
Mapping and Monitoring of the Invasive Species Dichrostachys cinerea (Marabú) in Central Cuba Using Landsat Imagery and Machine Learning (1994–2022)

 ,
, 


Abstract

1. Introduction
1.1. Invasive Plant Species in Cuba: The Case of Dichrostachys cinerea (L.)
1.2. Remote Sensing for Invasive Species Monitoring
1.3. Objective and Aims of This Work
2. Materials and Methods
2.1. Study Area
2.2. Satellite Data
2.3. Field Reference Data
2.4. Classification of Satellite Imagery
2.5. Validation
2.6. Classifier Choice
2.7. Land Use and Land Cover Change Analysis
3. Results
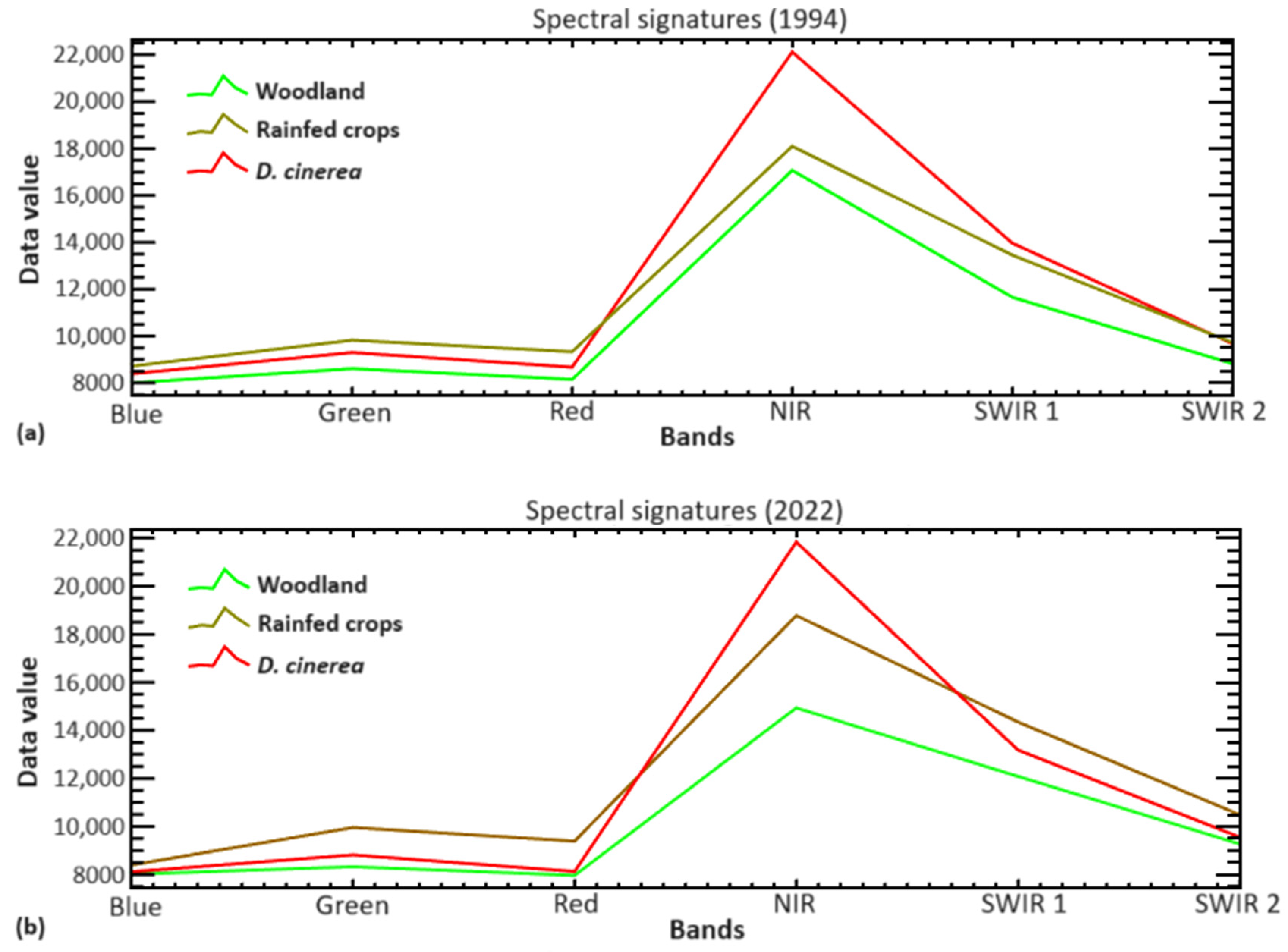
3.1. Spectral Characterization of D. cinerea
3.2. Dichrostachys cinerea Detection with Landsat 5 TM and Landsat 8 OLI images
3.3. Dichrostachys cinerea Spread from 1994 to 2022
3.4. Marabú-Induced Changes in other LULCs
4. Discussion
4.1. Dichrostachys cinerea Detection Using Remote Sensing
4.2. Dichrostachys cinerea Spread from 1994 to 2022 and Land Cover Changes
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubyna, D.V.; Dziuba, T.P.; Iemelianova, S.M.; Protopopova, V.V.; Shevera, M.V. Alien Species in the Pioneer and Ruderal Vegetation of Ukraine. Diversity 2022, 14, 1085. [Google Scholar] [CrossRef]
- Espínola, L.A.; Júlio Junior, H.F. Espécies invasoras: Conceitos, modelos e atributos. Interciencia 2007, 32, 580–585. [Google Scholar]
- Álvarez-Taboada, F.; Paredes, C.; Julián-Pelaz, J. Mapping of the invasive species Hakea sericea using Unmanned Aerial Vehicle (UAV) and Worldview-2 imagery and an object-oriented approach. Remote Sens. 2017, 9, 913. [Google Scholar] [CrossRef]
- de Francesco, M.C.; Tozzi, F.P.; Buffa, G.; Fantinato, E.; Innangi, M.; Stanisci, A. Identifying Critical Thresholds in the Impacts of Invasive Alien Plants and Dune Paths on Native Coastal Dune Vegetation. Land 2022, 12, 135. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P. Elton, C.S. 1958: The ecology of invasions by animals and plants. London: Methuen. Prog. Phys. Geogr. 2007, 31, 659–666. [Google Scholar] [CrossRef]
- Moyle, P.B.; Ellssworth, S. Alien Invaders, Essays on Wildlife Conservation. 2004. Available online: http://marinebio.org/Oceans/Conservation/Moyle (accessed on 1 January 2023).
- Izco Sevillano, J. Botánica; McGraw-Hill Interamericana de España S.L.: Madrid, Spain, 1997. [Google Scholar]
- Pippard, H.; Ralph, G.M.; Harvey, M.S.; Carpenter, K.E.; Buchanan, J.R.; Greenfield, D.W.; Harwell, H.D.; Larson, H.K.; Lawrence, A.; Linardich, C.; et al. The Conservation Status of Marine Biodiversity of the Pacific Islands of Oceania; IUCN: Gland, Switzerland, 2017; Volume viii, 59p. [Google Scholar] [CrossRef][Green Version]
- Mittenmeier, R.A.; Robles Gil, P.; Hoffman, M.; Pilgrim, J.; Brooks, T.; Goettsch Mittenmeier, C.; Lamoreux, J.; Da Fonseca, G. Hotspots Revisited: Earth’s Biologically Richest and Most Threatened Terrestrial Ecoregions; Conservation International: Ciudad de México, México; CEMEX: Mexico City, México, 2004. [Google Scholar]
- Aguilera Marín, N. Impactos de las Invasiones de Plantas en las Islas Oceánicas: El Caso de Dichrostachys cinerea (L.) Wight & Arn. 2010. Available online: https://www.researchgate.net/publication/284664079_Impactos_de_las_invasiones_de_plantas_en_las_islas_oceanicas_El_caso_de_Dichrostachys_cinerea_L_Wight_Arn/ (accessed on 1 January 2023).
- Sánchez-Hervás, J.M.; Ortz, I.; Maroño, M.; Ciria, P.; Ramos, R.; Arribas, L.; Domínguez, J. Gasificación de Biomasa e Hibridación AECID 2015/ACDE/001558. In Cogeneración de Energía, Eléctrica y Térmica, Mediante un Sistema Híbrido Biomasa-Solar para Explotaciones Agropecuarias en la Isla de Cuba; Informe Proyecto HYBRIDUS; Ciemat: Madrid, España, 2018. [Google Scholar]
- Méndez, I.; Moya, C.; Roquero, L. Primeras evidencias científicas de la presencia del marabú (Dichrostachys cinerea) en Cuba. An. La Acad. Cienc. Cuba 2022, 12. Available online: http://scielo.sld.cu/scielo.php?script=sci_arttext&pid=S2304-01062022000300012&lng=es&tlng=es (accessed on 1 January 2023).
- Prieto, R.; Oliver, P.; Caluff, M.; Regalado, L.; Ventosa, I.; Plasencia Fraga, J.; Baró, I.; González Gutiérrez, P.; Pérez-Camacho, J.; González-Oliva, L. Lista nacional de especies de plantas invasoras y potencialmente invasoras en la República de Cuba-2012. Bissea 2012, 6, 22–112. [Google Scholar]
- Nielsen, M.O.; Reinoso-Pérez, M.; Sørensen, M.; Hansen, H.; Gustafsson, J. Eco-Friendly Alternatives for Control and Use of Invasive Plants in Agroforestry Systems: The Case of Marabú (Dichrostachys cinerea) in Cuba. 2013. Available online: http://journal.um-surabaya.ac.id/index.php/JKM/article/view/2203 (accessed on 1 January 2023).
- Martín-Casas, N.; Reinoso-Pérez, M.; García-Díaz, J.R.; Hansen, H.H.; Nielsen, M.O. Evaluation of the feeding value of Dichrostachys cinerea pods for fattening pigs in Cuba. Trop. Anim. Health Prod. 2017, 49, 1235–1242. [Google Scholar] [CrossRef] [PubMed]
- Funes Monzote, R. El fin de los bosques y la plaga del marabú en Cuba. Historia de una “venganza ecológica”. Anu. Ecol. Cult. Soc. 2001, 1, 71–89. [Google Scholar]
- Ruiz Sinoga, J.D.; Remond Noa, R.; Fernández Perez, D. An Analysis of the Spatial Colonization of Scrubland Intrusive Species in the Itabo and Guanabo Watershed, Cuba. Remote Sens. 2010, 2, 740–757. [Google Scholar] [CrossRef]
- Jiménez Escudero, V.M. Desarrollo de Metodología de Teledetección para la Distribución Espacial de la Plaga Marabú (Dichrostachys cinerea) en Trinidad-Valle de los Ingenios (Patrimonio Cultural de la Humanidad UNESCO), Cuba. Master’s Thesis, Universidad Internacional de Andalucía, Seville, España, 2016. [Google Scholar]
- Grice, A.C.; Clarkson, J.R.; Calvert, M. Geographic Differentiation of Management Objectives for Invasive Species: A Case Study of Hymenachne Amplexicaulis in Australia. Environ. Sci. Policy 2011, 14, 986–997. [Google Scholar] [CrossRef]
- Mbaabu, P.R.; Ng, W.-T.; Schaffner, U.; Gichaba, M.; Olago, D.; Choge, S.; Oriaso, S.; Eckert, S. Spatial Evolution of Prosopis Invasion and its Effects on LULC and Livelihoods in Baringo, Kenya. Remote Sens. 2019, 11, 1217. [Google Scholar] [CrossRef]
- Bradley, B.A. Remote Detection of Invasive Plants: A Review of Spectral, Textural and Phenological Approaches. Biol. Invasions 2014, 16, 1411–1425. [Google Scholar] [CrossRef]
- Moreno, E.; Zabalo, A.; Gonzalez, E.; Alvarez, R.; Jimenez, V.M.; Menendez, J. Affordable Use of Satellite Imagery in Agriculture and Development Projects: Assessing the Spatial Distribution of Invasive Weeds in the UNESCO-Protected Areas of Cuba. Agriculture 2021, 11, 1057. [Google Scholar] [CrossRef]
- Betbeder, J.; Dubiez, E.; Gond, V.; Peltier, R. Rapport de Mission dans le Cadre de L’étude de Faisabilité Portant sur le Projet de Lutte contre la Prolifération de la Plante Invasive Marabú à Cuba; Centre de Coopération International en Recherche Agronomique pour le Développment: Montpellier, France, 2018. [Google Scholar]
- Almeida, E.; Dorta, D.; Alcantára, A. Metodología para estimación de área cubierta por D. cinerea a partir de imágenes satelitales. Univ. Cienc. 2010, 10, 32–44. [Google Scholar]
- Gaitán Rojas, D.J.; López Calle, M.I. Análisis Multitemporal de la Especie Vegetal Invasora Retamo Espinoso (Ulex europaeus) en el Embalse la Regadera, Zona Rural de la Localidad de Usme, a Partir de Imágenes Satelitales Sentinel 2 y Landsat 8 Mediante el Uso de Algoritmos de Clasificación; Universidad Distrital Francisco José de Caldas: Bogotá, Colombia, 2018. [Google Scholar]
- Jones, D.; Pike, S.; Thomas, M.; Murphy, D. Object- based image analysis for detection of Japanese Knotweed s.l. taxa (polygonaceae) in Wales (UK). Remote Sens. 2011, 3, 319–342. [Google Scholar] [CrossRef]
- Liu, M.; Li, H.; Li, L.; Man, W.; Jia, M.; Wang, Z.; Lu, C. Monitoring the invasion of Spartina alterniflora using multi-source high-resolution imagery in the Zhangjiang Estuary, China. Remote Sens. 2017, 9, 539. [Google Scholar] [CrossRef]
- Jensen, T.; Seerup Hass, F.; Seam Akbar, M.; Holm Petersen, P.; Jokar Arsanjani, J. Employing machine learning for detection of invasive species using sentinel-2 and Aviris data: The case of Kudzu in the United States. Sustainability 2020, 12, 3544. [Google Scholar] [CrossRef]
- Paz-Kagan, T.; Silver, M.; Panov, N.; Karnieli, A. Multispectral approach for identifying invasive plant species based on flowering phenology characteristics. Remote Sens. 2019, 11, 953. [Google Scholar] [CrossRef]
- Papp, L.; Van Leeuwen, B.; Szilassi, P.; Tobak, Z.; Szatmári, J.; Árvai, M.; Pásztor, L. Monitoring invasive plant species using hyperspectral remote sensing data. Land 2021, 10, 29. [Google Scholar] [CrossRef]
- Royimani, L.; Mutanga, O.; Odindi, J.; Dube, T.; Nyasha Matongera, T. Advancements in satellite remote sensing for mapping and monitoring of alien invasive plant species (AIPs). Phys. Chem. Earth Parts A/B/C 2019, 112, 237–245. [Google Scholar] [CrossRef]
- Matongera, T.N.; Mutanga, O.; Dube, T.; Sibanda, M. Detection and mapping the spatial distribution of bracken fern weeds using the Landsat 8 OLI new generation sensor. Int. J. Appl. Earth Obs. Geoinf. 2017, 57, 93–103. [Google Scholar] [CrossRef]
- Viana, H.; Aranha, J. Mapping invasive species (Acacia dealbata link) using ASTER/TERRA and LANDSAT 7 ETM+ imagery. In Forest Landscapes and Global Change-New Frontiers in Management, Conservation and Restoration Year, Proceedings of the IUFRO Landscape Ecology Working Group International Conference, Bragança, Portugal, 21–27 September 2010; IPB; IUFRO: Braganza, Portugal, 2010. [Google Scholar]
- Civco, D.L. Artificial neural networks for land-cover classification and mapping. Int. J. Geogr. Inf. Sci. 1993, 7, 173–186. [Google Scholar] [CrossRef]
- Geiß, C.; Aravena Pelizari, P.; Blickensdörfer, L.; Taubenböck, H. Virtual Support Vector Machines with Self-Learning Strategy for Classification of Multispectral Remote Sensing Imagery. ISPRS J. Photogramm. Remote Sens. 2019, 151, 42–58. [Google Scholar] [CrossRef]
- Carranza-García, M.; García-Gutiérrez, J.; Riquelme, J.C. A framework for evaluating land use and land cover classification using convolutional neural networks. Remote Sens. 2019, 11, 274. [Google Scholar] [CrossRef]
- Du, Q.; Chang, C.I. A linear constrained distance-based discriminant analysis for hyperspectral image classification. Pattern Recognit. 2001, 34, 361–373. [Google Scholar] [CrossRef]
- Kal-Yi, H. A synergistic automatic clustering technique (SYNERACT) for multispectral image Analysis. Photogramm. Eng. Remote Sens. 2002, 68, 33–40. [Google Scholar]
- Etter, A.; McAlpine, C.; Wilson, K.; Phinn, S.; Possingham, H. Regional patterns of agricultural land use and deforestation in Colombia. Agric. Ecosyst. Environ. 2006, 114, 369–386. [Google Scholar] [CrossRef]
- Boukabara, S.; Krasnopolsky, V.; Stewart, J.Q.; Maddy, E.S.; Shahroudi, N.; Hoffman, R.N. Leveraging Modern Artificial Intelligence for Remote Sensing and NWP: Benefits and Challenges. Bull. Am. Meteorol. Soc. 2019, 100, 473–491. [Google Scholar] [CrossRef]
- Geiß, C.; Pelizari, P.A.; Tunçbilek, O.; Taubenböck, H. Semi-supervised learning with constrained virtual support vector machines for classification of remote sensing image data. Int. J. Appl. Earth Obs. Geoinf. 2023, 125, 103571. [Google Scholar] [CrossRef]
- Ahmed, N.; Atzberger, C.; Zewdie, W. Integration of remote sensing and bioclimatic data for prediction of invasive species distribution in data-poor regions: A review on challenges and opportunities. Env. Syst. Res. 2020, 9, 32. [Google Scholar] [CrossRef]
- Kumar, M.; Padalia, H.; Singh, H. Remote sensing for mapping invasive alien plants: Opportunities and challenges. In A Handbook on Invasive Species, 1st ed.; Devi, K., Chaudhary, S.V., Kalia, S., Mishra, S.R., Eds.; Indian Council of Forestry Research and Education: Dehradun, India, 2020; Volume 1, pp. 16–31. [Google Scholar]
- Arasumani, M.; Bunyan, M.; Robin, V.V. Opportunities and challenges in using remote sensing for invasive tree species management, and in the identification of restoration sites in tropical montane grasslands. J. Environ. Manag. 2021, 280, 111759. [Google Scholar] [CrossRef] [PubMed]
- Shiferaw, H.; Bewket, W.; Eckert, S. Performances of machine learning algorithms for mapping fractional cover of an invasive plant species in a dryland ecosystem. Ecol. Evol. 2019, 9, 2562–2574. [Google Scholar] [CrossRef] [PubMed]
- Ouma, Y.O.; Gabasiane, T.G.; Nkhwanana, N. Mapping Prosopis L. (Mesquites) Using Sentinel-2 MSI Satellite Data, NDVI and SVI Spectral Indices with Maximum-Likelihood and Random Forest Classifiers. J. Sens. 2023, 2023, 18. [Google Scholar] [CrossRef]
- Huang, C.Y.; Asner, G.P. Applications of remote sensing to alien invasive plant studies. Sensors 2009, 9, 4869–4889. [Google Scholar] [CrossRef] [PubMed]
- Sims, D.A.; Gamon, J.A. Estimation of vegetation water content and photosynthetic tissue area from spectral reflectance: A comparison of indices based on liquid water and chlorophyll absorption features. Remote Sens. Environ. 2003, 84, 526–537. [Google Scholar] [CrossRef]
- Cervantes, J.; Garcia-Lamont, F.; Rodríguez-Mazahua, L.; Lopez, A. A comprehensive survey on support vector machine classification: Applications, challenges and trends. Neurocomputing 2020, 408, 189–215. [Google Scholar] [CrossRef]
- Vapnik, V. The Nature of Statistical Learning Theory, 2nd ed.; Michael, J., Lawless, J., Lauritzen, S., Nair, V., Eds.; Springer: Berlin/Heidelberg, Germany, 2000. [Google Scholar]
- Shang, X.; Chisholm, L. Classification of Australian Native Forest Species Using Hyperspectral Remote Sensing and Machine-Learning Classification Algorithms. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2014, 7, 2481–2489. [Google Scholar] [CrossRef]
- Cutler, D.R.; Edwards, T.C.; Beard, K.H.; Cutler, A.; Hess, K.T.; Gibson, J.; Lawler, J.J. Random Forests for Classification in Ecology. Ecology 2007, 88, 2783–2792. [Google Scholar] [CrossRef]
- Hernández-Blanco, Y.; Fernández-Rigondeaux, Y. Estudio de la evolución del sistema de asentamientos humanos de la provincia de Ciego de Ávila en el período 1981-2012. Noved. Poblac. 2019, 29, 192–202. [Google Scholar]
- Oficina Nacional de Estadística e Información República de Cuba (ONEI). Censo de Población y Viviendas 2012. Cuba. 2012. Available online: http://www.onei.gob.cu/node/13001 (accessed on 16 February 2023).
- Kulkarni, K.; Vijaya, P.A. Separability analysis of the band combinations for land cover classification of satellite images. Int. J. Eng. Trends Technol. 2021, 69, 138–144. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Sheykhmousa, M.; Mahdianpari, M.; Ghanbari, H.; Mohammadimanesh, F.; Ghamisi, P.; Homayouni, S. Support Vector Machine Versus Random Forest for Remote Sensing Image Classification: A Meta-Analysis and Systematic Review. IEEE J. Sel. Top. Appl. Earth Obs. Remote Sens. 2020, 13, 6308–6325. [Google Scholar] [CrossRef]
- Liaw, A.; Weiner, M. randomForest: Breiman and Cutler’s Random Forests for Classification and Regression; cran.r-project, R Package Version 4.6-7; R Package: Vienna, Austria, 2012. [Google Scholar]
- Congalton, R.G.; Green, K. Assessing the Accuracy of Remotely Sensed Data, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2009. [Google Scholar]
- Olofsson, P.; Foody, G.M.; Stehman, S.V.; Woodcock, C.E. Making better use of accuracy data in land change studies: Estimating accuracy and area and quantifying uncertainty using stratified estimation. Remote Sens. Environ. 2013, 129, 122–131. [Google Scholar] [CrossRef]
- Anaya, J.A.; Rodríguez-Buriticá, S.; Londoño, M.C. Clasificación de cobertura vegetal con resolución espacial de 10 metros en bosques del Caribe colombiano basado en misiones Sentinel 1 y 2. Rev. Teledetec. 2023, 61, 29–41. [Google Scholar] [CrossRef]
- Congalton, R.G. A review of assessing the accuracy of classifications of remotely sensed data. Remote Sens. Environ. 1991, 37, 35–46. [Google Scholar] [CrossRef]
- Sauro, J.; Lewis, J.R. Estimating completion rates from small samples using binomial confidence intervals: Comparisons and recommendations. In Proceedings of the Human Factors and Ergonomics Society Annual Meeting, Orlando, FL, USA, 26–30 September 2005; SAGE Publications: Thousand Oaks, CA, USA; Sage: Los Angeles, CA, USA, 2005. [Google Scholar]
- He, H.; Garcia, E.A. Learning from Imbalanced Data. IEEE Trans. Knowl. Data Eng. 2009, 21, 1263–1284. [Google Scholar]
- Fawcett, T. An introduction to ROC analysis. Pattern Recog. Lett. 2006, 27, 861–874. [Google Scholar] [CrossRef]
- Bradley, A.P. The use of the area under the ROC curve in the evaluation of machine learning algorithms. Pattern Recognit. 1997, 30, 1145–1159. [Google Scholar] [CrossRef]
- Demšar, J. Statistical comparisons of classifiers over multiple data sets. J. Mach. Learn. Res. 2006, 7, 1–30. [Google Scholar]
- Foody, G.M. Classification accuracy comparison: Hypothesis tests and the use of confidence intervals in evaluations of difference, equivalence and non-inferiority. Remote Sens. Environ. 2009, 113, 1658–1663. [Google Scholar] [CrossRef]
- Tonbul, H.; Colkesen, I.; Kavzoglu, T. Classification of poplar trees with object-based ensemble learning algorithms using Sentinel-2A imagery. J. Geod. Sci. 2020, 10, 14–22. [Google Scholar] [CrossRef]
- Ndlovu, H.S.; Sibanda, M.; Odindi, J.; Buthelezi, S.; Mutanga, O. Detecting and mapping the spatial distribution of Chromoleana odorata invasions in communal areas of South Africa using Sentinel-2 multispectral remotely sensed data. Phys. Chem. Earth Parts A/B/C 2022, 126, 103081. [Google Scholar] [CrossRef]
- Pouteau, R.; Meyer, J.Y.; Taputuarai, R.; Stoll, B. Support vector machines to map rare and endangered native plants in Pacific islands forests. Ecol. Inform. 2012, 9, 37–46. [Google Scholar] [CrossRef]
- Linhui, L.; Weipeng, J.; Huihui, W. Extracting the forest type from remote sensing images by random forest. IEEE Sens. J. 2020, 21, 17447–17454. [Google Scholar] [CrossRef]
- Jombo, S.; Adam, E. Comparison between Maximum likelihood and Support Vector Machines classifiers in mapping urban tree species using spot 7 imagery. In Geography and Community Research, Learning, Impact, Proceedings of the Biennial Conference of the Society of South African Geographers; University of the Free State: Bloemfontein, South Africa, 2018; Volume 1, p. 684. [Google Scholar]
- Wulder, M.A.; Hermosilla, T.; White, J.C.; Hobart, G.; Masek, J.G. Augmenting Landsat time series with Harmonized Landsat Sentinel-2 data products: Assessment of spectral correspondence. Sci. Remote Sens. 2021, 4, 100031. [Google Scholar] [CrossRef]
- Galford, G.L.; Fernandez, M.; Roman, J.; Monasterolo, I.; Ahamed, S.; Fiske, G.; González Díaz, P.; Kaufman, L. Cuban land use and conservation, from rainforests to coral reefs. Bull. Mar. Sci. 2018, 94, 171–191. [Google Scholar] [CrossRef]
- Valero-Jorge, A.; González-De Zayas, R.; Alcántara-Martín, A.; Álvarez-Taboada, F.; Matos-Pupo, F.; Brown-Manrique, O. Water area and volume calculation of two reservoirs in Central Cuba using remote sensing methods. A new perspective. Rev. Teledetec. 2022, 60, 71–87. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| LULC | Description | Training Areas (Polygons/Pixels) | |
|---|---|---|---|
| 1994 | 2022 | ||
| D. cinerea (marabú) | Refers to wooded and scrub areas of D. cinerea | 44/308 | 62/310 |
| Grassland | Includes areas of natural herbaceous vegetation or grass cover | 63/504 | 54/432 |
| Irrigated crops | Includes herbaceous and woody crops under irrigated conditions | 161/966 | 71/726 |
| Rainfed crops | Includes dryland herbaceous crops that depend on rainfall for water | 69/552 | 44/564 |
| Woodland | Natural areas of tree vegetation, such as forests of any species | 64/743 | 82/738 |
| Mangrove | Includes coastal areas of mangroves as the main vegetation | 166/1358 | 147/1323 |
| Flood-prone areas | Includes flooded crops and areas near the coast with a high potential for flooding | 152/1912 | 128/1152 |
| Bare soil | Includes areas devoid of vegetation, dirt roads, fallow lands, mining deposits, etc. (uncovered soils) | 130/912 | 81/648 |
| Infrastructure | Includes built-up areas, generally with urban or industrial use, and paved roads | 86/516 | 94/658 |
| Water | Bodies of water such as rivers, ponds, or reservoirs. | 64/384 | 68/544 |
| D. cinerea | All Classes | |||||
|---|---|---|---|---|---|---|
| Year | Algorithm | PA (%) | UA (%) | AUC | F-Score | OA (%) |
| 1994 | RF | 83.64 (71.51–91.37) | 93.88 (82.85–98.52) | 0.92 | 0.88 | 90.91 (88.20–93.05) |
| ML | 58.18 (45.02–70.27) | 86.49 (71.55–94.56) | 0.72 | 0.69 | 78.18 (74.54–81.44) | |
| SVM | 56.36 (43.26–68.63) | 83.78 (68.48–92.73) | 0.69 | 0.67 | 74.73 (70.93–78.18) | |
| 2022 | RF | 98.18 (89.49–99.99) | 96.43 (87.18–99.72) | 0.97 | 0.97 | 95.09 (92.93–96.63) |
| ML | 80.00 (67.46–88.62) | 74.58 (62.11–84.04) | 0.87 | 0.77 | 77.63 (73.96–80.93) | |
| SVM | 80.00 (67.46–88.62) | 64.71 (52.81–75.02) | 0.72 | 0.71 | 71.27 (67.35–74.90) | |
| Year | Pairwise Comparison | McNemar´s |
|---|---|---|
| 1994 | RF vs. ML | 28.82 |
| RF vs. SVM | 41.91 | |
| ML vs. SVM | 1.39 | |
| 2022 | RF vs. ML | 61.44 |
| RF vs. SVM | 92.76 | |
| ML vs. SVM | 4.35 |
| LULC Classes | 1994 | 2022 | ||
|---|---|---|---|---|
| Ha | % of Total Area | Ha | % of Total Area | |
| Water | 12,799.42 | 2.04 | 12,898.86 | 2.05 |
| Woodland | 44,241.82 | 7.07 | 30,613.68 | 4.86 |
| Infrastructure | 11,410.92 | 1.82 | 31,160.16 | 4.95 |
| Grassland | 163,501.12 | 26.14 | 125,860.58 | 20.01 |
| Irrigated crops | 19,320.3 | 3.08 | 17,264.19 | 2.74 |
| Bare soil | 16,622.47 | 2.65 | 73,823.71 | 12.33 |
| Flood-prone areas | 95,312.79 | 15.24 | 99,268.14 | 15.78 |
| D. cinerea | 61,977.59 | 9.91 | 91,533.47 | 14.56 |
| Mangrove | 25,871.49 | 4.14 | 19,229.40 | 3.06 |
| Rainfed crops | 174,248.46 | 27.86 | 123,366.68 | 19.62 |
| Municipality | Area 1994 (ha) | Area 2022 (ha) | Net Change (ha) | % of Class Area in 1994 |
|---|---|---|---|---|
| Primero de Enero | 10,229.08 | 13,061.79 | 2832.71 | 27.69 |
| Majagua | 3180.96 | 6737.71 | 3556.75 | 111.81 |
| Ciro Redondo | 9869.36 | 7898.66 | −1970.70 | −19.96 |
| Florencia | 6238.19 | 7387.68 | 1149.00 | 18.41 |
| Ciego de Ávila | 6468.68 | 6364.71 | −103.97 | −1.61 |
| Bolivia | 8831.28 | 18,729.04 | 9897.76 | 112.07 |
| Morón | 3257.55 | 5834.08 | 2576.53 | 79.09 |
| Chambas | 7396.34 | 8649.63 | 1253.29 | 38.47 |
| Baraguá | 4563.04 | 5994.61 | 1431.57 | 31.37 |
| Venezuela | 1943.11 | 10,875.56 | 8932.45 | 459.69 |
| Net Changes 1994–2022 | |||
|---|---|---|---|
| LULC | ha | % of Total Area | % of Change per Class by Area from 1994 to 2022 |
| Water | 99.44 | 0.01 | 0.77 |
| Woodland | −13,268.14 | −2.21 | −29.99 |
| Infrastructure | 19,749.24 | 3.13 | 173.07 |
| Grassland | −37,640.54 | −6.13 | −23.02 |
| Irrigated crops | −2056.11 | −0.34 | −10.64 |
| Bare soil | 60,929.24 | 9.68 | 366.54 |
| Flood-prone areas | 3955.35 | 0.54 | 4.14 |
| D. cinerea | 29,555.88 | 4.65 | 47.68 |
| Mangrove | −6642.09 | −1.08 | −25.67 |
| Rainfed crops | −50,881.78 | −8.24 | −29.20 |
| LULC | Losses to D. cinerea (ha) | Gains from D. cinerea (ha) | Net Change (ha) | % of Change for the Total Area from 1994 to 2022 |
|---|---|---|---|---|
| Water | 24.30 | 101.07 | 76.77 | 0.59 |
| Woodland | 24,691.05 | 7900.38 | −16,790.67 | −37.95 |
| Infrastructure | 288.27 | 1860.21 | 1571.94 | 13.77 |
| Grassland | 18,838.80 | 14,652.36 | −4186.44 | −2.56 |
| Irrigated crops | 1388.43 | 1196.01 | −192.42 | −1.00 |
| Bare soil | 1212.84 | 5166.99 | 3954.15 | 23.78 |
| Flood-prone areas | 6477.03 | 4392.27 | −2084.76 | −2.19 |
| Mangrove | 1135.26 | 55.53 | −1079.73 | −4.17 |
| Rainfed crops | 20,031.39 | 13,260.15 | −6771.24 | −3.88 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Valero-Jorge, A.; González-De Zayas, R.; Matos-Pupo, F.; Becerra-González, A.L.; Álvarez-Taboada, F. Mapping and Monitoring of the Invasive Species Dichrostachys cinerea (Marabú) in Central Cuba Using Landsat Imagery and Machine Learning (1994–2022). Remote Sens. 2024, 16, 798. https://doi.org/10.3390/rs16050798
Valero-Jorge A, González-De Zayas R, Matos-Pupo F, Becerra-González AL, Álvarez-Taboada F. Mapping and Monitoring of the Invasive Species Dichrostachys cinerea (Marabú) in Central Cuba Using Landsat Imagery and Machine Learning (1994–2022). Remote Sensing. 2024; 16(5):798. https://doi.org/10.3390/rs16050798
Chicago/Turabian StyleValero-Jorge, Alexey, Roberto González-De Zayas, Felipe Matos-Pupo, Angel Luis Becerra-González, and Flor Álvarez-Taboada. 2024. "Mapping and Monitoring of the Invasive Species Dichrostachys cinerea (Marabú) in Central Cuba Using Landsat Imagery and Machine Learning (1994–2022)" Remote Sensing 16, no. 5: 798. https://doi.org/10.3390/rs16050798
APA StyleValero-Jorge, A., González-De Zayas, R., Matos-Pupo, F., Becerra-González, A. L., & Álvarez-Taboada, F. (2024). Mapping and Monitoring of the Invasive Species Dichrostachys cinerea (Marabú) in Central Cuba Using Landsat Imagery and Machine Learning (1994–2022). Remote Sensing, 16(5), 798. https://doi.org/10.3390/rs16050798