
Solar-Induced Fluorescence as Indicator of Downy Oak and the Influence of Some Environmental Variables at the End of the Growing Season

 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Site
2.1.1. Location
2.1.2. History and Infrastructure
2.1.3. Characteristics of the Oak Forest
2.2. Data Monitoring
Measurements of Environmental Conditions: ICOS Tower and O3HP Site
2.3. Data Filtering
2.4. Data Analysis
3. Results
3.1. Seasonal Dynamics of Photosynthesis and Phenology
3.1.1. Evolution of the Solar-Induced Fluorescence (SIF)
3.1.2. Evolution of Vegetation Indices
3.1.3. Pearson Correlations: SIFs—Environmental Conditions and SIFs—Vegetation Indices
3.2. Diurnal Dynamics of SIFs, Vegetation Indices and Environmental Parameters
3.2.1. Evolution of Solar-Induced Fluorescence
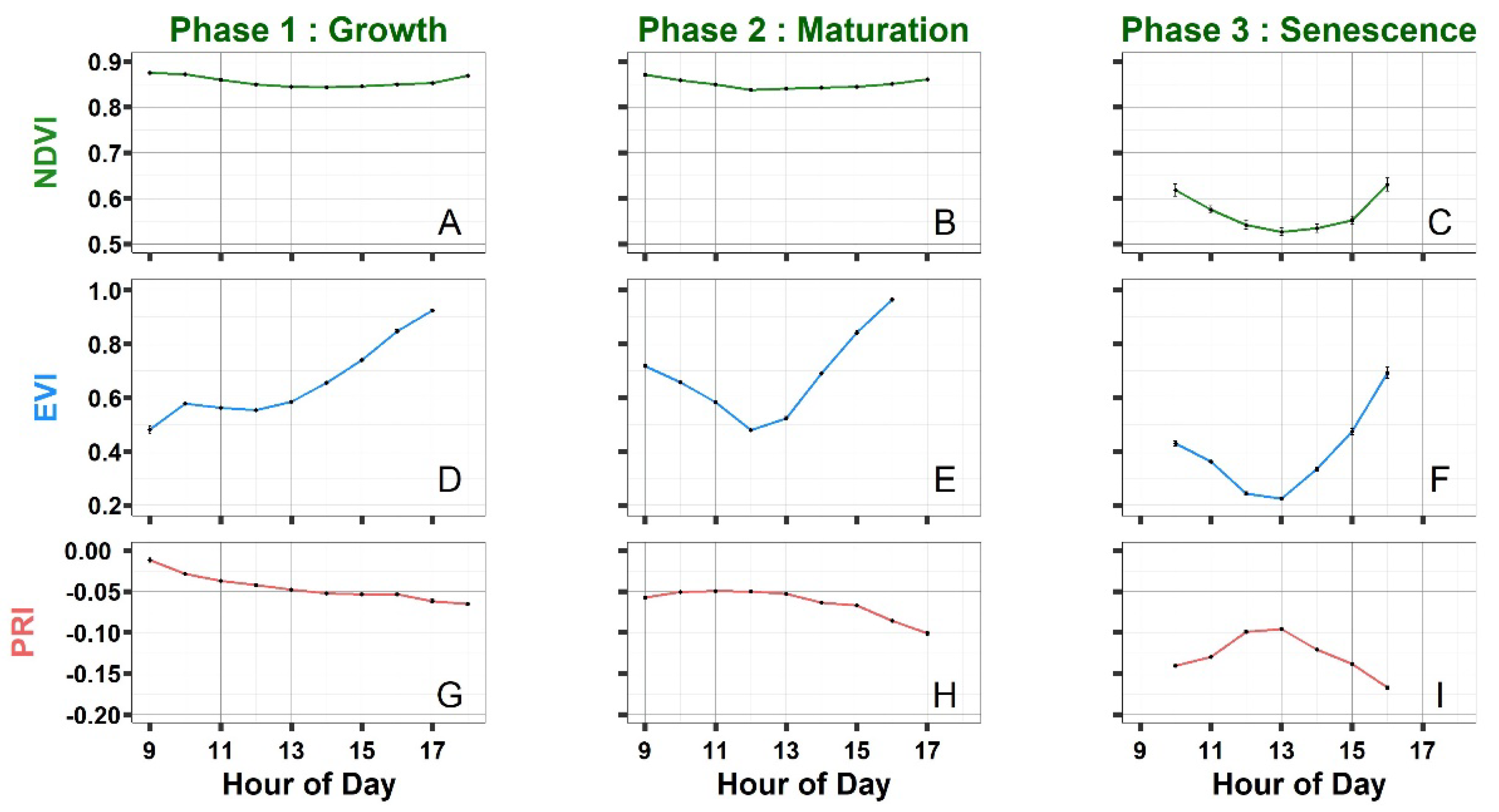
3.2.2. Evolution of Vegetation Indices
3.2.3. Pearson Correlations: SIFs—Environmental Conditions and SIFs—Vegetation Indices
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thomson, A.M.; César Izaurralde, R.; Smith, S.J.; Clarke, L.E. Integrated Estimates of Global Terrestrial Carbon Sequestration. Glob. Environ. Change 2008, 18, 192–203. [Google Scholar] [CrossRef]
- Archibold, O.W. Temperate Forest Ecosystems. In Ecology of World Vegetation; Springer Science + Business Media: Dordrecht, The Netherlands, 1995; pp. 165–203. ISBN 978-94-010-4008-2. [Google Scholar]
- Martin, P.; Nabuurs, G.-J.; Aubinet, M.; Karjalainen, T.; Vine, E.; Kinsman, J.; Heath, L. Carbon Sinks in Temperate Forests1. Annu. Rev. Energy Environ. 2001, 26, 436–465. [Google Scholar] [CrossRef]
- Pan, Y.; Birdsey, R.; Fang, J.; Houghton, R.; Kauppi, P.; Kurz, W.; Phillips, O.; Shvidenko, A.; Lewis, S.; Canadell, J.; et al. A Large and Persistent Carbon Sink in the World’s Forests. Science 2011, 333, 988–993. [Google Scholar] [CrossRef] [PubMed]
- Xie, J.; Jia, X.; He, G.; Zhou, C.; Yu, H.; Wu, Y.; Bourque, C.P.-A.; Liu, H.; Zha, T. Environmental Control over Seasonal Variation in Carbon Fluxes of an Urban Temperate Forest Ecosystem. Landsc. Urban Plan. 2015, 142, 63–70. [Google Scholar] [CrossRef]
- Allaby, M. Temperate Forests (Biomes of the Earth); AbeBooks: Victoria, BC, Canada, 2006; ISBN 9780816053216. [Google Scholar]
- Luyssaert, S.; Marie, G.; Valade, A.; Chen, Y.-Y.; Njakou Djomo, S.; Ryder, J.; Otto, J.; Naudts, K.; Lansø, A.; Ghattas, J.; et al. Author Correction: Trade-Offs in Using European Forests to Meet Climate Objectives. Nature 2019, 567, E13. [Google Scholar] [CrossRef]
- Vose, J.M.; Peterson, D.L.; Patel-Weynand, T. Effects of Climatic Variability and Change on Forest Ecosystems: A Comprehensive Science Synthesis for the U.S.; General Technical Report PNW-GTR-870; U.S. Department of Agriculture, Forest Service, Pacific Northwest Research Station: Portland, OR, USA, 2012; Volume 870, 265p. [Google Scholar] [CrossRef]
- Kim, J.; Woo, H.R.; Nam, H.G. Toward Systems Understanding of Leaf Senescence: An Integrated Multi-Omics Perspective on Leaf Senescence Research. Mol. Plant 2016, 9, 813–825. [Google Scholar] [CrossRef]
- Li, W.; Wang, L.; He, Z.; Lu, Z.; Cui, J.; Xu, N.; Jin, B.; Wang, L. Physiological and Transcriptomic Changes During Autumn Coloration and Senescence in Ginkgo Biloba Leaves. Hortic. Plant J. 2020, 6, 396–408. [Google Scholar] [CrossRef]
- Wen, C.-H.; Lin, S.-S.; Chu, F.-H. Transcriptome Analysis of a Subtropical Deciduous Tree: Autumn Leaf Senescence Gene Expression Profile of Formosan Gum. Plant Cell Physiol. 2015, 56, 163–174. [Google Scholar] [CrossRef]
- Gao, S.; Zhong, R.; Yan, K.; Ma, X.; Chen, X.; Pu, J.; Gao, S.; Qi, J.; Yin, G.; Myneni, R.B. Evaluating the Saturation Effect of Vegetation Indices in Forests Using 3D Radiative Transfer Simulations and Satellite Observations. Remote Sens. Environ. 2023, 295, 113665. [Google Scholar] [CrossRef]
- Wang, S.; Zhang, L.; Huang, C.; Qiao, N. An NDVI-Based Vegetation Phenology Is Improved to Be More Consistent with Photosynthesis Dynamics through Applying a Light Use Efficiency Model over Boreal High-Latitude Forests. Remote Sens. 2017, 9, 695. [Google Scholar] [CrossRef]
- Frankenberg, C.; Berry, J. 3.10—Solar Induced Chlorophyll Fluorescence: Origins, Relation to Photosynthesis and Retrieval. In Comprehensive Remote Sensing; Liang, S., Ed.; Elsevier: Oxford, UK, 2018; pp. 143–162. ISBN 978-0-12-803221-3. [Google Scholar]
- Zhang, J.; Xiao, J.; Tong, X.; Zhang, J.; Meng, P.; Li, J.; Liu, P.; Yu, P. NIRv and SIF Better Estimate Phenology than NDVI and EVI: Effects of Spring and Autumn Phenology on Ecosystem Production of Planted Forests. Agric. For. Meteorol. 2022, 315, 108819. [Google Scholar] [CrossRef]
- van der Tol, C.; Julitta, T.; Yang, P.; Sabater, N.; Reiter, I.; Tudoroiu, M.; Schuettemeyer, D.; Drusch, M. Retrieval of Chlorophyll Fluorescence from a Large Distance Using Oxygen Absorption Bands. Remote Sens. Environ. 2023, 284, 113304. [Google Scholar] [CrossRef]
- Lelandais, L.; Xueref-Remy, I.; Riandet, A.; Blanc, P.E.; Armengaud, A.; Oppo, S.; Yohia, C.; Ramonet, M.; Delmotte, M. Analysis of 5.5 Years of Atmospheric CO2, CH4, CO Continuous Observations (2014–2020) and Their Correlations, at the Observatoire de Haute Provence, a Station of the ICOS-France National Greenhouse Gases Observation Network. Atmos. Environ. 2022, 277, 119020. [Google Scholar] [CrossRef]
- Xueref-Remy, I.; Milne, M.; Zoghbi, N.; Lelandais, L.; Riandet, A.; Armengaud, A.; Gille, G.; Lanzi, L.; Oppo, S.; Brégonzio-Rozier, L.; et al. Analysis of Atmospheric CO2 Variability in the Marseille City Area and the North-West Mediterranean Basin at Different Time Scales. Atmos. Environ. X 2023, 17, 100208. [Google Scholar] [CrossRef]
- Alonso, L.; Gomez-Chova, L.; Vila-Frances, J.; Amoros-Lopez, J.; Guanter, L.; Calpe, J.; Moreno, J. Improved Fraunhofer Line Discrimination Method for Vegetation Fluorescence Quantification. IEEE Geosci. Remote Sens. Lett. 2008, 5, 620–624. [Google Scholar] [CrossRef]
- Stamford, J.D.; Vialet-Chabrand, S.; Cameron, I.; Lawson, T. Development of an Accurate Low Cost NDVI Imaging System for Assessing Plant Health. Plant Methods 2023, 19, 9. [Google Scholar] [CrossRef]
- Muggeo, V. Segmented: An R Package to Fit Regression Models with Broken-Line Relationships. R News 2008, 8, 20–25. [Google Scholar]
- Campbell, P.K.E.; Huemmrich, K.F.; Middleton, E.M.; Ward, L.A.; Julitta, T.; Daughtry, C.S.T.; Burkart, A.; Russ, A.L.; Kustas, W.P. Diurnal and Seasonal Variations in Chlorophyll Fluorescence Associated with Photosynthesis at Leaf and Canopy Scales. Remote Sens. 2019, 11, 488. [Google Scholar] [CrossRef]
- Wang, F.; Chen, B.; Lin, X.; Zhang, H. Solar-Induced Chlorophyll Fluorescence as an Indicator for Determining the End Date of the Vegetation Growing Season. Ecol. Indic. 2020, 109, 105755. [Google Scholar] [CrossRef]
- Dobrowski, S.Z.; Pushnik, J.C.; Zarco-Tejada, P.J.; Ustin, S.L. Simple Reflectance Indices Track Heat and Water Stress-Induced Changes in Steady-State Chlorophyll Fluorescence at the Canopy Scale. Remote Sens. Environ. 2005, 97, 403–414. [Google Scholar] [CrossRef]
- Garbulsky, M.F.; Filella, I.; Verger, A.; Peñuelas, J. Photosynthetic Light Use Efficiency from Satellite Sensors: From Global to Mediterranean Vegetation. Environ. Exp. Bot. 2014, 103, 3–11. [Google Scholar] [CrossRef]
- Magney, T.S.; Bowling, D.R.; Logan, B.A.; Grossmann, K.; Stutz, J.; Blanken, P.D.; Burns, S.P.; Cheng, R.; Garcia, M.A.; Köhler, P.; et al. Mechanistic Evidence for Tracking the Seasonality of Photosynthesis with Solar-Induced Fluorescence. Proc. Natl. Acad. Sci. USA 2019, 116, 11640–11645. [Google Scholar] [CrossRef] [PubMed]
- Kováč, D.; Novotný, J.; Šigut, L.; Ač, A.; Peñuelas, J.; Grace, J.; Urban, O. Estimation of Photosynthetic Dynamics in Forests from Daily Measured Fluorescence and PRI Data with Adjustment for Canopy Shadow Fraction. Sci. Total Environ. 2023, 898, 166386. [Google Scholar] [CrossRef]
- Chang, C.Y.; Wen, J.; Han, J.; Kira, O.; LeVonne, J.; Melkonian, J.; Riha, S.J.; Skovira, J.; Ng, S.; Gu, L.; et al. Unpacking the Drivers of Diurnal Dynamics of Sun-Induced Chlorophyll Fluorescence (SIF): Canopy Structure, Plant Physiology, Instrument Configuration and Retrieval Methods. Remote Sens. Environ. 2021, 265, 112672. [Google Scholar] [CrossRef]
- Siegmann, B.; Cendrero-Mateo, M.P.; Cogliati, S.; Damm, A.; Gamon, J.; Herrera, D.; Jedmowski, C.; Junker-Frohn, L.V.; Kraska, T.; Muller, O.; et al. Downscaling of Far-Red Solar-Induced Chlorophyll Fluorescence of Different Crops from Canopy to Leaf Level Using a Diurnal Data Set Acquired by the Airborne Imaging Spectrometer HyPlant. Remote Sens. Environ. 2021, 264, 112609. [Google Scholar] [CrossRef]
- Yang, X.; Tang, J.; Mustard, J.F.; Lee, J.-E.; Rossini, M.; Joiner, J.; Munger, J.W.; Kornfeld, A.; Richardson, A.D. Solar-Induced Chlorophyll Fluorescence That Correlates with Canopy Photosynthesis on Diurnal and Seasonal Scales in a Temperate Deciduous Forest: Fluorescence and Photosynthesis. Geophys. Res. Lett. 2015, 42, 2977–2987. [Google Scholar] [CrossRef]
- Magney, T.S.; Frankenberg, C.; Köhler, P.; North, G.; Davis, T.S.; Dold, C.; Dutta, D.; Fisher, J.B.; Grossmann, K.; Harrington, A.; et al. Disentangling Changes in the Spectral Shape of Chlorophyll Fluorescence: Implications for Remote Sensing of Photosynthesis. J. Geophys. Res. Biogeosciences 2019, 124, 1491–1507. [Google Scholar] [CrossRef]
- Wu, L.; Zhang, Y.; Zhang, Z.; Zhang, X.; Wu, Y.; Chen, J.M. Deriving Photosystem-Level Red Chlorophyll Fluorescence Emission by Combining Leaf Chlorophyll Content and Canopy Far-Red Solar-Induced Fluorescence: Possibilities and Challenges. Remote Sens. Environ. 2024, 304, 114043. [Google Scholar] [CrossRef]
- Martínez-Ferri, E.; Balaguer, L.; Valladares, F.; Chico, J.M.; Manrique, E. Energy Dissipation in Drought-Avoiding and Drought-Tolerant Tree Species at Midday during the Mediterranean Summer. Tree Physiol. 2000, 20, 131–138. [Google Scholar] [CrossRef]
- Pons, T.; Welschen, R. Midday Depression of Net Photosynthesis in the Tropical Rainforest Tree Eperua Grandiflora: Contributions of Stomatal and Internal Conductances, Respiration and Rubisco Functioning. Tree Physiol. 2003, 23, 937–947. [Google Scholar] [CrossRef]
- Deng, Z.; Chen, J.; Wang, S.; Li, T.; Huang, K.; Gu, P.; Peng, H.; Chen, Z. Response of Vegetation Photosynthesis to the 2022 Drought in Yangtze River Basin by Diurnal Orbiting Carbon Observatory-2/3 Satellite Observations. J. Remote Sens. 2025, 5, 0445. [Google Scholar] [CrossRef]
- Belviso, S.; Reiter, I.M.; Loubet, B.; Gros, V.; Lathière, J.; Montagne, D.; Delmotte, M.; Ramonet, M.; Kalogridis, C.; Lebegue, B.; et al. A Top-down Approach of Surface Carbonyl Sulfide Exchange by a Mediterranean Oak Forest Ecosystem in Southern France. Atmos. Chem. Phys. 2016, 16, 14909–14923. [Google Scholar] [CrossRef]
- Paul-Limoges, E.; Damm, A.; Hueni, A.; Liebisch, F.; Eugster, W.; Schaepman, M.E.; Buchmann, N. Effect of Environmental Conditions on Sun-Induced Fluorescence in a Mixed Forest and a Cropland. Remote Sens. Environ. 2018, 219, 310–323. [Google Scholar] [CrossRef]
- Biriukova, K.; Celesti, M.; Evdokimov, A.; Pacheco-Labrador, J.; Julitta, T.; Migliavacca, M.; Giardino, C.; Miglietta, F.; Colombo, R.; Panigada, C.; et al. Effects of Varying Solar-View Geometry and Canopy Structure on Solar-Induced Chlorophyll Fluorescence and PRI. Int. J. Appl. Earth Obs. Geoinf. 2020, 89, 102069. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, W.; Wu, J.; Liu, S.; Teng, Y.; Yang, J.; Han, X. The Impacts of Growth and Environmental Parameters on Solar-Induced Chlorophyll Fluorescence at Seasonal and Diurnal Scales. Remote Sens. 2019, 11, 2002. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Baulard, A.; Mevy, J.-P.; Xueref-Remy, I.; Reiter, I.M.; Julitta, T.; Miglietta, F. Solar-Induced Fluorescence as Indicator of Downy Oak and the Influence of Some Environmental Variables at the End of the Growing Season. Remote Sens. 2025, 17, 1252. https://doi.org/10.3390/rs17071252
Baulard A, Mevy J-P, Xueref-Remy I, Reiter IM, Julitta T, Miglietta F. Solar-Induced Fluorescence as Indicator of Downy Oak and the Influence of Some Environmental Variables at the End of the Growing Season. Remote Sensing. 2025; 17(7):1252. https://doi.org/10.3390/rs17071252
Chicago/Turabian StyleBaulard, Antoine, Jean-Philippe Mevy, Irène Xueref-Remy, Ilja Marco Reiter, Tommaso Julitta, and Franco Miglietta. 2025. "Solar-Induced Fluorescence as Indicator of Downy Oak and the Influence of Some Environmental Variables at the End of the Growing Season" Remote Sensing 17, no. 7: 1252. https://doi.org/10.3390/rs17071252
APA StyleBaulard, A., Mevy, J.-P., Xueref-Remy, I., Reiter, I. M., Julitta, T., & Miglietta, F. (2025). Solar-Induced Fluorescence as Indicator of Downy Oak and the Influence of Some Environmental Variables at the End of the Growing Season. Remote Sensing, 17(7), 1252. https://doi.org/10.3390/rs17071252