Use of Landsat and SRTM Data to Detect Broad-Scale Biodiversity Patterns in Northwestern Amazonia

 , and
, and
Abstract
:
1. Introduction
2. Materials and Methods
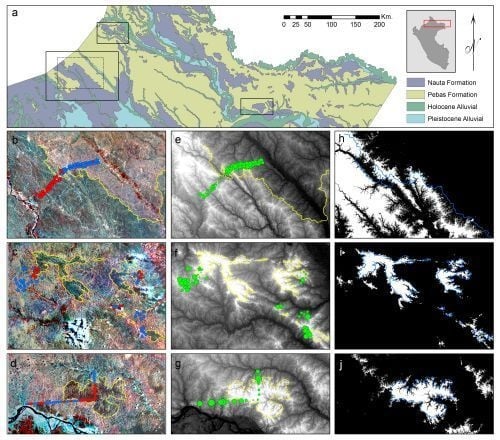
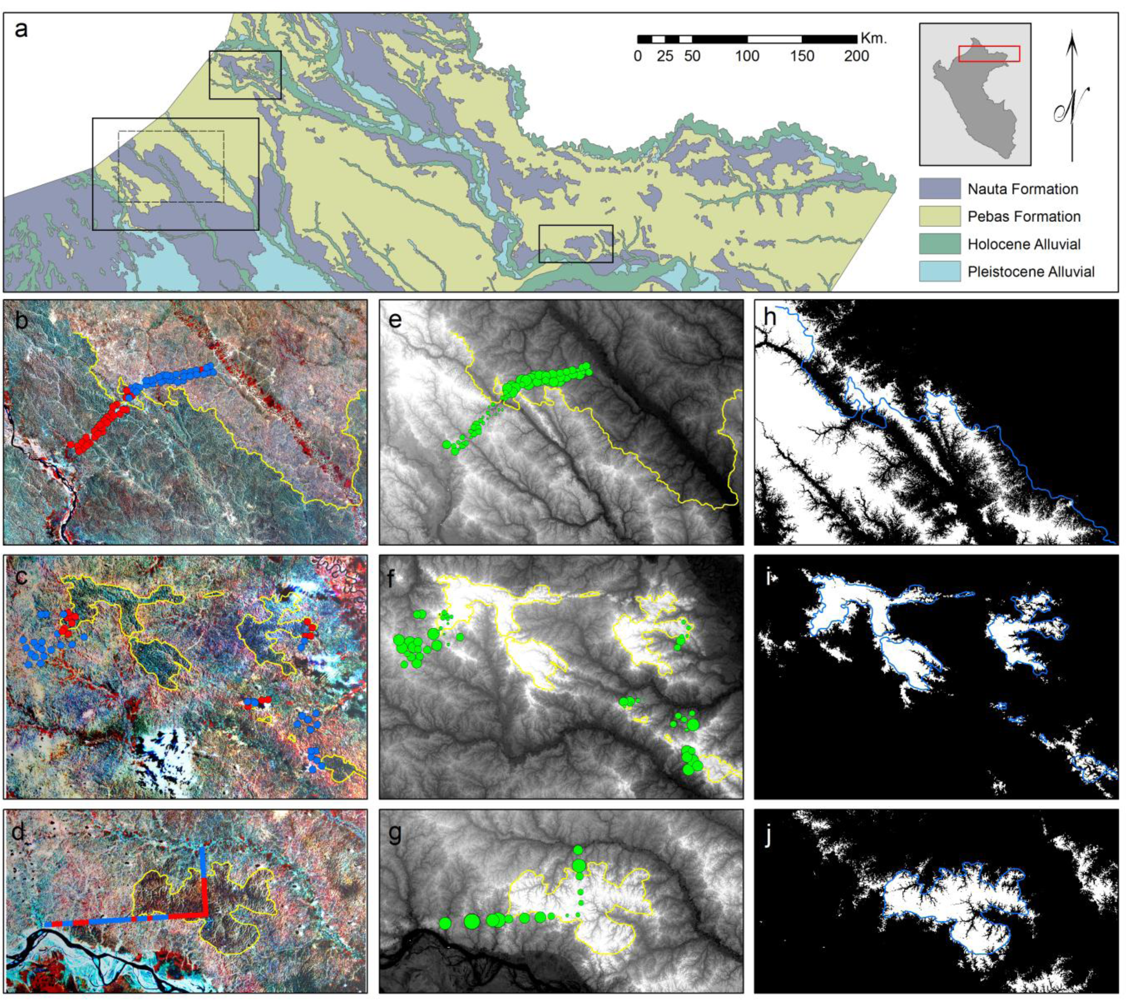
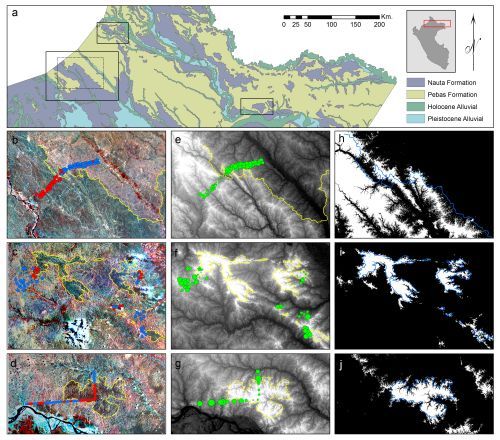
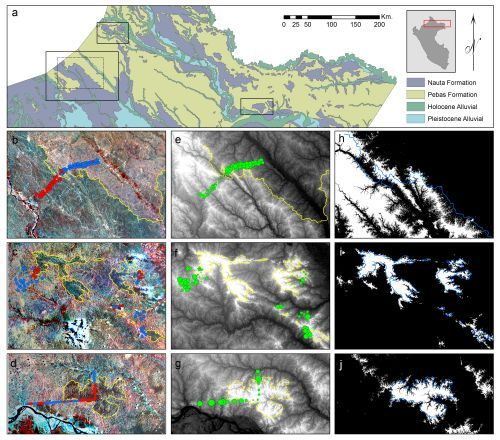
2.1. Study Areas
2.2. Satellite Imagery
2.3. Field Data
2.4. Data Analysis
3. Results
3.1. Image Interpretation
3.2. Floristic and Edaphic Patterns
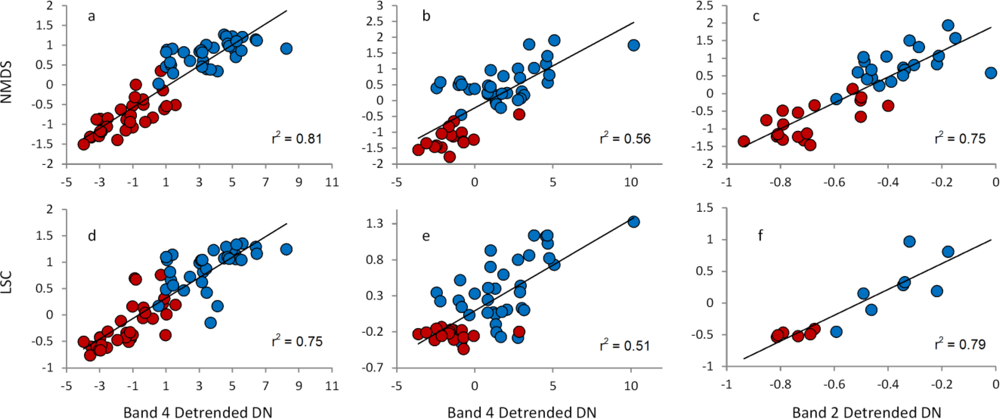
3.3. Relationships between Field Data and Landsat Data
3.4. Modeling of Floristic Patterns Based on SRTM Data
4. Discussion
5. Conclusions
Supporting Information 1. Landsat Image Detrending
remotesensing-04-02401-s001.pdfAcknowledgments
References
- Margules, C.R.; Pressey, R.L. Systematic conservation planning. Nature 2000, 405, 243–253. [Google Scholar]
- Duivenvoorden, J.F.; Lips, J.M. Ecología del paisaje del Medio Caquetá: Mapas; Estudios en la Amazonia Colombiana III B; Tropenbos: Bogotá, Colombia, 1993; Volume 3. [Google Scholar]
- Condit, R.; Pitman, N.; Leigh, E.G.; Chave, J.; Terborgh, J.; Foster, R.B.; Nunez, P.; Aguilar, S.; Valencia, R.; Villa, G.; Muller-Landau, H.C.; Losos, E.; Hubbell, S.P. Beta-diversity in tropical forest trees. Science 2002, 295, 666–669. [Google Scholar]
- Malhi, Y.; Baker, T.R.; Phillips, O.L.; Almeida, S.; Alvarez, E.; Arroyo, L.; Chave, J.; Czimczik, C.I.; Di Fiore, A.; Higuchi, N.; et al. The above-ground coarse wood productivity of 104 Neotropical forest plots. Glob. Change Biol 2004, 10, 563–591. [Google Scholar]
- Pitman, N.C.A.; Terborgh, J.W.; Silman, M.R.; Nunez, P.; Neill, D.A.; Ceron, C.E.; Palacios, W.A.; Aulestia, M. Dominance and distribution of tree species in upper Amazonian terra firme forests. Ecology 2001, 82, 2101–2117. [Google Scholar]
- ter Steege, H.; Pitman, N.C.A.; Phillips, O.L.; Chave, J.; Sabatier, D.; Duque, A.; Molino, J.F.; Prevost, M.F.; Spichiger, R.; Castellanos, H.; von Hildebrand, P.; Vasquez, R. Continental-scale patterns of canopy tree composition and function across Amazonia. Nature 2006, 443, 444–447. [Google Scholar]
- Duivenvoorden, J.F. Tree species composition and rain forest-environment relationships in the Middle Caqueta Area, Colombia, NW Amazonia. Vegetatio 1995, 120, 91–113. [Google Scholar]
- Higgins, M.A.; Ruokolainen, K.; Tuomisto, H.; Llerena, N.; Cardenas, G.; Phillips, O.L.; Vasquez, R.; Rasanen, M. Geological control of floristic composition in Amazonian forests. J. Biogeogr 2011, 38, 2136–2149. [Google Scholar]
- Phillips, O.L.; Vargas, P.N.; Monteagudo, A.L.; Cruz, A.P.; Zans, M.E.C.; Sanchez, W.G.; Yli-Halla, M.; Rose, S. Habitat association among Amazonian tree species: A landscape-scale approach. J. Ecol 2003, 91, 757–775. [Google Scholar]
- Pitman, N.C.A.; Mogollon, H.; Davila, N.; Rios, M.; Garcia-Villacorta, R.; Guevara, J.; Baker, T.R.; Monteagudo, A.; Phillips, O.L.; Vasquez-Martinez, R.; et al. Tree community change across 700 km of lowland Amazonian forest from the Andean foothills to Brazil. Biotropica 2008, 40, 525–535. [Google Scholar]
- Tuomisto, H.; Ruokolainen, K.; Yli-Halla, M. Dispersal, environment, and floristic variation of western Amazonian forests. Science 2003, 299, 241–244. [Google Scholar]
- Rodriguez, E.; Morris, C.S.; Belz, J.E. A global assessment of the SRTM performance. Photogramm. Eng. Remote Sensing 2006, 72, 249–260. [Google Scholar]
- Tucker, C.J.; Grant, D.M.; Dykstra, J.D. NASA’s global orthorectified Landsat data set. Photogramm. Eng. Remote Sensing 2004, 70, 313–322. [Google Scholar]
- Hill, R.A.; Foody, G.M. Separability of tropical rain-forest types in the Tambopata-Candamo Reserved Zone, Peru. Int. J. Remote Sens 1994, 15, 2687–2693. [Google Scholar]
- Salovaara, K.J.; Thessler, S.; Malik, R.N.; Tuomisto, H. Classification of Amazonian primary rain forest vegetation using Landsat ETM plus satellite imagery. Remote Sens. Environ 2005, 97, 39–51. [Google Scholar]
- Thessler, S.; Ruokolainen, K.; Tuomisto, H.; Tomppo, E. Mapping gradual landscape-scale floristic changes in Amazonian primary rain forests by combining ordination and remote sensing. Glob. Ecol. Biogeogr 2005, 14, 315–325. [Google Scholar]
- Tuomisto, H.; Poulsen, A.D.; Ruokolainen, K.; Moran, R.C.; Quintana, C.; Celi, J.; Canas, G. Linking floristic patterns with soil heterogeneity and satellite imagery in Ecuadorian Amazonia. Ecol. Appl 2003, 13, 352–371. [Google Scholar]
- Tuomisto, H.; Ruokolainen, K. Variación de los Bosques Naturales en las Áreas Piloto a lo Largo de Transectos y en Imágenes de Satélite. In Evaluación de Recursos Forestales no Maderables en la Amazonía Noroccidental; Duivenvoorden, J.F., Balslev, H., Cavelier, J., Grández, C., Tuomisto, H., Valencia, R., Eds.; IBED, Universiteit van Amsterdam: Amsterdam, The Netherlands, 2001; pp. 63–96. [Google Scholar]
- Tuomisto, H.; Ruokolainen, K.; Aguilar, M.; Sarmiento, A. Floristic patterns along a 43-km long transect in an Amazonian rain forest. J. Ecol 2003, 91, 743–756. [Google Scholar]
- Tuomisto, H.; Ruokolainen, K.; Kalliola, R.; Linna, A.; Danjoy, W.; Rodriguez, Z. Dissecting Amazonian biodiversity. Science 1995, 269, 63–66. [Google Scholar]
- Ruokolainen, K.; Tuomisto, H. Vegetación natural de la zona de Iquitos. In Geoecología y Desarrollo Amazonico: Estudio Integrado en la zona de Iquitos, Perú; Kalliola, R., Flores, S., Eds.; Annales Universitatis Turkuensis Ser A II: Turku, Finland, 1998; Volume 114, pp. 253–365. [Google Scholar]
- Rossetti, D.F.; Valeriano, M.M. Evolution of the lowest Amazon Basin modeled from the integration of geological and SRTM topographic data. Catena 2007, 70, 253–265. [Google Scholar]
- Asner, G.P.; Knapp, D.E.; Broadbent, E.N.; Oliveira, P.J.C.; Keller, M.; Silva, J.N. Selective logging in the Brazilian Amazon. Science 2005, 310, 480–482. [Google Scholar]
- Oliveira, P.J.C.; Asner, G.P.; Knapp, D.E.; Almeyda, A.; Galvan-Gildemeister, R.; Keene, S.; Raybin, R.F.; Smith, R.C. Land-use allocation protects the Peruvian Amazon. Science 2007, 317, 1233–1236. [Google Scholar]
- Toivonen, T.; Kalliola, R.; Ruokolainen, K.; Malik, R.N. Across-path DN gradient in Landsat TM imagery of Amazonian forests: A challenge for image interpretation and mosaicking. Remote Sens. Environ 2006, 100, 550–562. [Google Scholar]
- INGEMMET, Mapa Geológico del Perú; Instituto Geologico Minero Y Metalurgico: Lima, 2000.
- Hoorn, C.; Wesselingh, F.P.; ter Steege, H.; Bermudez, M.A.; Mora, A.; Sevink, J.; Sanmartin, I.; Sanchez-Meseguer, A.; Anderson, C.L.; Figueiredo, J.P.; et al. Amazonia through time: Andean uplift, climate change, landscape evolution, and biodiversity. Science 2010, 330, 927–931. [Google Scholar]
- Räsänen, M.E.; Linna, A.M.; Santos, J.C.R.; Negri, F.R. Late miocene tidal deposits in the Amazonian Foreland Basin. Science 1995, 269, 386–390. [Google Scholar]
- Rebata, L.A.; Gingras, M.K.; Räsänen, M.E.; Barberi, M. Tidal-channel deposits on a delta plain from the Upper Miocene Nauta Formation, Marañon Foreland Sub-basin, Peru. Sedimentology 2006, 53, 971–1013. [Google Scholar]
- Figueiredo, J.; Hoorn, C.; van der Ven, P.; Soares, E. Late Miocene onset of the Amazon River and the Amazon deep-sea fan: Evidence from the Foz do Amazonas Basin. Geology 2009, 37, 619–622. [Google Scholar]
- Räsänen, M.E.; Salo, J.S.; Jungner, H.; Pittman, L.R. Evolution of the western Amazonian lowland relief: Impact of Andean foreland dynamics. Terra Nova 1990, 2, 320–332. [Google Scholar]
- Roddaz, M.; Baby, P.; Brusset, S.; Hermoza, W.; Darrozes, J.M. Forebulge dynamics and environmental control in Western Amazonia: The case study of the Arch of Iquitos (Peru). Tectonophysics 2005, 399, 87–108. [Google Scholar]
- Tuomisto, H.; Linna, A.; Kalliola, R. Use of digitally processed satellite images in studies of tropical rain-forest vegetation. Int. J. Remote Sens 1994, 15, 1595–1610. [Google Scholar]
- Ruokolainen, K.; Linna, A.; Tuomisto, H. Use of Melastomataceae and pteridophytes for revealing phytogeographical patterns in Amazonian rain forests. J. Trop. Ecol 1997, 13, 243–256. [Google Scholar]
- Ruokolainen, K.; Tuomisto, H.; Macia, M.J.; Higgins, M.A.; Yli-Halla, M. Are floristic and edaphic patterns in Amazonian rain forests congruent for trees, pteridophytes and Melastomataceae? J. Trop. Ecol 2007, 23, 13–25. [Google Scholar]
- Jones, M.M.; Tuomisto, H.; Olivas, P.C. Differences in the degree of environmental control on large and small tropical plants: Just a sampling effect? J. Ecol 2008, 96, 367–377. [Google Scholar]
- Thiers, B. Index Herbariorum: A Global Directory of Public Herbaria and Associated Staff. New York Botanical Garden’s Virtual Herbarium. Available online: http://sweetgum.nybg.org/ih/ (accessed on 20 June 2012).
- Legendre, P.; Legendre, L. Numerical Ecology; Elsevier: Amsterdam, The Netherlands, 1998; Volume 20, pp. 1–853. [Google Scholar]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees; Wadsworth and Brooks/Cole: Monterey, CA, USA, 1984. [Google Scholar]
- Fine, P.V.A.; Mesones, I.; Coley, P.D. Herbivores promote habitat specialization by trees in Amazonian forests. Science 2004, 305, 663–665. [Google Scholar]
- Asner, G.P.; Martin, R.E. Canopy phylogenetic, chemical and spectral assembly in a lowland Amazonian forest. New Phytol 2011, 189, 999–1012. [Google Scholar]
- Lieberman, D.; Lieberman, M.; Peralta, R.; Hartshorn, G.S. Tropical forest structure and composition on a large-scale altitudinal gradient in Costa Rica. J. Ecol 1996, 84, 137–152. [Google Scholar]
- Vormisto, J.; Tuomisto, H.; Oksanen, J. Palm distribution patterns in Amazonian rainforests: What is the role of topographic variation? J. Veg. Sci 2004, 15, 485–494. [Google Scholar]
- Higgins, M.; Ruokolainen, K. Rapid tropical forest inventory: A comparison of techniques based on inventory data from western Amazonia. Conserv. Biol 2004, 18, 799–811. [Google Scholar]
- UNEP-WCMC. World Database on Protected Areas (WDPA) Annual Release 2009. Available online: http://www.wdpa.org/AnnualRelease.aspx (accessed on 20 June 2009).
- Anderson, L.O. Biome-scale forest properties in Amazonia based on field and satellite observations. Remote Sens 2012, 4, 1245–1271. [Google Scholar]
- Zuquim, G.; Costa, F.R.C.; Prado, J.; Tuomisto, H. Guide to the Ferns and Lycophytes of REBIO Uatumã, Central Amazonia; Instituto Nacional de Pesquisas da Amazônia: Manaus, Brazil, 2008. [Google Scholar]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pastaza-Tigre | Curaray | Sucusari | ||||
|---|---|---|---|---|---|---|
| Band | Raw | Detrended | Raw | Detrended | Raw | Detrended |
| 1 | 0.32 | (0.11) | (0.01) | (0.01) | 0.27 | 0.24 |
| 2 | (0.07) | 0.40 | (0.04) | (0.06) | 0.56 | 0.75 |
| 3 | 0.40 | 0.23 | (0.10) | (0.11) | (0.17) | (0.11) |
| 4 | 0.68 | 0.81 | 0.55 | 0.56 | 0.34 | 0.24 |
| 5 | 0.40 | 0.55 | (0.03) | 0.20 | 0.55 | 0.39 |
| 7 | 0.47 | 0.58 | (0.00) | (0.12) | 0.64 | 0.50 |
| NDVI | 0.75 | 0.83 | 0.53 | 0.51 | (0.20) | (0.11) |
| Pastaza-Tigre | Curaray | Sucusari | ||||
|---|---|---|---|---|---|---|
| Band | Raw | Detrended | Raw | Detrended | Raw | Detrended |
| 1 | 0.19 | (0.07) | (0.00) | (0.00) | (0.56) | (0.37) |
| 2 | (0.09) | 0.31 | (0.06) | (0.06) | 0.74 | 0.79 |
| 3 | 0.38 | 0.26 | (0.04) | (0.06) | (0.38) | (0.26) |
| 4 | 0.68 | 0.75 | 0.54 | 0.51 | (0.22) | (0.08) |
| 5 | 0.39 | 0.49 | (0.03) | (0.12) | (0.23) | (0.08) |
| 7 | 0.42 | 0.49 | (0.01) | (0.08) | (0.35) | (0.19) |
| NDVI | 0.75 | 0.78 | 0.43 | 0.42 | (0.08) | (0.01) |
| Independent Variable | Imagery | All Bands | Bands 4, 5, 7 | Bands 4, 5, 7, Enhanced |
|---|---|---|---|---|
| Floristic composition | Raw | 0.52 | 0.61 | 0.53 |
| Detrended | 0.75 | 0.75 | 0.65 | |
| Soils | Raw | 0.53 | 0.60 | 0.50 |
| Detrended | 0.69 | 0.68 | 0.58 | |
Share and Cite
Higgins, M.A.; Asner, G.P.; Perez, E.; Elespuru, N.; Tuomisto, H.; Ruokolainen, K.; Alonso, A. Use of Landsat and SRTM Data to Detect Broad-Scale Biodiversity Patterns in Northwestern Amazonia. Remote Sens. 2012, 4, 2401-2418. https://doi.org/10.3390/rs4082401
Higgins MA, Asner GP, Perez E, Elespuru N, Tuomisto H, Ruokolainen K, Alonso A. Use of Landsat and SRTM Data to Detect Broad-Scale Biodiversity Patterns in Northwestern Amazonia. Remote Sensing. 2012; 4(8):2401-2418. https://doi.org/10.3390/rs4082401
Chicago/Turabian StyleHiggins, Mark A., Gregory P. Asner, Eneas Perez, Nydia Elespuru, Hanna Tuomisto, Kalle Ruokolainen, and Alfonso Alonso. 2012. "Use of Landsat and SRTM Data to Detect Broad-Scale Biodiversity Patterns in Northwestern Amazonia" Remote Sensing 4, no. 8: 2401-2418. https://doi.org/10.3390/rs4082401
APA StyleHiggins, M. A., Asner, G. P., Perez, E., Elespuru, N., Tuomisto, H., Ruokolainen, K., & Alonso, A. (2012). Use of Landsat and SRTM Data to Detect Broad-Scale Biodiversity Patterns in Northwestern Amazonia. Remote Sensing, 4(8), 2401-2418. https://doi.org/10.3390/rs4082401