
Global Distribution and Local Variation of Pre-Rain Green-Up in Tropical Dryland


Abstract
1. Introduction
- To describe the global distribution of the pre-rain green-up phenomenon by vegetation type and topography conditions using coarse-resolution imagery.
- To observe and compare the spring phenology of pre-rain green-up and post-rain green-up vegetation in mountainous regions at the small watershed scale and to analyze the relationship between spring phenology and elevation using high-resolution satellite data.
- To conduct regional-scale mapping and quantitative analysis of the aforementioned relationships.
2. Materials and Methods
2.1. Data
2.1.1. Phenology and Rainfall Data
2.1.2. Land Cover and Topography Data
2.2. Study Area
2.3. Methods
2.3.1. Extraction of the Start of the Growing Season
2.3.2. Extraction of the Start of the Rainy Season
2.3.3. Covariation Between SOS and Elevation
3. Results
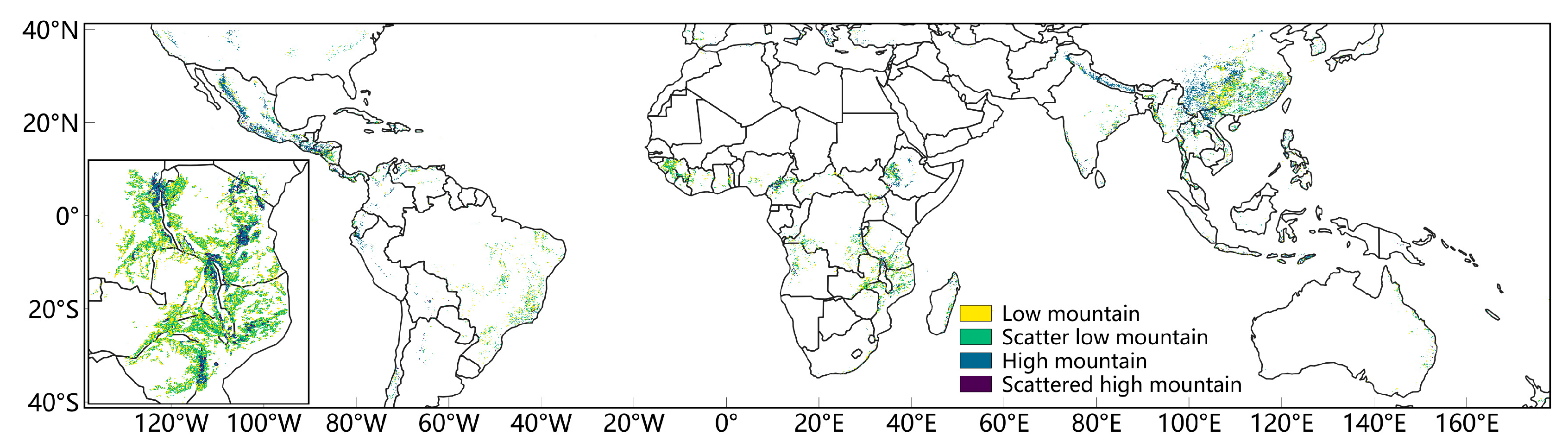
3.1. Spatial Distribution of the Pre-Rain Green-Up Phenomenon in the Mid to Low Latitudes
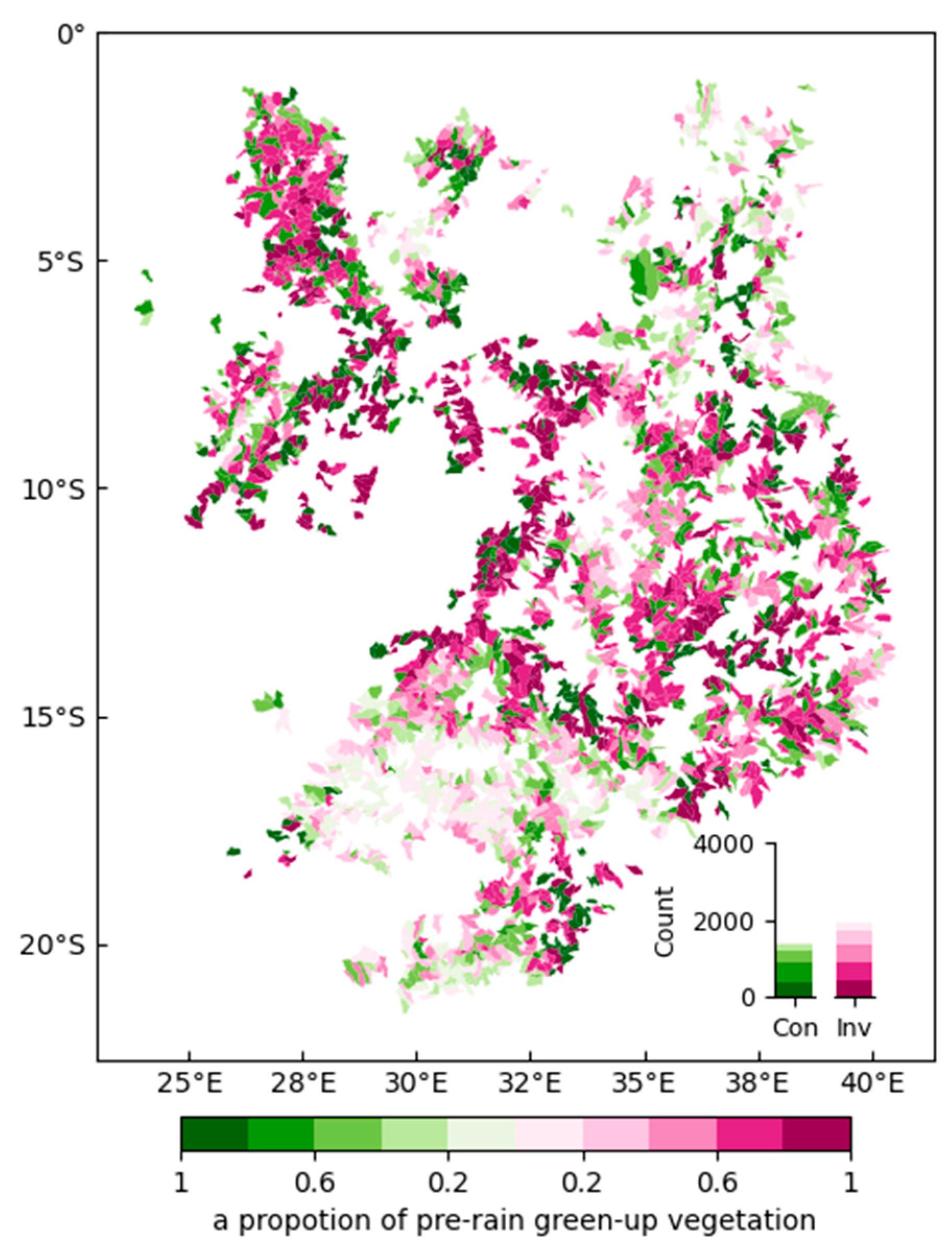
3.2. High-Resolution Mapping of the Pre-Rain Green-Up Phenomenon in African Mountainous Areas
3.3. The SOS–Elevation Relationship of Pre-Rain Green-Up Areas
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A




References
- Ryan, C.M.; Williams, M.; Grace, J.; Woollen, E.; Lehmann, C.E.R. Pre-rain Green-up Is Ubiquitous across Southern Tropical Africa: Implications for Temporal Niche Separation and Model Representation. New Phytol. 2017, 213, 625–633. [Google Scholar] [CrossRef]
- Verger, A.; Filella, I.; Baret, F.; Peñuelas, J. Vegetation Baseline Phenology from Kilometric Global LAI Satellite Products. Remote Sens. Environ. 2016, 178, 1–14. [Google Scholar] [CrossRef]
- Di Lucchio, L.M.; Fensholt, R.; Markussen, B.; Ræbild, A. Leaf Phenology of Thirteen African Origins of Baobab (Adansonia digitata (L.)) as Influenced by Daylength and Water Availability. Ecol. Evol. 2018, 8, 11261–11272. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Myers, B.A.; Muller, W.J.; Duff, G.A.; Eamus, D. Leaf phenology of woody species in a north Australian tropical savanna. Ecology 1997, 78, 2542–2558. [Google Scholar] [CrossRef]
- Elliott, S.; Baker, P.J.; Borchert, R. Leaf Flushing during the Dry Season: The Paradox of Asian Monsoon Forests. Glob. Ecol. Biogeogr. 2006, 15, 248–257. [Google Scholar] [CrossRef]
- Arantes, A.E.; Ferreira, L.G.; Coe, M.T. The Seasonal Carbon and Water Balances of the Cerrado Environment of Brazil: Past, Present, and Future Influences of Land Cover and Land Use. ISPRS J. Photogramm. Remote Sens. 2016, 117, 66–78. [Google Scholar] [CrossRef]
- Adole, T.; Dash, J.; Atkinson, P.M. Large-Scale Prerain Vegetation Green-up across Africa. Glob. Chang. Biol. 2018, 24, 4054–4068. [Google Scholar] [CrossRef]
- Chen, X.; Chen, W.; Xu, M. Remote-Sensing Monitoring of Postfire Vegetation Dynamics in the Greater Hinggan Mountain Range Based on Long Time-Series Data: Analysis of the Effects of Six Topographic and Climatic Factors. Remote Sens. 2022, 14, 2958. [Google Scholar] [CrossRef]
- Afuye, G.A.; Kalumba, A.M.; Orimoloye, I.R. Characterisation of Vegetation Response to Climate Change: A Review. Sustainability 2021, 13, 7265. [Google Scholar] [CrossRef]
- Wigley, B.J.; Coetsee, C.; February, E.C.; Dobelmann, S.; Higgins, S.I. Will Trees or Grasses Profit from Changing Rainfall Regimes in Savannas? New Phytol. 2024, 241, 2379–2394. [Google Scholar] [CrossRef]
- Dohn, J.; Dembélé, F.; Karembé, M.; Moustakas, A.; Amévor, K.A.; Hanan, N.P. Tree Effects on Grass Growth in Savannas: Competition, Facilitation and the Stress-Gradient Hypothesis. J. Ecol. 2013, 101, 202–209. [Google Scholar] [CrossRef]
- House, J.I.; Archer, S.; Breshears, D.D.; Scholes, R.J. NCEAS Tree–Grass Interactions Participants Conundrums in Mixed Woody–Herbaceous Plant Systems. J. Biogeogr. 2003, 30, 1763–1777. [Google Scholar] [CrossRef]
- Dobson, A.; Hopcraft, G.; Mduma, S.; Ogutu, J.O.; Fryxell, J.; Anderson, T.M.; Archibald, S.; Lehmann, C.; Poole, J.; Caro, T.; et al. Savannas Are Vital but Overlooked Carbon Sinks. Science 2022, 375, 392. [Google Scholar] [CrossRef]
- Zhou, Y.; Bomfim, B.; Bond, W.J.; Boutton, T.W.; Case, M.F.; Coetsee, C.; Davies, A.B.; February, E.C.; Gray, E.F.; Silva, L.C.R.; et al. Soil Carbon in Tropical Savannas Mostly Derived from Grasses. Nat. Geosci. 2023, 16, 710–716. [Google Scholar] [CrossRef]
- Guan, K.; Wood, E.F.; Medvigy, D.; Kimball, J.; Pan, M.; Caylor, K.K.; Sheffield, J.; Xu, X.; Jones, M.O. Terrestrial Hydrological Controls on Land Surface Phenology of African Savannas and Woodlands. J. Geophys. Res. Biogeosciences 2014, 119, 1652–1669. [Google Scholar] [CrossRef]
- Seghieri, J.; Do, F.C.; Devineau, J.-L.; Fournier, A.; Seghieri, J.; Do, F.C.; Devineau, J.-L.; Fournier, A. Phenology of Woody Species Along the Climatic Gradient in West Tropical Africa. In Phenology and Climate Change; IntechOpen: London, UK, 2012; ISBN 978-953-51-0336-3. [Google Scholar]
- Whitecross, M.A.; Witkowski, E.T.F.; Archibald, S. Savanna Tree-Grass Interactions: A Phenological Investigation of Green-up in Relation to Water Availability over Three Seasons. South Afr. J. Bot. 2017, 108, 29–40. [Google Scholar] [CrossRef]
- Venter, S.M.; Witkowski, E.T.F. Phenology, Flowering and Fruit-Set Patterns of Baobabs, Adansonia Digitata, in Southern Africa. For. Ecol. Manag. 2019, 453, 117593. [Google Scholar] [CrossRef]
- Godlee, J.L.; Ryan, C.M.; Siampale, A.; Dexter, K.G. Tree Species Diversity Drives the Land Surface Phenology of Seasonally Dry Tropical Woodlands. J. Ecol. 2024, 112, 1978–1991. [Google Scholar] [CrossRef]
- Adole, T.; Dash, J.; Atkinson, P.M. Characterising the Land Surface Phenology of Africa Using 500 m MODIS EVI. Appl. Geogr. 2018, 90, 187–199. [Google Scholar] [CrossRef]
- Cizek, A.; Aplin, P.; Powell, I. Measuring the Timing of Woody Green-Up in African Savannas—Which Modis Data to Use? In Proceedings of the 2021 IEEE International Geoscience and Remote Sensing Symposium IGARSS, Brussels, Belgium, 11–16 July 2021; pp. 1398–1401. [Google Scholar]
- Guan, K.; Good, S.P.; Caylor, K.K.; Medvigy, D.; Pan, M.; Wood, E.F.; Sato, H.; Biasutti, M.; Chen, M.; Ahlström, A.; et al. Simulated Sensitivity of African Terrestrial Ecosystem Photosynthesis to Rainfall Frequency, Intensity, and Rainy Season Length. Environ. Res. Lett. 2018, 13, 025013. [Google Scholar] [CrossRef]
- Adole, T.; Dash, J.; Rodriguez-Galiano, V.; Atkinson, P.M. Photoperiod Controls Vegetation Phenology across Africa. Commun. Biol. 2019, 2, 391. [Google Scholar] [CrossRef] [PubMed]
- Stocker, B.D.; Tumber-Dávila, S.J.; Konings, A.G.; Anderson, M.C.; Hain, C.; Jackson, R.B. Global Patterns of Water Storage in the Rooting Zones of Vegetation. Nat. Geosci. 2023, 16, 250–256. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Si, B.; Ma, X.; Wu, P. Deep Soil Water Extraction by Apple Sequesters Organic Carbon via Root Biomass Rather than Altering Soil Organic Carbon Content. Sci. Total Environ. 2019, 670, 662–671. [Google Scholar] [CrossRef] [PubMed]
- Sulla-Menashe, D.; Friedl, M.A. User Guide to Collection 6 MODIS Land Cover (MCD12Q1 and MCD12C1) Product; USGS: Reston, VA, USA, 2018. [Google Scholar]
- Funk, C.; Peterson, P.; Landsfeld, M.; Pedreros, D.; Verdin, J.; Shukla, S.; Husak, G.; Rowland, J.; Harrison, L.; Hoell, A.; et al. The Climate Hazards Infrared Precipitation with Stations—A New Environmental Record for Monitoring Extremes. Sci. Data 2015, 2, 150066. [Google Scholar] [CrossRef]
- Sayre, R.; Frye, C.; Karagulle, D.; Krauer, J.; Breyer, S.; Aniello, P.; Wright, D.J.; Payne, D.; Adler, C.; Warner, H.; et al. A New High-Resolution Map of World Mountains and an Online Tool for Visualizing and Comparing Characterizations of Global Mountain Distributions. Mt. Res. Dev. 2018, 38, 240–249. [Google Scholar] [CrossRef]
- Lehner, B.; Grill, G. Global River Hydrography and Network Routing: Baseline Data and New Approaches to Study the World’s Large River Systems. Hydrol. Process. 2013, 27, 2171–2186. [Google Scholar] [CrossRef]
- Crippen, R.; Buckley, S.; Agram, P.; Belz, E.; Gurrola, E.; Hensley, S.; Kobrick, M.; Lavalle, M.; Martin, J.; Neumann, M.; et al. Nasadem global elevation model: Methods and progress. Int. Arch. Photogramm. Remote Sens. Spat. Inf. Sci. 2016, 41, 125–128. [Google Scholar] [CrossRef]
- Buckley, S.M.; Agram, P.S.; Belz, J.E.; Crippen, R.E.; Gurrola, E.M.; Hensley, S.; Kobrick, M.; Lavalle, M.; Martin, J.M.; Neumann, M.; et al. NASADEM: User Guide. 2020. Available online: https://lpdaac.usgs.gov/documents/592/NASADEM_User_Guide_V1.pdf (accessed on 1 September 2022).
- ESA. Level-2A Algorithm Theoretical Basis Document; Remote Sensing Systems: Santa Rosa, CA, USA, 2021. [Google Scholar]
- Gray, J.; Sulla-Menashe, D.; Friedl, M.A. User Guide to Collection 6 MODIS Land Cover Dynamics (MCD12Q2) Product; NASA EOSDIS Land Processes DAAC: Missoula, MT, USA, 2019. [Google Scholar]
- Zanaga, D.; Van De Kerchove, R.; De Keersmaecker, W.; Souverijns, N.; Brockmann, C.; Quast, R.; Wevers, J.; Grosu, A.; Paccini, A.; Vergnaud, S.; et al. ESA WorldCover 10 m 2020 V100. 2021. Available online: https://pure.iiasa.ac.at/id/eprint/17832/ (accessed on 1 September 2022).
- Campbell, J.T. Middle Passages: African American Journeys to Africa, 1787–2005; Penguin Books: New York, NY, USA, 2007; ISBN 978-0-14-311198-6. [Google Scholar]
- Zhou, J.; Menenti, M.; Jia, L.; Gao, B.; Zhao, F.; Cui, Y.; Xiong, X.; Liu, X.; Li, D. A Scalable Software Package for Time Series Reconstruction of Remote Sensing Datasets on the Google Earth Engine Platform. Int. J. Digit. Earth 2023, 16, 988–1007. [Google Scholar] [CrossRef]
- Jonsson, P.; Eklundh, L. Seasonality Extraction by Function Fitting to Time-Series of Satellite Sensor Data. IEEE Trans. Geosci. Remote Sens. 2002, 40, 1824–1832. [Google Scholar] [CrossRef]
- Wang, L.; Tian, F.; Wang, Y.; Wu, Z.; Schurgers, G.; Fensholt, R. Acceleration of Global Vegetation Greenup from Combined Effects of Climate Change and Human Land Management. Glob. Chang. Biol. 2018, 24, 5484–5499. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. TIMESAT—A Program for Analyzing Time-Series of Satellite Sensor Data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Dunning, C.M.; Black, E.C.L.; Allan, R.P. The Onset and Cessation of Seasonal Rainfall over Africa. J. Geophys. Res. Atmos. 2016, 121, 11405–11424. [Google Scholar] [CrossRef]
- Bolton, D.K.; Gray, J.M.; Melaas, E.K.; Moon, M.; Eklundh, L.; Friedl, M.A. Continental-Scale Land Surface Phenology from Harmonized Landsat 8 and Sentinel-2 Imagery. Remote Sens. Environ. 2020, 240, 111685. [Google Scholar] [CrossRef]
- Liebmann, B.; Bladé, I.; Kiladis, G.N.; Carvalho, L.M.V.; Senay, G.B.; Allured, D.; Leroux, S.; Funk, C. Seasonality of African Precipitation from 1996 to 2009. J. Clim. 2012, 25, 4304–4322. [Google Scholar] [CrossRef]
- Tian, J.; Luo, X.; Xu, H.; Green, J.K.; Tang, H.; Wu, J.; Piao, S. Slower Changes in Vegetation Phenology than Precipitation Seasonality in the Dry Tropics. Glob. Chang. Biol. 2024, 30, e17134. [Google Scholar] [CrossRef]
- Dunning, C.M.; Black, E.; Allan, R.P. Later Wet Seasons with More Intense Rainfall over Africa under Future Climate Change. J. Clim. 2018, 31, 9719–9738. [Google Scholar] [CrossRef]
- Ding, C.; Li, Y.; Xie, Q.; Li, H.; Zhang, B. Impacts of Terrain on Land Surface Phenology Derived from Harmonized Landsat 8 and Sentinel-2 in the Tianshan Mountains, China. GIScience Remote Sens. 2023, 60, 2242621. [Google Scholar] [CrossRef]
- Higgins, S.I.; Delgado-Cartay, M.D.; February, E.C.; Combrink, H.J. Is There a Temporal Niche Separation in the Leaf Phenology of Savanna Trees and Grasses? J. Biogeogr. 2011, 38, 2165–2175. [Google Scholar] [CrossRef]
- Yan, D.; Zhang, X.; Yu, Y.; Guo, W. Characterizing Land Cover Impacts on the Responses of Land Surface Phenology to the Rainy Season in the Congo Basin. Remote Sens. 2017, 9, 461. [Google Scholar] [CrossRef]
- Gao, M.; Piao, S.; Chen, A.; Yang, H.; Liu, Q.; Fu, Y.H.; Janssens, I.A. Divergent Changes in the Elevational Gradient of Vegetation Activities over the Last 30 Years. Nat. Commun. 2019, 10, 2970. [Google Scholar] [CrossRef]
- An, S.; Zhang, X.; Chen, X.; Yan, D.; Henebry, G.M. An Exploration of Terrain Effects on Land Surface Phenology across the Qinghai–Tibet Plateau Using Landsat ETM+ and OLI Data. Remote Sens. 2018, 10, 1069. [Google Scholar] [CrossRef]
- Hua, X.; Ohlemüller, R.; Sirguey, P. Differential Effects of Topography on the Timing of the Growing Season in Mountainous Grassland Ecosystems. Environ. Adv. 2022, 8, 100234. [Google Scholar] [CrossRef]
- Ding, C.; Huang, W.; Liu, M.; Zhao, S. Change in the Elevational Pattern of Vegetation Greenup Date across the Tianshan Mountains in Central Asia during 2001–2020. Ecol. Indic. 2022, 136, 108684. [Google Scholar] [CrossRef]
- Fisher, J.; Mustard, J.; Vadeboncoeur, M. Green Leaf Phenology at Landsat Resolution: Scaling from the Field to the Satellite. Remote Sens. Environ. 2006, 100, 265–279. [Google Scholar] [CrossRef]
- Cizek, A.F. Complex Spatial Patterns of Leaf Phenology within Semi-arid African Savanna Landscapes Driven by Rainfall and Topo-edaphic Variation Revealed Using Satellite Remote Sensing; Edge Hill University: Ormskirk, UK, 2024. [Google Scholar]
- Angoboy Ilondea, B.; Beeckman, H.; Van Acker, J.; Van Den Bulcke, J.; Fayolle, A.; Couralet, C.; Hubau, W.; Kafuti, C.; Rousseau, M.; Kaka di-Makwala, A.; et al. Variation in Onset of Leaf Unfolding and Wood Formation in a Central African Tropical Tree Species. Front. For. Glob. Chang. 2021, 4, 673575. [Google Scholar] [CrossRef]
- Scholes, R.J.; Walker, B.H. An African Savanna: Synthesis of the Nylsvley Study, 1st ed.; Cambridge University Press: Cambridge, UK, 1993; ISBN 978-0-521-41971-0. [Google Scholar]
- Archibald, S.; Scholes, R.J. Leaf Green-up in a Semi-Arid African Savanna -Separating Tree and Grass Responses to Environmental Cues. J. Veg. Sci. 2007, 18, 583–594. [Google Scholar] [CrossRef]
- Do, F.C.; Goudiaby, V.A.; Gimenez, O.; Diagne, A.L.; Diouf, M.; Rocheteau, A.; Akpo, L.E. Environmental Influence on Canopy Phenology in the Dry Tropics. For. Ecol. Manag. 2005, 215, 319–328. [Google Scholar] [CrossRef]
- Bond, W.J. What Limits Trees in C4 Grasslands and Savannas? Annu. Rev. Ecol. Evol. Syst. 2008, 39, 641–659. [Google Scholar] [CrossRef]
- Zhou, Y.; Wigley, B.J.; Case, M.F.; Coetsee, C.; Staver, A.C. Rooting Depth as a Key Woody Functional Trait in Savannas. New Phytol. 2020, 227, 1350–1361. [Google Scholar] [CrossRef]
- Cheng, Y.; Vrieling, A.; Fava, F.; Meroni, M.; Marshall, M.; Gachoki, S. Phenology of Short Vegetation Cycles in a Kenyan Rangeland from PlanetScope and Sentinel-2. Remote Sens. Environ. 2020, 248, 112004. [Google Scholar] [CrossRef]
- Tian, F.; Cai, Z.; Jin, H.; Hufkens, K.; Scheifinger, H.; Tagesson, T.; Smets, B.; Van Hoolst, R.; Bonte, K.; Ivits, E.; et al. Calibrating Vegetation Phenology from Sentinel-2 Using Eddy Covariance, PhenoCam, and PEP725 Networks across Europe. Remote Sens. Environ. 2021, 260, 112456. [Google Scholar] [CrossRef]
- Abdi, A.M.; Brandt, M.; Abel, C.; Fensholt, R. Satellite Remote Sensing of Savannas: Current Status and Emerging Opportunities. J. Remote Sens. 2022, 2022. [Google Scholar] [CrossRef]
- Haverd, V.; Smith, B.; Raupach, M.; Briggs, P.; Nieradzik, L.; Beringer, J.; Hutley, L.; Trudinger, C.M.; Cleverly, J. Coupling Carbon Allocation with Leaf and Root Phenology Predicts Tree–Grass Partitioning along a Savanna Rainfall Gradient. Biogeosciences 2016, 13, 761–779. [Google Scholar] [CrossRef]
- Whitecross, M.A.; Witkowski, E.T.F.; Archibald, S. Assessing the Frequency and Drivers of Early-greening in Broad-leaved Woodlands along a Latitudinal Gradient in Southern Africa. Austral Ecol. 2017, 42, 341–353. [Google Scholar] [CrossRef]
- Sang, Y.; Tian, F.; Jin, H.; Cai, Z.; Feng, L.; Dou, Y.; Eklundh, L. Assessing Topographic Effects on Forest Responses to Drought with Multiple Seasonal Metrics from Sentinel-2. Int. J. Appl. Earth Obs. Geoinf. 2024, 128, 103789. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Datasets | Spatial Resolution | Time | References |
|---|---|---|---|
| Harmonized Sentinel-2 Level-2A Collection | 10 m | 2019–2020 | ESA, 2020 [32] |
| MCD12Q1 | 500 m | 2019 | Sulla-Menashe & Friedl, 2018 [26] |
| MCD12Q2 | 500 m | 2019–2020 | Gray et al., 2019 [33] |
| CHIRPS | 0.05° | 2000–2020 | Funk et al., 2015 [27] |
| ESA WorldCover product | 10 m | 2020 | Zanaga et al., 2021 [34] |
| Sayre’s world mountain map | 250 m | 2018 | Sayre et al., 2018 [28] |
| HydroBASINS | - | 2013 | Lehner & Grill, 2013 [29] |
| NASADEM | 30 m | 2019 | Crippen et al., 2016 [30] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, S.; Sang, Y.; Cai, Z.; Tian, F. Global Distribution and Local Variation of Pre-Rain Green-Up in Tropical Dryland. Remote Sens. 2025, 17, 1377. https://doi.org/10.3390/rs17081377
Huang S, Sang Y, Cai Z, Tian F. Global Distribution and Local Variation of Pre-Rain Green-Up in Tropical Dryland. Remote Sensing. 2025; 17(8):1377. https://doi.org/10.3390/rs17081377
Chicago/Turabian StyleHuang, Shuyi, Yirong Sang, Zhanzhang Cai, and Feng Tian. 2025. "Global Distribution and Local Variation of Pre-Rain Green-Up in Tropical Dryland" Remote Sensing 17, no. 8: 1377. https://doi.org/10.3390/rs17081377
APA StyleHuang, S., Sang, Y., Cai, Z., & Tian, F. (2025). Global Distribution and Local Variation of Pre-Rain Green-Up in Tropical Dryland. Remote Sensing, 17(8), 1377. https://doi.org/10.3390/rs17081377