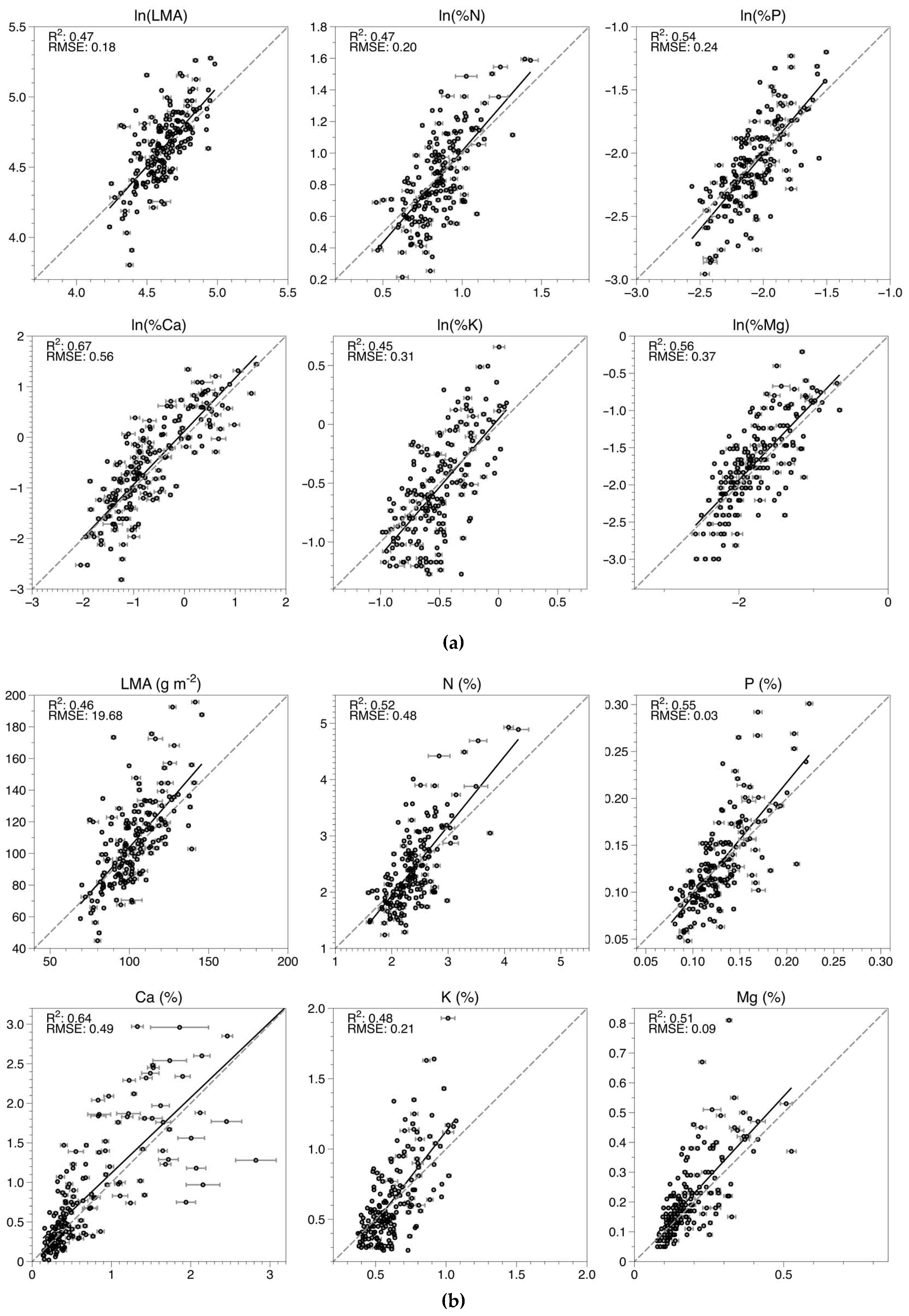
3.1. Model Fitting and Selection
| Foliar Trait | R2 | Cal RMSE | nRMSE | R2 | Val RMSE | nRMSE | LV * | R2 | Test RMSE | nRMSE |
|---|---|---|---|---|---|---|---|---|---|---|
| LMA | 0.46 | 18.69 | 0.12 | 0.43 | 19.25 | 0.12 | 9 | 0.26 | 14.71 | 0.10 |
| N | 0.56 | 0.45 | 0.12 | 0.53 | 0.47 | 0.13 | 9 | 0.36 | 0.38 | 0.10 |
| P | 0.53 | 0.03 | 0.14 | 0.50 | 0.04 | 0.14 | 9 | 0.36 | 0.03 | 0.11 |
| Ca | 0.66 | 0.52 | 0.12 | 0.64 | 0.53 | 0.13 | 9 | 0.56 | 0.47 | 0.11 |
| K | 0.51 | 0.20 | 0.11 | 0.48 | 0.21 | 0.11 | 9 | 0.27 | 0.17 | 0.09 |
| Mg | 0.69 | 0.06 | 0.08 | 0.67 | 0.06 | 0.08 | 9 | 0.57 | 0.06 | 0.08 |
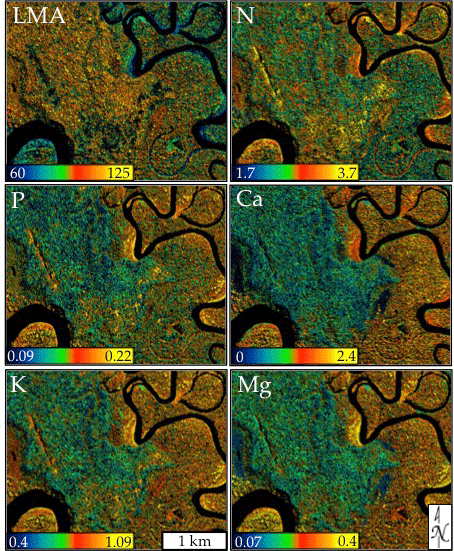
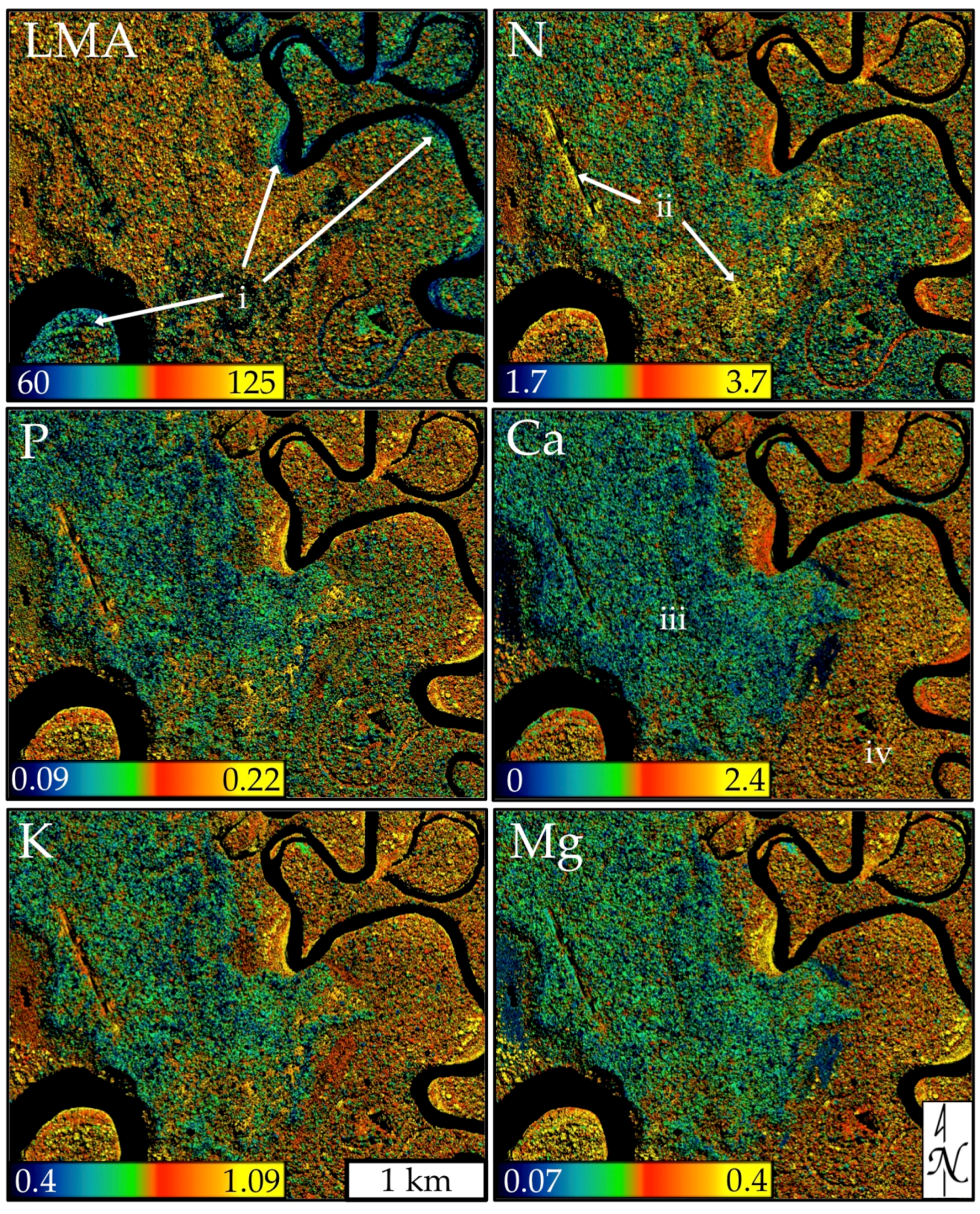
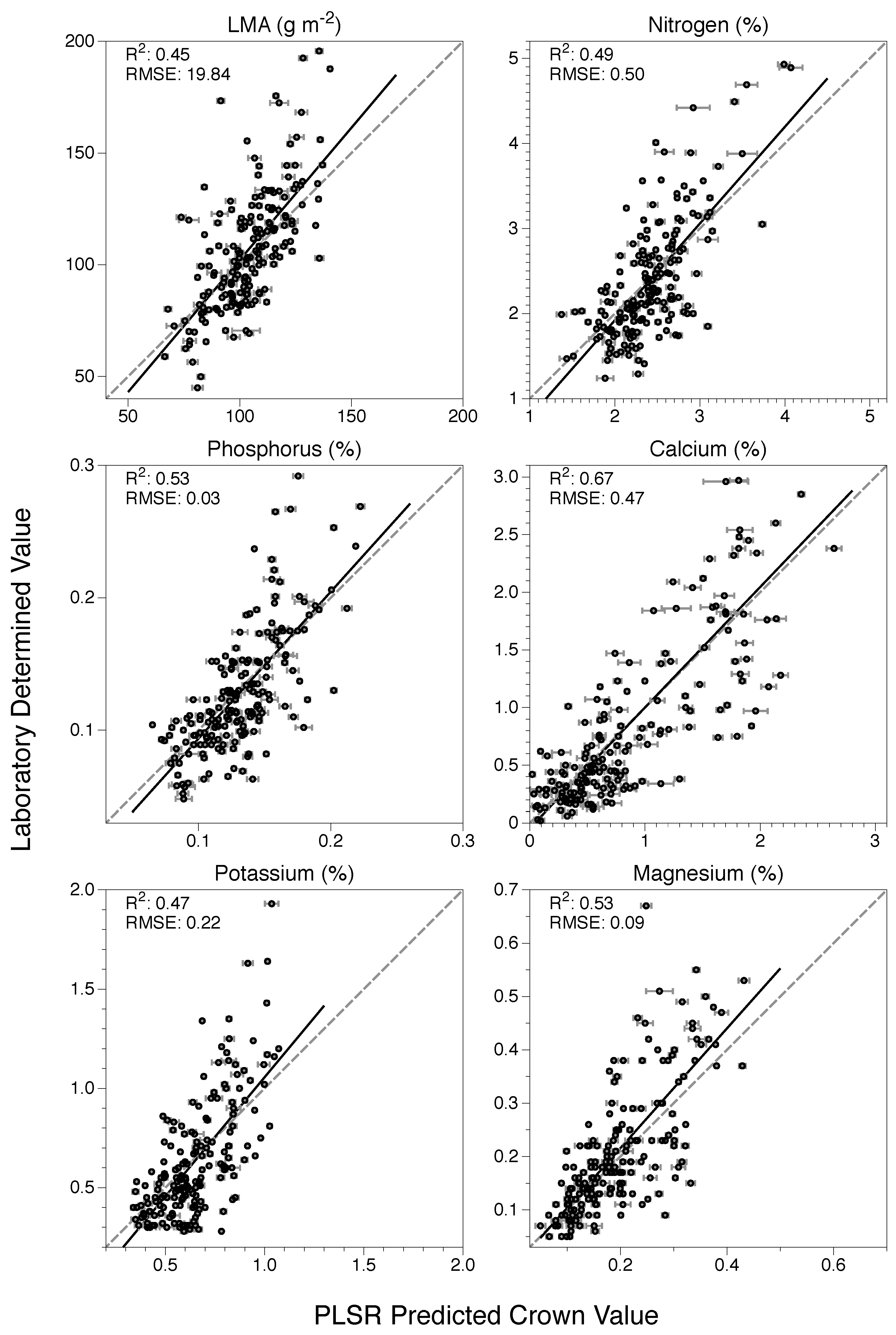
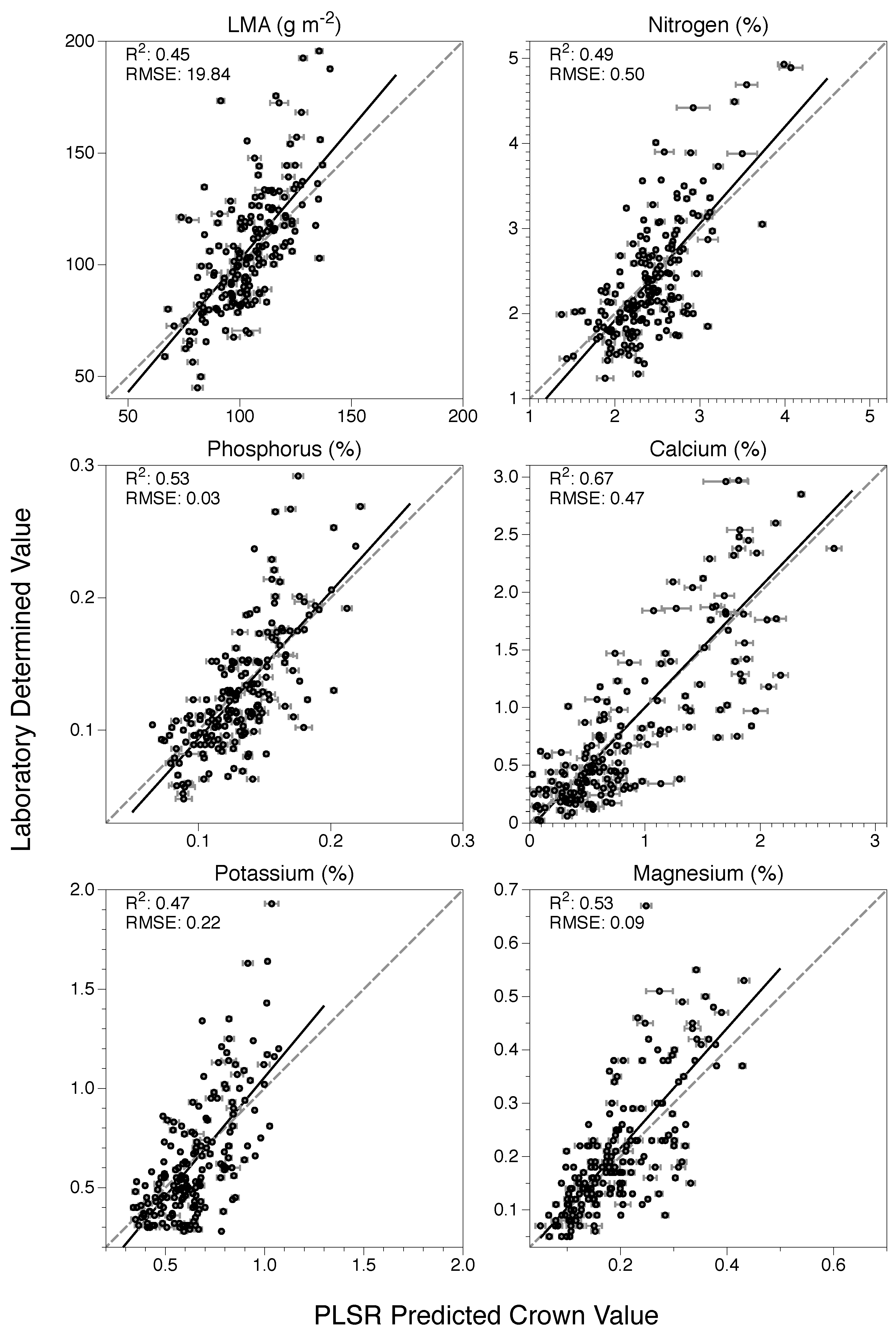
3.2. Model Application and Evaluation
| Trait | Intercept (95% CI) | Slope (95% CI) | R2 | RMSE | nRMSE |
|---|---|---|---|---|---|
| LMA | −16.12 (−36.2, 3.97) | 1.18 (0.99, 1.37) | 0.45 | 19.8 | 0.13 |
| N | −0.35 (−0.77, 0.06) | 1.14 (0.97, 1.31) | 0.49 | 0.50 | 0.14 |
| P | −0.02 (−0.04, 0.003) | 1.11 (0.96, 1.26) | 0.53 | 0.03 | 0.12 |
| Ca | −0.05 (−0.16, 0.07) | 1.05 (0.94, 1.15) | 0.67 | 0.47 | 0.11 |
| K | −0.14 (−0.27, −0.02) | 1.2 (1.02, 1.39) | 0.47 | 0.22 | 0.12 |
| Mg | −0.01 (−0.04, 0.02) | 1.12 (0.97, 1.28) | 0.53 | 0.09 | 0.13 |
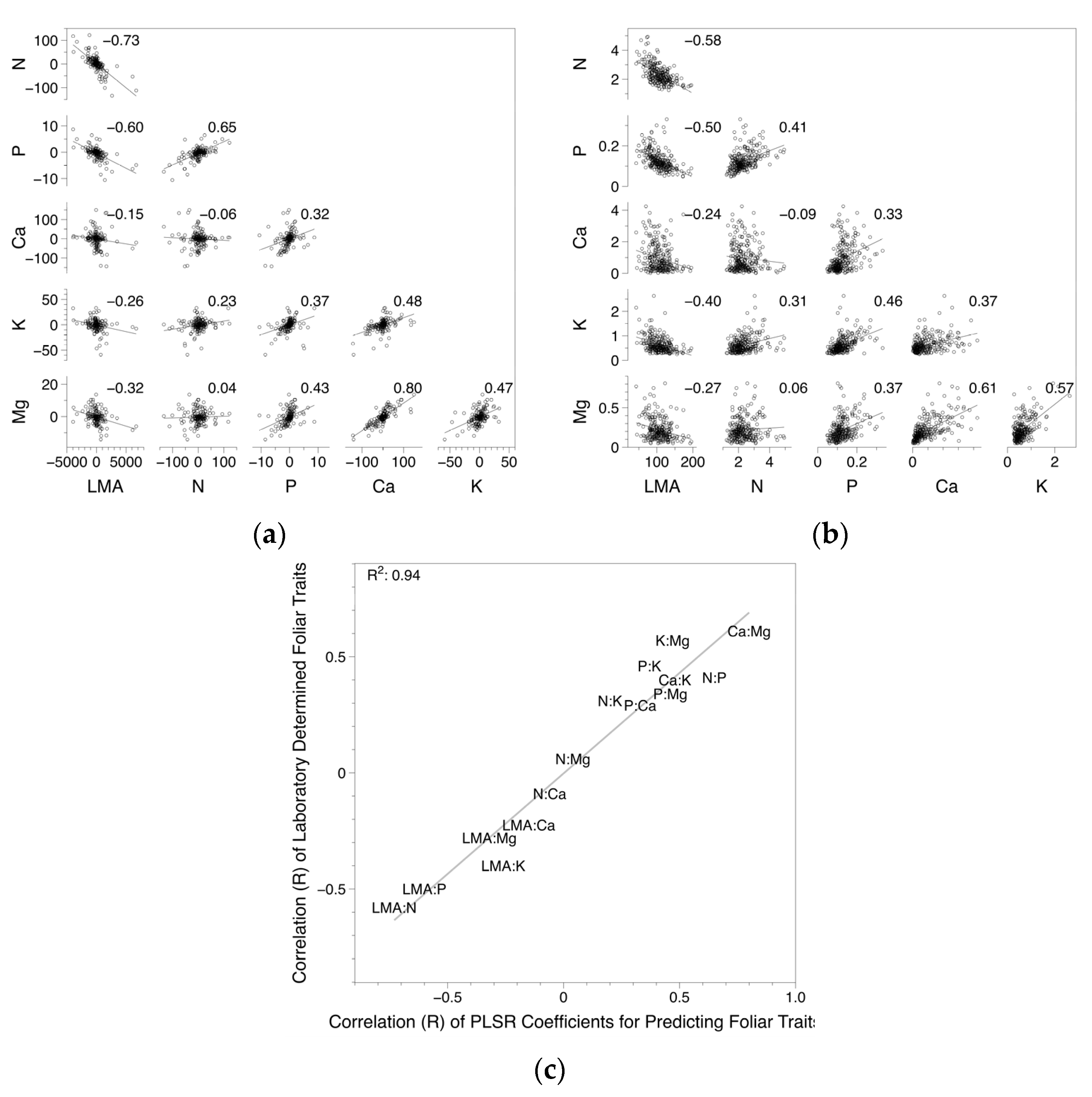
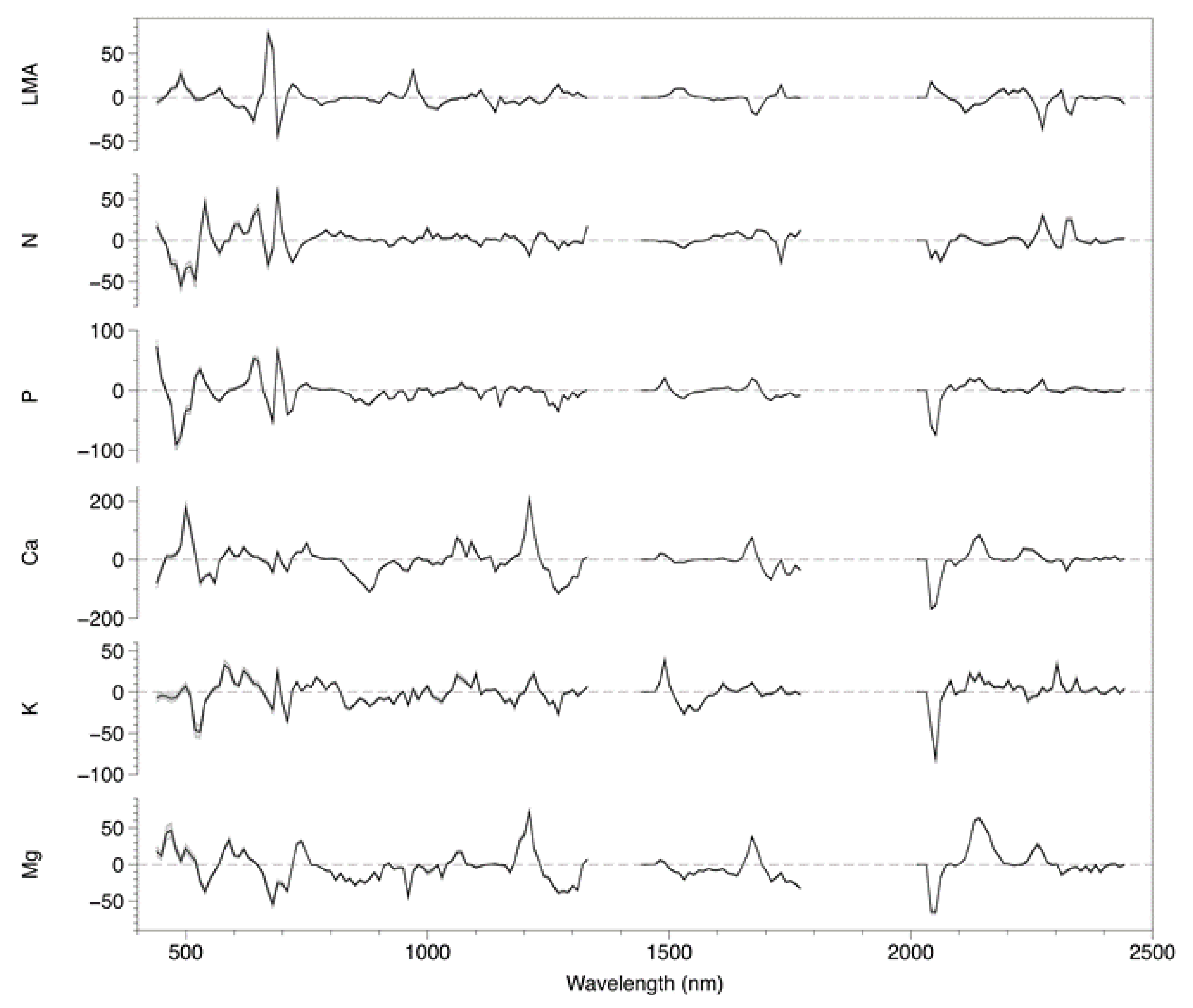
3.3. Coefficient and Trait Correlations

4. Discussion
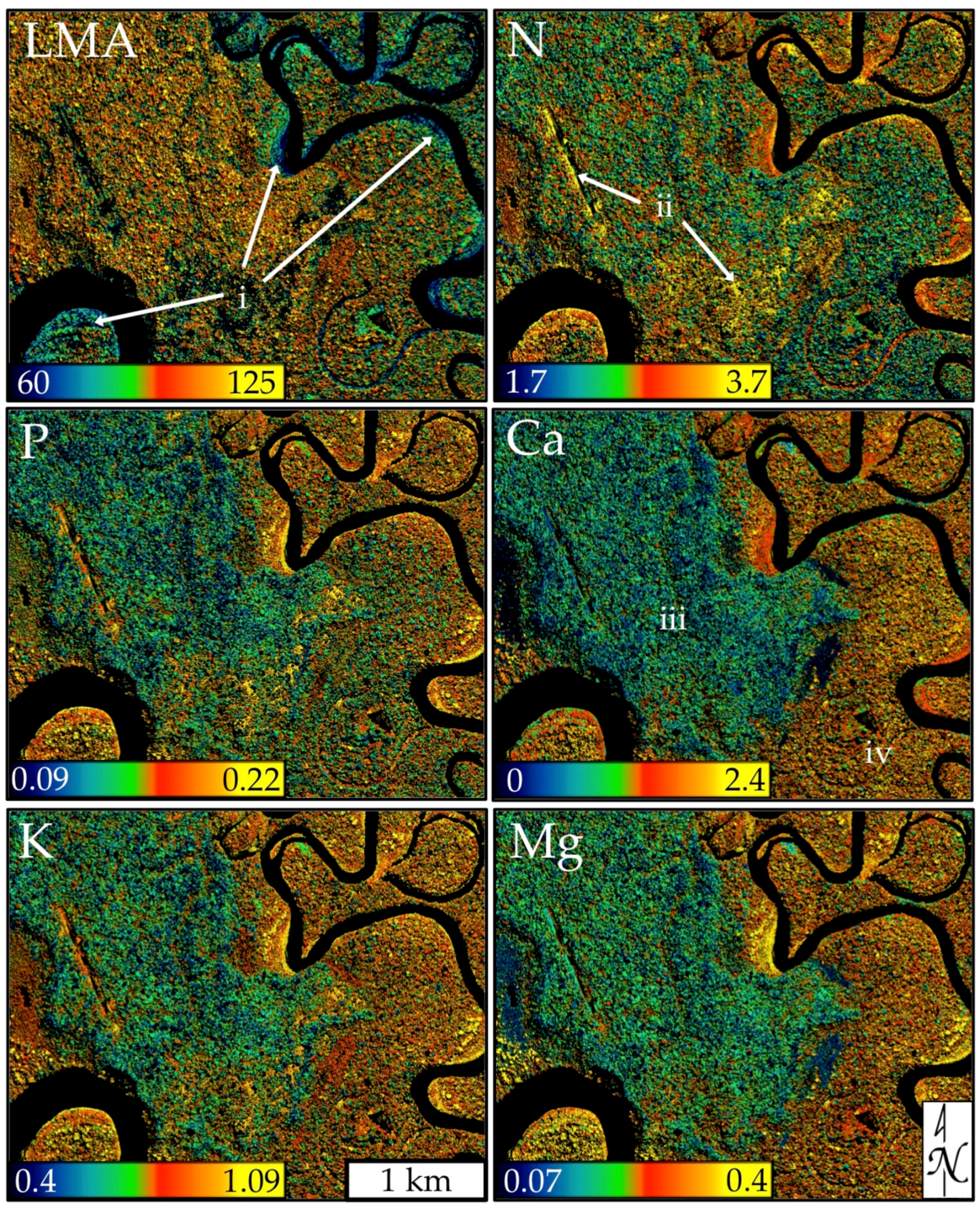
4.1. Mapped Canopy Traits
4.2. Foliar Trait and Canopy Reflectance Interrelationships
4.3. Future Research Directions
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CAO-AToMS | Carnegie Airborne Observatory—Airborne Taxonomic Mapping System |
| DCM | Digital canopy model |
| DSM | Digital surface model |
| DTM | Digital terrain model |
| GPS | Global positioning system |
| HiFIS | High Fidelity Imaging Spectroscopy |
| ICP-OES | inductively coupled plasma optical emission spectrometer |
| IMU | Inertial measurement unit |
| LACC | Los Amigos Conservation Concession |
| LiDAR | Light detection and ranging |
| LMA | Leaf mass per area |
| NDVI | Normalized difference vegetation index |
| PLSR | Partial least squares regression |
| RDN | Rock derived nutrients |
| RMSE | Root mean square error |
| VSWIR | Visible to shortwave infrared |
Appendix A
| Trait | Natural Log Transformed | Back Transformed | ||
|---|---|---|---|---|
| Intercept (95% CI) | Slope (95% CI) | Intercept (95% CI) | Slope (95% CI) | |
| LMA | −0.56 (−1.37, 0.25) | 1.13 (0.95, 1.3) | −11.71 (−30.75, 7.33) | 1.15 (0.97, 1.34) |
| N | −0.14 (−0.3, 0.01) | 1.16 (0.98, 1.34) | −0.53 (−0.95, −0.11) | 1.24 (1.06, 1.41) |
| P | 0.37 (0.04, 0.706) | 1.19 (1.03, 1.35) | −0.03 (−0.05, −0.005) | 1.22 (1.05, 1.38) |
| Ca | 0.12 (0.01, 0.23) | 1.05 (0.94, 1.16) | 0.16 (0.06, 0.27) | 0.95 (0.85, 1.06) |
| K | 0.05 (−0.06, 0.15) | 1.16 (0.97, 1.35) | −0.16 (−0.29, −0.04) | 1.29 (1.09, 1.49) |
| Mg | 0.16 (−0.09, 0.41) | 1.05 (0.91, 1.18) | 0.01 (−0.02, 0.04) | 1.09 (0.93, 1.24) |
References
- Violle, C.; Reich, P.B.; Pacala, S.W.; Enquist, B.J.; Kattge, J. The emergence and promise of functional biogeography. Proc. Natl. Acad. Sci. USA 2014, 111, 13690–13696. [Google Scholar] [CrossRef] [PubMed]
- Malhi, Y.; Grace, J. Tropical forests and atmospheric carbon dioxide. Trends Ecol. Evol. 2000, 15, 332–337. [Google Scholar] [CrossRef]
- Schimel, D.; Stephens, B.B.; Fisher, J.B. Effect of increasing CO2 on the terrestrial carbon cycle. Proc. Natl. Acad. Sci. USA 2015, 112, 436–441. [Google Scholar] [CrossRef] [PubMed]
- Townsend, A.R.; Asner, G.P.; Cleveland, C.C. The biogeochemical heterogeneity of tropical forests. Trends Ecol. Evol. 2008, 23, 424–431. [Google Scholar] [CrossRef] [PubMed]
- Kokaly, R.F.; Asner, G.P.; Ollinger, S.V.; Martin, M.E.; Wessman, C.A. Characterizing canopy biochemistry from imaging spectroscopy and its application to ecosystem studies. Remote Sens. Environ. 2009, 113, S78–S91. [Google Scholar] [CrossRef]
- Ollinger, S.V. Sources of variability in canopy reflectance and the convergent properties of plants. New Phytol. 2011, 189, 375–394. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.P.; Martin, R.E.; Knapp, D.E.; Tupayachi, R.; Anderson, C.; Carranza, L.; Martinez, P.; Houcheime, M.; Sinca, F.; Weiss, P. Spectroscopy of canopy chemicals in humid tropical forests. Remote Sens. Environ. 2011, 115, 3587–3598. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Tupayachi, R.; Emerson, R.; Martinez, P.; Sinca, F.; Powell, G.V.N.; Wright, S.J.; Lugo, A.E. Taxonomy and remote sensing of leaf mass per area (LMA) in humid tropical forests. Ecol. Appl. 2011, 21, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Feilhauer, H.; Asner, G.P.; Martin, R.E.; Schmidtlein, S. Brightness-normalized Partial Least Squares Regression for hyperspectral data. J. Quant. Spectrosc. Radiat. Transf. 2010, 111, 1947–1957. [Google Scholar] [CrossRef]
- Feilhauer, H.; Asner, G.P.; Martin, R.E. Multi-method ensemble selection of spectral bands related to leaf biochemistry. Remote Sens. Environ. 2015, 164, 57–65. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E.; Anderson, C.B.; Knapp, D.E. Quantifying forest canopy traits: Imaging spectroscopy versus field survey. Remote Sens. Environ. 2015, 158, 15–27. [Google Scholar] [CrossRef]
- Asner, G.P.; Anderson, C.B.; Martin, R.E.; Tupayachi, R.; Knapp, D.E.; Sinca, F. Landscape biogeochemistry reflected in shifting distributions of chemical traits in the Amazon forest canopy. Nat. Geosci. 2015, 8, 567–573. [Google Scholar] [CrossRef]
- Reich, P.B.; Walters, M.B.; Ellsworth, D.S. Leaf life-span in relation to leaf, plant, and stand characteristics among diverse ecosystems. Ecol. Monogr. 1992, 62, 365–392. [Google Scholar] [CrossRef]
- Wright, I.J.; Reich, P.B.; Westoby, M.; Ackerly, D.D.; Baruch, Z.; Bongers, F.; Cavender-Bares, J.; Chapin, T.; Cornelissen, J.H.C.; Diemer, M.; et al. The worldwide leaf economics spectrum. Nature 2004, 428, 821–827. [Google Scholar] [CrossRef] [PubMed]
- Demarty, M.; Morvan, C.; Thellier, M. Calcium and the cell wall. Plant Cell Environ. 1984, 7, 441–448. [Google Scholar] [CrossRef]
- McLaughlin, S.B.; Wimmer, R. Calcium physiology and terrestrial ecosystem processes. New Phytol. 1999, 142, 373–417. [Google Scholar] [CrossRef]
- Vitousek, P.M. Litterfall, nutrient cycling, and nutrient limitation in tropical forests. Ecology 1984, 65, 285–298. [Google Scholar] [CrossRef]
- Cuevas, E.; Medina, E. Nutrient dynamics within Amazonian forests. Oecologia 1988, 76, 222–235. [Google Scholar] [CrossRef]
- Kaspari, M.; Garcia, M.; Harms, K. Multiple nutrients limit litterfall and decomposition in a tropical forest. Ecology 2008, 11, 35–43. [Google Scholar] [CrossRef] [PubMed]
- Townsend, A.R.; Cleveland, C.C.; Houlton, B.Z.; Alden, C.B.; White, J.W. Multi-element regulation of the tropical forest carbon cycle. Front. Ecol. Environ. 2011, 9, 9–17. [Google Scholar] [CrossRef]
- Wright, S.J.; Yavitt, J.B.; Wurzburger, N.; Turner, B.L.; Tanner, E.V.J.; Sayer, E.J.; Santiago, L.S.; Kaspari, M.; Hedin, L.O.; Harms, K.E.; et al. Potassium, phosphorus, or nitrogen limit root allocation, tree growth, or litter production in a lowland tropical forest. Ecology 2011, 92, 1616–1625. [Google Scholar] [CrossRef] [PubMed]
- Koerselman, W.; Meuleman, A.F.M. The Vegetation N:P Ratio: A New Tool to Detect the Nature of Nutrient Limitation. J. Appl. Ecol. 1996, 33, 1441–1450. [Google Scholar] [CrossRef]
- Güsewell, S. N:P Ratios in Terrestrial Plants: Variation and Functional Significance. New Phytol. 2004, 164, 243–266. [Google Scholar] [CrossRef]
- Rigsby, C.A.; Hemric, E.M.E.; Baker, P.A. Late Quaternary Paleohydrology of the Madre de Dios River, southwestern Amazon Basin, Peru. Geomorphology 2009, 113, 158–172. [Google Scholar] [CrossRef]
- Osher, L.J.; Buol, S.W. Relationship of soil properties to parent material and landscape position in eastern Madre de Dios, Peru. Geoderma 1998, 83, 143–166. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Airborne spectranomics: Mapping canopy chemical and taxonomic diversity in tropical forests. Front. Ecol. Environ. 2009, 7, 269–276. [Google Scholar] [CrossRef]
- Asner, G.P.; Martin, R.E. Canopy phylogenetic, chemical and spectral assembly in a lowland Amazonian forest. New Phytol. 2011, 189, 999–1012. [Google Scholar] [CrossRef] [PubMed]
- Asner, G.P.; Martin, R.E.; Tupayachi, R.; Anderson, C.B.; Sinca, F.; Carranza-Jimenez, L.; Martinez, P.; Carranza-Jiménez, L.; Martinez, P. Amazonian functional diversity from forest canopy chemical assembly. Proc. Natl. Acad. Sci. USA 2014, 111, 5604–5609. [Google Scholar] [CrossRef] [PubMed]
- Carnegie Spectranomics. Available online: http://spectranomics.ciw.edu (accessed on 19 January 2016).
- Stanford Environmental Measurements Facility. Available online: http://em1.stanford.edu (accessed on 19 January 2016).
- Asner, G.P.; Knapp, D.E.; Boardman, J.; Green, R.O.; Kennedy-Bowdoin, T.; Eastwood, M.; Martin, R.E.; Anderson, C.; Field, C.B. Carnegie Airborne Observatory-2: Increasing science data dimensionality via high-fidelity multi-sensor fusion. Remote Sens. Environ. 2012, 124, 454–465. [Google Scholar] [CrossRef]
- Asner, G.; Martin, R. Spectral and chemical analysis of tropical forests: Scaling from leaf to canopy levels. Remote Sens. Environ. 2008, 112, 3958–3970. [Google Scholar] [CrossRef]
- Asner, G.P.; Knapp, D.E.; Kennedy-Bowdoin, T.; Jones, M.O.; Martin, R.E.; Boardman, J.; Field, C.B. Carnegie Airborne Observatory: In-flight fusion of hyperspectral imaging and waveform light detection and ranging for three-dimensional studies of ecosystems. J. Appl. Remote Sens. 2007, 1. [Google Scholar] [CrossRef]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- RDC Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2012. [Google Scholar]
- Schmidtlein, S.; Feilhauer, H.; Bruelheide, H. Mapping plant strategy types using remote sensing. J. Veg. Sci. 2012, 23, 395–405. [Google Scholar] [CrossRef]
- Piñeiro, G.; Perelman, S.; Guerschman, J.P.; Paruelo, J.M. How to evaluate models: Observed vs. predicted or predicted vs. observed? Ecol. Model. 2008, 216, 316–322. [Google Scholar] [CrossRef]
- Salo, J.S.; Kalliola, R.; Häkkinen, I.; Mäkinen, Y.; Niemelä, P.; Puhakka, M.; Coley, P.D. River dynamics and the diversity of Amazon lowland forest. Nature 1986, 322, 254–258. [Google Scholar] [CrossRef]
- Puhakka, M.; Kalliola, R.; Rajasilta, M.; Salo, J. River Types, Site Evolution and Successional Vegetation Patterns in Peruvian Amazonia. J. Biogeogr. 1992, 19, 651–665. [Google Scholar] [CrossRef]
- Pitman, N.C.A. An Overview of the Los Amigos Watershed, Madre de Dios, Southeastern Peru; Draft Report; Amazon Conservation Association: Washington, DC, USA, 2010. [Google Scholar]
- Chadwick, K.D.; Asner, G.P. Tropical soil nutrient distributions determined by biotic and hillslope processes. Biogeochemistry 2016. [Google Scholar] [CrossRef]
- Fyllas, N.M.; Patiño, S.; Baker, T.R.; Nardoto, G.B.; Martinelli, L.A.; Quesada, C.A.; Paiva, R.; Schwarz, M.; Horna, V.; Mercado, L.M.; et al. Basin-wide variations in foliar properties of Amazonian forest: Phylogeny, soils and climate. Biogeosciences 2009, 6, 2677–2708. [Google Scholar] [CrossRef]
- Townsend, P.A.; Serbin, S.P.; Kruger, E.L.; Gamon, J.A. Disentangling the contribution of biological and physical properties of leaves and canopies in imaging spectroscopy data. Proc. Natl. Acad. Sci. USA 2013, 110. [Google Scholar] [CrossRef] [PubMed]
- Knyazikhin, Y.; Schull, M.A.; Stenberg, P.; Mõttus, M.; Rautiainen, M.; Yang, Y.; Marshak, A.; Latorre Carmona, P.; Kaufmann, R.K.; Lewis, P.; et al. Hyperspectral remote sensing of foliar nitrogen content. Proc. Natl. Acad. Sci. USA 2013, 110. [Google Scholar] [CrossRef] [PubMed]
- Phillips, O.L.; Vargas, P.N.; Lorenzo, A.; Cruz, A.P.; Chuspe, M.; Sánchez, W.G.; Yli-halla, M.; Rose, S.A.M. Habitat association among Amazonian tree species: A landscape-scale approach. J. Ecol. 2003, 91, 757–775. [Google Scholar] [CrossRef]
- Asner, G.P. Organismic Remote Sensing for Tropical Forest Ecology and Conservation. Ann. Mo. Bot. Gard. 2015, 100, 127–140. [Google Scholar] [CrossRef]
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chadwick, K.D.; Asner, G.P. Organismic-Scale Remote Sensing of Canopy Foliar Traits in Lowland Tropical Forests. Remote Sens. 2016, 8, 87. https://doi.org/10.3390/rs8020087
Chadwick KD, Asner GP. Organismic-Scale Remote Sensing of Canopy Foliar Traits in Lowland Tropical Forests. Remote Sensing. 2016; 8(2):87. https://doi.org/10.3390/rs8020087
Chicago/Turabian StyleChadwick, K. Dana, and Gregory P. Asner. 2016. "Organismic-Scale Remote Sensing of Canopy Foliar Traits in Lowland Tropical Forests" Remote Sensing 8, no. 2: 87. https://doi.org/10.3390/rs8020087
APA StyleChadwick, K. D., & Asner, G. P. (2016). Organismic-Scale Remote Sensing of Canopy Foliar Traits in Lowland Tropical Forests. Remote Sensing, 8(2), 87. https://doi.org/10.3390/rs8020087