

Unraveling Phenological Dynamics: Exploring Early Springs, Late Autumns, and Climate Drivers Across Different Vegetation Types in Northeast China




Abstract

1. Introduction
2. Materials and Methods
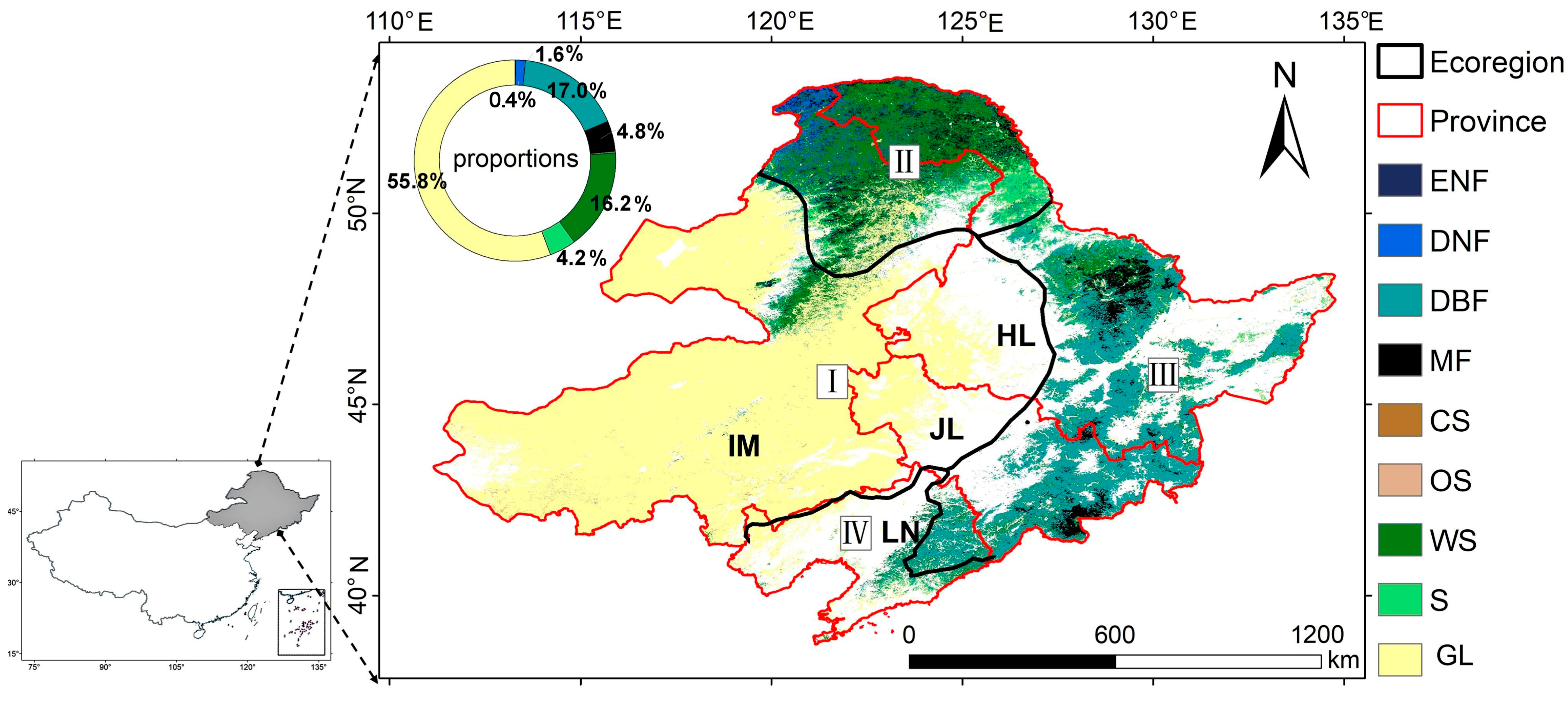
2.1. Study Area
2.2. Remotely Sensed Data
2.3. Ground-Based Phenological Observation Data
2.4. Methodology
2.4.1. The Extraction of Phenology Indicators Based on 4-Day kNDVI Sequence
2.4.2. Trend Analysis of LSP Indicators
- (1)
- Significantly delayed or extended (slope > 0 and |Z| > 1.96);
- (2)
- Slightly delayed or extended (slope > 0 and |Z| ≤ 1.96);
- (3)
- Significantly advanced or shortened (slope < 0 and |Z| > 1.96);
- (4)
- Slightly advanced or shortened (slope < 0 and |Z| ≤ 1.96);
- (5)
- No significant change (slope = 0).
2.4.3. Response of LSP to Climate Factors
- Evaluation of the importance of seasonal climate variables
- Correlation analysis
3. Results
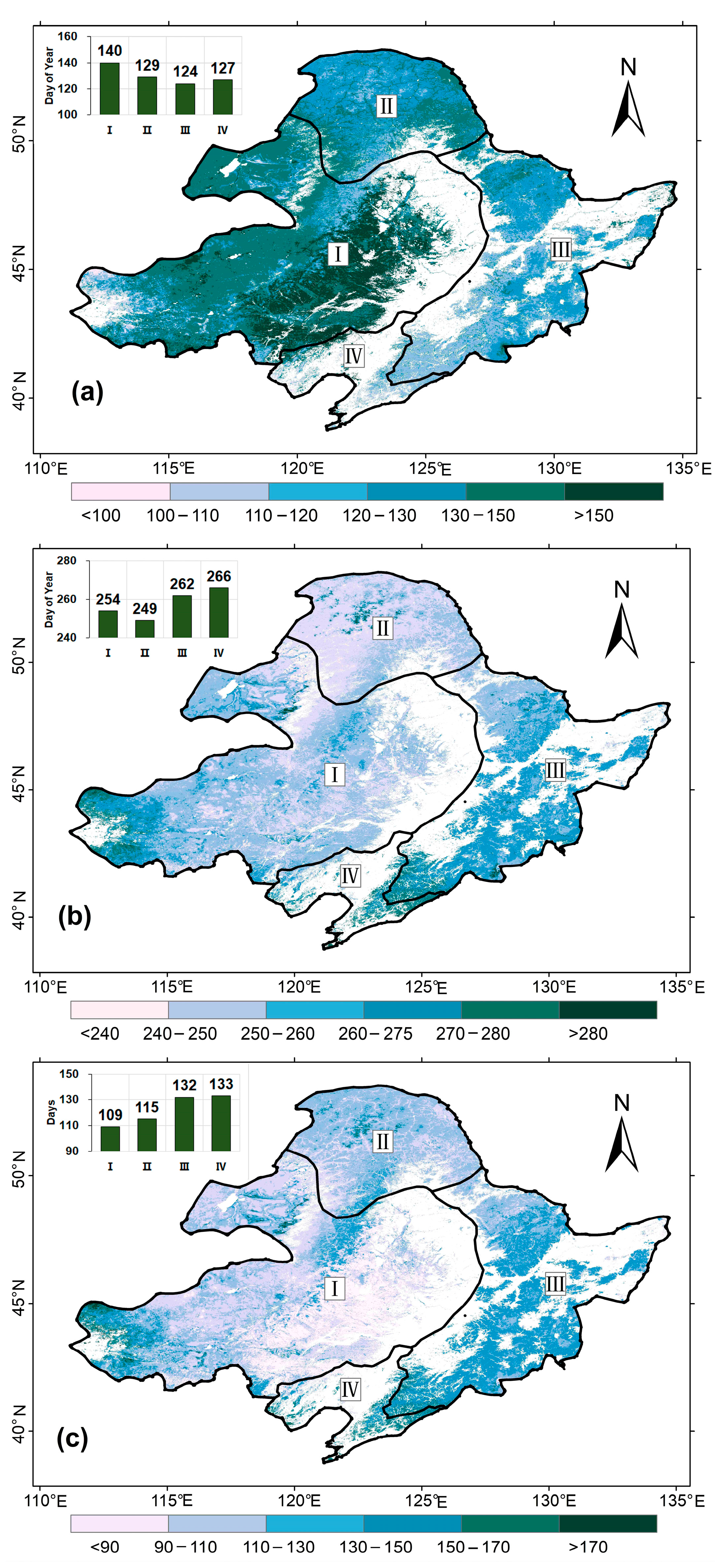
3.1. The Extraction of LSP Indicators Using Optimal Dynamic Thresholds
3.2. Spatiotemporal Trend of the LSP Indicators in the NEC
3.3. Response of LSP to Climate Factors
3.3.1. Gradient Relationship Between Elevation and LSP
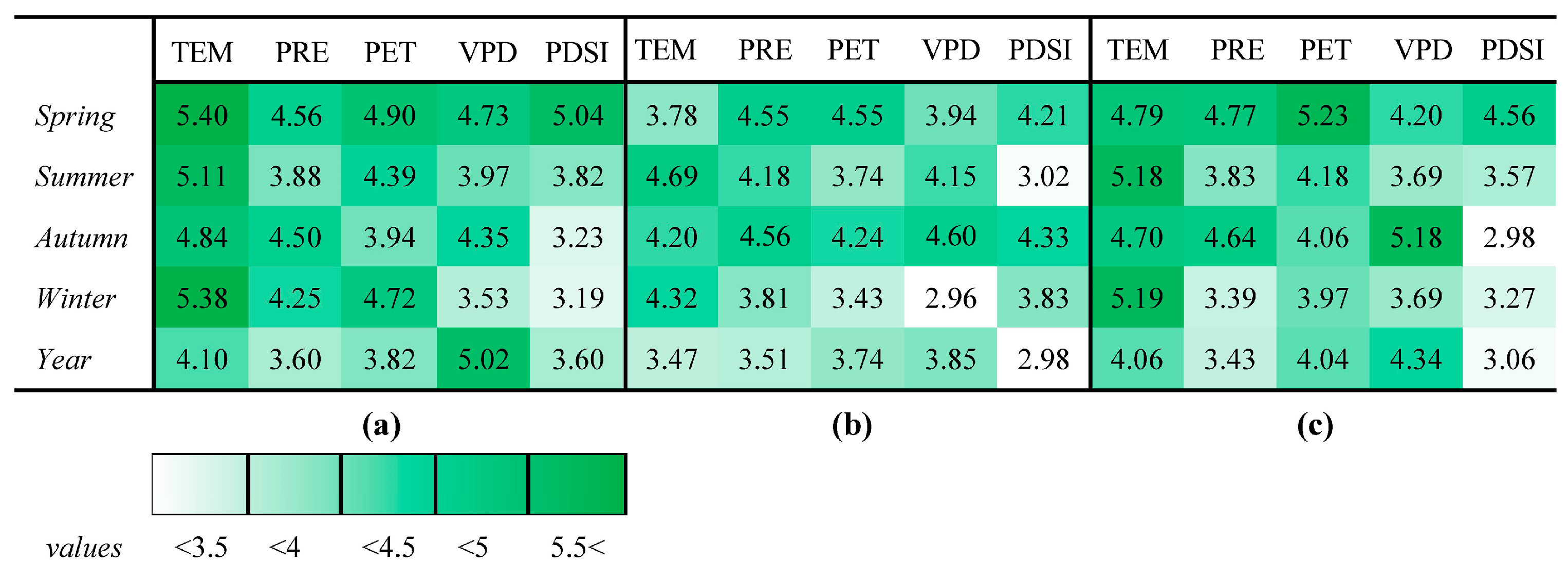
3.3.2. Evaluation of the Importance of Seasonal Climate Variables
3.3.3. Correlation Analysis
4. Discussion
4.1. Spatial Pattern of LSP in the NEC
4.2. Trend of LSP in the NEC
4.3. Response of LSP to Climate Factors
4.4. Limitations and Future Work
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lieth, H. Phenology in Productivity Studies. In Analysis of Temperate Forest Ecosystems; Reichle, D.E., Ed.; Springer: Berlin/Heidelberg, Germany, 1973; pp. 29–46. [Google Scholar] [CrossRef]
- Roetzer, T.; Wittenzeller, M.; Haeckel, H.; Nekovar, J. Phenology in Central Europe—Differences and Trends of Spring Phenophases in Urban and Rural Areas. Int. J. Biometeorol. 2000, 44, 60–66. [Google Scholar] [CrossRef] [PubMed]
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate Change, Phenology, and Phenological Control of Vegetation Feedbacks to the Climate System. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- de Beurs, K.M.; Henebry, G.M. Land Surface Phenology, Climatic Variation, and Institutional Change: Analyzing Agricultural Land Cover Change in Kazakhstan. Remote Sens. Environ. 2004, 89, 497–509. [Google Scholar] [CrossRef]
- Xu, X.; Zhou, G.; Du, H.; Mao, F.; Xu, L.; Li, X.; Liu, L. Combined MODIS Land Surface Temperature and Greenness Data for Modeling Vegetation Phenology, Physiology, and Gross Primary Production in Terrestrial Ecosystems. Sci. Total Environ. 2020, 726, 137948. [Google Scholar] [CrossRef]
- Ma, X.; Zhu, X.; Xie, Q.; Jin, J.; Zhou, Y.; Luo, Y.; Liu, Y.; Tian, J.; Zhao, Y. Monitoring Nature’s Calendar from Space: Emerging Topics in Land Surface Phenology and Associated Opportunities for Science Applications. Glob. Change Biol. 2022, 28, 7186–7204. [Google Scholar] [CrossRef]
- Post, E.; Stenseth, N.C. Climatic Variability, Plant Phenology, and Northern Ungulates. Ecology 1999, 80, 1322–1339. [Google Scholar] [CrossRef]
- Katal, N.; Rzanny, M.; Mäder, P.; Wäldchen, J. Deep Learning in Plant Phenological Research: A Systematic Literature Review. Front. Plant Sci. 2022, 13, 805738. [Google Scholar] [CrossRef]
- Zhong, R.; Yan, K.; Gao, S.; Yang, K.; Zhao, S.; Ma, X.; Zhu, P.; Fan, L.; Yin, G. Response of Grassland Growing Season Length to Extreme Climatic Events on the Qinghai-Tibetan Plateau. Sci. Total Environ. 2024, 909, 168488. [Google Scholar] [CrossRef]
- Jönsson, P.; Eklundh, L. TIMESAT—A Program for Analyzing Time-Series of Satellite Sensor Data. Comput. Geosci. 2004, 30, 833–845. [Google Scholar] [CrossRef]
- Chang, Q.; Xiao, X.; Jiao, W.; Wu, X.; Doughty, R.; Wang, J.; Du, L.; Zou, Z.; Qin, Y. Assessing Consistency of Spring Phenology of Snow-Covered Forests as Estimated by Vegetation Indices, Gross Primary Production, and Solar-Induced Chlorophyll Fluorescence. Agric. For. Meteorol. 2019, 275, 305–316. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the Radiometric and Biophysical Performance of the MODIS Vegetation Indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Myneni, R.B.; Hoffman, S.; Knyazikhin, Y.; Privette, J.L.; Glassy, J.; Tian, Y.; Wang, Y.; Song, X.; Zhang, Y.; Smith, G.R.; et al. Global Products of Vegetation Leaf Area and Fraction Absorbed PAR from Year One of MODIS Data. Remote Sens. Environ. 2002, 83, 214–231. [Google Scholar] [CrossRef]
- Tucker, C.J. Red and Photographic Infrared Linear Combinations for Monitoring Vegetation. Remote Sens. Environ. 1979, 8, 127–150. [Google Scholar] [CrossRef]
- Zhou, L.; Zhou, W.; Chen, J.; Xu, X.; Wang, Y.; Zhuang, J.; Chi, Y. Land Surface Phenology Detections from Multi-Source Remote Sensing Indices Capturing Canopy Photosynthesis Phenology across Major Land Cover Types in the Northern Hemisphere. Ecol. Indic. 2022, 135, 108579. [Google Scholar] [CrossRef]
- Wang, Q.; Moreno-Martínez, Á.; Muñoz-Marí, J.; Campos-Taberner, M.; Camps-Valls, G. Estimation of Vegetation Traits with Kernel NDVI. ISPRS J. Photogramm. Remote Sens. 2023, 195, 408–417. [Google Scholar] [CrossRef]
- Zeng, L.; Wardlow, B.D.; Xiang, D.; Hu, S.; Li, D. A Review of Vegetation Phenological Metrics Extraction Using Time-Series, Multispectral Satellite Data. Remote Sens. Environ. 2020, 237, 111511. [Google Scholar] [CrossRef]
- Xia, C. Review of Advances in Vegetation Phenology Monitoring by Remote Sensing. Natl. Remote Sens. Bull. 2013, 17, 1–16. [Google Scholar] [CrossRef]
- Puchi, P.F.; Castagneri, D.; Rossi, S.; Carrer, M. Wood Anatomical Traits in Black Spruce Reveal Latent Water Constraints on the Boreal Forest. Glob. Change Biol. 2020, 26, 1767–1777. [Google Scholar] [CrossRef]
- Wang, J.; Meng, S.; Zhu, W.; Xu, Z. Phenological Changes and Their Influencing Factors under the Joint Action of Water and Temperature in Northeast Asia. Remote Sens. 2023, 15, 5298. [Google Scholar] [CrossRef]
- Yang, Y.; Fan, F. Land Surface Phenology and Its Response to Climate Change in the Guangdong-Hong Kong-Macao Greater Bay Area during 2001–2020. Ecol. Indic. 2023, 154, 110728. [Google Scholar] [CrossRef]
- Tao, Z.; Wang, H.; Liu, Y.; Xu, Y.; Dai, J. Phenological Response of Different Vegetation Types to Temperature and Precipitation Variations in Northern China during 1982–2012. Int. J. Remote Sens. 2017, 38, 3236–3252. [Google Scholar] [CrossRef]
- Feng, Z.; Chen, J.; Huang, R.; Yang, Y.; You, H.; Han, X. Spatial and Temporal Variation in Alpine Vegetation Phenology and Its Response to Climatic and Topographic Factors on the Qinghai-Tibet Plateau. Sustainability 2022, 14, 12802. [Google Scholar] [CrossRef]
- Fang, X.; Zhu, Q.; Chen, H.; Ma, Z.; Wang, W.; Song, X.; Zhao, P.; Peng, C. Analysis of Vegetation Dynamics and Climatic Variability Impacts on Greenness across Canada Using Remotely Sensed Data from 2000 to 2009. J. Appl. Remote Sens. 2014, 8, 083666. [Google Scholar] [CrossRef]
- Gaertner, B.A.; Zegre, N.; Warner, T.; Fernandez, R.; He, Y.; Merriamb, E.R. Climate, Forest Growing Season, and Evapotranspiration Changes in the Central Appalachian Mountains, USA. Sci. Total Environ. 2019, 650, 1371–1381. [Google Scholar] [CrossRef]
- Guo, S.; Bai, H.; Huang, X.; Meng, Q.; Zhao, T. Remote Sensing Phenology of Larix chinensis Forest in Response to Climate Change in Qinling Mountains. Chin. J. Ecol. 2019, 38, 1123–1132. Available online: https://www.cje.net.cn/CN/Y2019/V38/I4/1123 (accessed on 15 March 2024). (In Chinese).
- Kovalskyy, V.; Henebry, G.M. Alternative Methods to Predict Actual Evapotranspiration Illustrate the Importance of Accounting for Phenology—Part 2: The Event Driven Phenology Model. Biogeosciences 2012, 9, 161–177. [Google Scholar] [CrossRef]
- Huang, J.-G.; Zhang, Y.; Wang, M.; Yu, X.; Deslauriers, A.; Fonti, P.; Liang, E.; Makinen, H.; Oberhuber, W.; Rathgeber, C.B.K.; et al. A Critical Thermal Transition Driving Spring Phenology of Northern Hemisphere Conifers. Glob. Change Biol. 2023, 29, 1606–1617. [Google Scholar] [CrossRef]
- Ren, S.; Li, Y.; Peichl, M. Diverse Effects of Climate at Different Times on Grassland Phenology in Mid-Latitude of the Northern Hemisphere. Ecol. Indic. 2020, 113, 106260. [Google Scholar] [CrossRef]
- Hou, X.; Niu, Z.; Gao, S. Phenology of Forest Vegetation in Northeast of China in Ten Years Using Remote Sensing. Spectrosc. Spectr. Anal. 2014, 34, 515–519. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, L.; Wang, H.; Zhang, Y.; Wang, N.; Chen, J. Land Surface Phenology of Northeast China during 2000–2015: Temporal Changes and Relationships with Climate Changes. Environ. Monit. Assess. 2017, 189, 531. [Google Scholar] [CrossRef]
- Yu, X.; Wang, Q.; Yan, H.; Wang, Y.; Wen, K.; Zhuang, D.; Wang, Q. Forest Phenology Dynamics and Its Responses to Meteorological Variations in Northeast China. Adv. Meteorol. 2014, 2014, 592106. [Google Scholar] [CrossRef]
- Yuan, M.; Zhao, L.; Lin, A.; Wang, L.; Li, Q.; She, D.; Qu, S. Impacts of Preseason Drought on Vegetation Spring Phenology across the Northeast China Transect. Sci. Total Environ. 2020, 738, 140297. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Jiang, Q.; Deng, X.; Lv, K.; Zhang, Z. Spatio-Temporal Evolution, Future Trend and Phenology Regularity of Net Primary Productivity of Forests in Northeast China. Remote Sens. 2020, 12, 3670. [Google Scholar] [CrossRef]
- Shen, X.; Liu, B.; Henderson, M.; Wang, L.; Jiang, M.; Lu, X. Vegetation Greening, Extended Growing Seasons, and Temperature Feedbacks in Warming Temperate Grasslands of China. J. Clim. 2022, 35, 5103–5117. [Google Scholar] [CrossRef]
- Wu, R.; Zhang, H.; Zhao, J.; Yu, S.; Guo, X.; Hong, Y.; Deng, G.; Li, H. Promote the Advance of the Start of the Growing Season from Combined Effects of Climate Change and Wildfire. Ecol. Indic. 2021, 125, 107483. [Google Scholar] [CrossRef]
- Meng, F.; Zhou, Y.; Cui, S.; Wang, Q.; Tsechoe, D.; Wang, S. Effects of Climate Changes on Plant Phenology at High-Latitude and Alpine Regions. J. Univ. Chin. Acad. Sci. 2017, 34, 498–507. Available online: http://journal.ucas.ac.cn/EN/10.7523/j.issn.2095-6134.2017.04.012 (accessed on 15 March 2024).
- Wang, J.; Zhou, T.; Peng, P. Phenology Response to Climatic Dynamic across China’s Grasslands from 1985 to 2010. ISPRS Int. J. Geo-Inf. 2018, 7, 290. [Google Scholar] [CrossRef]
- Su, Y.; Guo, Q.; Hu, T.; Guan, H.; Jin, S.; An, S.; Chen, X.; Guo, K.; Hao, Z.; Hu, Y.; et al. An Updated Vegetation Map of China (1:1000000). Sci. Bull. 2020, 65, 1125–1136. [Google Scholar] [CrossRef]
- Friedl, M.; Sulla-Menashe, D. MODIS/Terra+Aqua Land Cover Type Yearly L3 Global 500m SIN Grid V061. NASA LP DAAC. 2022. Available online: https://lpdaac.usgs.gov/products/mcd12q1v061/ (accessed on 15 March 2024).
- Peng, S. 1 km Monthly Potential Evapotranspiration Dataset for China (1901–2023). National Tibetan Plateau/Third Pole Environment Data Center. 2022. Available online: https://data.tpdc.ac.cn/en/data/8b11da09-1a40-4014-bd3d-2b86e6dccad4 (accessed on 15 March 2024).
- Peng, S. 1-km Monthly Precipitation Dataset for China (1901–2023). National Tibetan Plateau/Third Pole Environment Data Center. 2020. Available online: https://data.tpdc.ac.cn/en/data/faae7605-a0f2-4d18-b28f-5cee413766a2 (accessed on 15 March 2024).
- Peng, S. 1-km Monthly Mean Temperature Dataset for China (1901–2023). National Tibetan Plateau/Third Pole Environment Data Center. 2019. Available online: https://data.tpdc.ac.cn/en/data/71ab4677-b66c-4fd1-a004-b2a541c4d5bf/ (accessed on 15 March 2024).
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a High-Resolution Global Dataset of Monthly Climate and Climatic Water Balance from 1958–2015. Sci. Data 2018, 5, 170191. [Google Scholar] [CrossRef]
- Berra, E.F.; Gaulton, R. Remote Sensing of Temperate and Boreal Forest Phenology: A Review of Progress, Challenges and Opportunities in the Intercomparison of In-Situ and Satellite Phenological Metrics. For. Ecol. Manag. 2021, 480, 118663. [Google Scholar] [CrossRef]
- Savitzky, A.; Golay, M.J.E. Smoothing and Differentiation of Data by Simplified Least Squares Procedures. Anal. Chem. 1964, 36, 1627–1639. [Google Scholar] [CrossRef]
- Li, J.; Tang, Z.; Deng, G.; Sang, G.; Wang, J. Remote Sensing Monitoring of Grassland Phenological Changes in the Qinghai-Tibetan Plateau During 2001–2020. Res. Soil Water Conserv. 2023, 30, 265–274. (In Chinese) [Google Scholar] [CrossRef]
- Tian, Y. Multi-Scale Study on the Response of Vegetation Phenology to Climate Change in the Heilongjiang Basin. Ph.D. Thesis, University of Chinese Academy of Sciences (Northeast Institute of Geography and Agroecology, Chinese Academy of Sciences), Changchun, China, 2020. (In Chinese) [Google Scholar] [CrossRef]
- Tian, Y.; Bai, X.; Wang, S.; Qin, L.; Li, Y. Spatial-Temporal Changes of Vegetation Cover in Guizhou Province, Southern China. Chin. Geogr. Sci. 2017, 27, 25–38. [Google Scholar] [CrossRef]
- Genuer, R.; Poggi, J.; Tuleau-Malot, C. Variable Selection Using Random Forests. Pattern Recognit. Lett. 2010, 31, 2225–2236. [Google Scholar] [CrossRef]
- Philipp, M.; Wegmann, M.; Kubert-Flock, C. Quantifying the Response of German Forests to Drought Events via Satellite Imagery. Remote Sens. 2021, 13, 1845. [Google Scholar] [CrossRef]
- Sedgwick, P. Statistical Question: Spearman’s Rank Correlation Coefficient. BMJ-Br. Med. J. 2014, 349, g7327. [Google Scholar] [CrossRef]
- Gao, M.; Wang, X.; Meng, F.; Liu, Q.; Li, X.; Zhang, Y.; Piao, S. Three-dimensional change in temperature sensitivity of northern vegetation phenology. Glob. Change Biol. 2020, 26, 5189–5201. [Google Scholar] [CrossRef]
- Hopkins, A.D. The Bioclimatic Law. J. Wash. Acad. Sci. 1920, 10, 34–40. Available online: https://www.jstor.org/stable/24521154 (accessed on 15 March 2024).
- Liu, X.; Zhu, X.; Zhu, W.; Pan, Y.; Zhang, C.; Zhang, D. Changes in Spring Phenology in the Three-Rivers Headwater Region from 1999 to 2013. Remote Sens. 2014, 6, 9130–9144. [Google Scholar] [CrossRef]
- Yu, X.; Zhuang, D. Monitoring Forest Phenophases of Northeast China based on MODIS NDVI Data. Resour. Sci. 2006, 28, 111–117. Available online: https://www.resci.cn/CN/Y2006/V28/I4/111 (accessed on 15 March 2024). (In Chinese).
- Wei, X.; Xu, M.; Zhao, H.; Liu, X.; Guo, Z.; Li, X.; Zha, T. Exploring Sensitivity of Phenology to Seasonal Climate Differences in Temperate Grasslands of China Based on Normalized Difference Vegetation Index. Land 2024, 13, 399. [Google Scholar] [CrossRef]
- Pellerin, M.; Delestrade, A.; Mathieu, G.; Rigault, O.; Yoccoz, N.G. Spring tree phenology in the Alps: Effects of air temperature, altitude and local topography. Eur. J. For. Res. 2012, 131, 1957–1965. [Google Scholar] [CrossRef]
- Shen, M.; Zhang, G.; Cong, N.; Wang, S.; Kong, W.; Piao, S. Increasing altitudinal gradient of spring vegetation phenology during the last decade on the Qinghai-Tibetan Plateau. Agric. For. Meteorol. 2014, 189, 71–80. [Google Scholar] [CrossRef]
- Chang, S.; He, H.S.; Huang, F.; Krohn, J. Spring temperature and snow cover co-regulate variations of forest phenology in Changbai Mountains, Northeast China. Eur. J. For. Res. 2023, 143, 547–560. [Google Scholar] [CrossRef]
- Zhang, Y.; Yin, P.; Li, X.; Niu, Q.; Wang, Y.; Cao, W.; Huang, J.; Chen, H.; Yao, X.; Yu, L.; et al. The divergent response of vegetation phenology to urbanization: A case study of Beijing city, China. Sci. Total Environ. 2022, 803, 150079. [Google Scholar] [CrossRef]
- Zhao, J.; Wang, Y.; Zhang, Z.; Zhang, H.; Guo, X.; Yu, S.; Du, W.; Huang, F. The Variations of Land Surface Phenology in Northeast China and Its Responses to Climate Change from 1982 to 2013. Remote Sens. 2016, 8, 400. [Google Scholar] [CrossRef]
- Sakai, A.; Larcher, W. Frost Survival of Plants: Responses and Adaptation to Freezing Stress; Springer Science and Business Media: Berlin/Heidelberg, Germany, 2012; Available online: https://link.springer.com/book/10.1007/978-3-642-71745-1 (accessed on 15 March 2024).
- Tranquillini, W. Physiological Ecology of the Alpine Timberline: Tree Existence at High Altitudes with Special Reference to the European Alps; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2012; Available online: https://link.springer.com/book/10.1007/978-3-642-67107-4 (accessed on 15 March 2024).
- Zhai, P.; Pan, X. Change in Extreme Temperature and Precipitation over Northern China During the Second Half of the 20th Century. Acta Geogr. Sin. 2003, 58, 1–10. [Google Scholar] [CrossRef]
- Liu, Y.Y.; van Dijk, A.I.J.M.; de Jeu, R.A.M.; Canadell, J.G.; McCabe, M.F.; Evans, J.P.; Wang, G. Recent reversal in loss of global terrestrial biomass. Nat. Clim. Chang. 2015, 5, 470–474. [Google Scholar] [CrossRef]
- Cook, B.; Smerdon, J.; Seager, R.; Coats, S. Global warming and 21st century drying. Clim. Dynam. 2014, 43, 2607–2627. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Y. Impacts of climate, phenology, elevation and their interactions on the net primary productivity of vegetation in Yunnan, China under global warming. Ecol. Indic. 2023, 154, 110533. [Google Scholar] [CrossRef]
- Ren, S.; Peichl, M. Enhanced spatiotemporal heterogeneity and the climatic and biotic controls of autumn phenology in northern grasslands. Sci. Total Environ. 2021, 788, 147806. [Google Scholar] [CrossRef] [PubMed]
- Dronova, I.; Taddeo, S. Remote sensing of phenology: Towards the comprehensive indicators of plant community dynamics from species to regional scales. J. Ecol. 2022, 110, 1460–1484. [Google Scholar] [CrossRef]
- Mayer, A. Phenology and Citizen Science: Volunteers have documented seasonal events for more than a century, and scientific studies are benefiting from the data. Bioscience 2010, 60, 172–175. [Google Scholar] [CrossRef]
- Richardson, A.D.; Hufkens, K.; Milliman, T.; Frolking, S. Intercomparison of phenological transition dates derived from the PhenoCam Dataset V1.0 and MODIS satellite remote sensing. Sci. Rep. 2018, 8, 5679. [Google Scholar] [CrossRef]
- Templ, B.; Koch, E.; Bolmgren, K.; Ungersböck, M.; Paul, A.; Scheifinger, H.; Rutishauser, T.; Busto, M.; Chmielewski, F.-M.; Hájková, L.; et al. Pan European Phenological database (PEP725): A single point of access for European data. Int. J. Biometeorol. 2018, 62, 1109–1113. [Google Scholar] [CrossRef]
- Wheeler, K.I.; Dietze, M.C. Improving the monitoring of deciduous broadleaf phenology using the Geostationary Operational Environmental Satellite (GOES) 16 and 17. Biogeosciences 2021, 18, 1971–1985. [Google Scholar] [CrossRef]
- Wei, B.; Wei, J.; Jia, X.; Ye, Z.; Yu, S.; Yin, S. Spatiotemporal Patterns of Land Surface Phenology from 2001 to 2021 in the Agricultural Pastoral Ecotone of Northern China. Sustainability 2023, 15, 5830. [Google Scholar] [CrossRef]
- Testa, S.; Mondino, E.C.B.; Pedroli, C. Correcting MODIS 16-day composite NDVI time-series with actual acquisition dates. Eur. J. Remote Sens. 2014, 47, 285–305. [Google Scholar] [CrossRef]
- Zheng, W.; Liu, Y.; Yang, X.; Fan, W. Spatiotemporal Variations of Forest Vegetation Phenology and Its Response to Climate Change in Northeast China. Remote Sens. 2022, 14, 2909. [Google Scholar] [CrossRef]
- Shen, X.; Shen, M.; Wu, C.; Peñuelas, J.; Ciais, P.; Zhang, J.; Freeman, C.; Palmer, P.I.; Liu, B.; Henderson, M.; et al. Critical role of water conditions in the responses of autumn phenology of marsh wetlands to climate change on the Tibetan Plateau. Glob. Change Biol. 2023, 30, e17097. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Description | Units | Spatial Resolution |
|---|---|---|---|
| TEM | Temperature | °C | 1 km |
| PRE | Precipitation | mm | 1 km |
| PET | Potential evapotranspiration | mm | 1 km |
| VPD | Vapor pressure deficit | kPa | 4 km |
| PDSI | Palmer drought severity index | * | 4 km |
| Year | City | Province | Latitude | Longitude | SOS | EOS | LOS | Vegetation Types |
|---|---|---|---|---|---|---|---|---|
| 1974–1996 | HH | HL | 50.25°N | 127.50°E | 140 | 269 | 130 | woody |
| 1966–1996 | JMS | HL | 46.81°N | 130.37°E | 129 | 283 | 154 | woody |
| 1963–2012 | HRB | HL | 45.77°N | 126.64°E | 124 | 275 | 137 | woody |
| 1964–1996 | MDJ | HL | 44.58°N | 129.62°E | 125 | 261 | 138 | woody |
| 2014 | HHHT | IM East | 43.93°N | 116.05°E | 100 | 289 | 192 | herbaceous |
| 1986–2012 | CC | JL | 43.82°N | 125.32°E | 121 | 267 | 147 | woody |
| 1964–2012 | SY | LN | 41.81°N | 123.43°E | 116 | 261 | 146 | woody |
| Variables | SOS | EOS | LOS |
|---|---|---|---|
| TEM | Spring | Summer | Summer |
| PRE | Spring | Autumn | Spring |
| PET | Spring | Spring | Spring |
| VPD | Year | Autumn | Autumn |
| PDSI | Spring | Autumn | Spring |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, J.; Zou, H.; Zhao, Y.; Wang, X.; Zhen, Z. Unraveling Phenological Dynamics: Exploring Early Springs, Late Autumns, and Climate Drivers Across Different Vegetation Types in Northeast China. Remote Sens. 2025, 17, 1853. https://doi.org/10.3390/rs17111853
Liu J, Zou H, Zhao Y, Wang X, Zhen Z. Unraveling Phenological Dynamics: Exploring Early Springs, Late Autumns, and Climate Drivers Across Different Vegetation Types in Northeast China. Remote Sensing. 2025; 17(11):1853. https://doi.org/10.3390/rs17111853
Chicago/Turabian StyleLiu, Jiayu, Haifeng Zou, Yinghui Zhao, Xiaochun Wang, and Zhen Zhen. 2025. "Unraveling Phenological Dynamics: Exploring Early Springs, Late Autumns, and Climate Drivers Across Different Vegetation Types in Northeast China" Remote Sensing 17, no. 11: 1853. https://doi.org/10.3390/rs17111853
APA StyleLiu, J., Zou, H., Zhao, Y., Wang, X., & Zhen, Z. (2025). Unraveling Phenological Dynamics: Exploring Early Springs, Late Autumns, and Climate Drivers Across Different Vegetation Types in Northeast China. Remote Sensing, 17(11), 1853. https://doi.org/10.3390/rs17111853