
Matching Vegetation Indices and Tree Vigor in Pyrenean Silver Fir Stands

 ,
,  ,
,  , ,
, ,  ,
,  and
and 
Abstract
1. Introduction
2. Materials and Methods
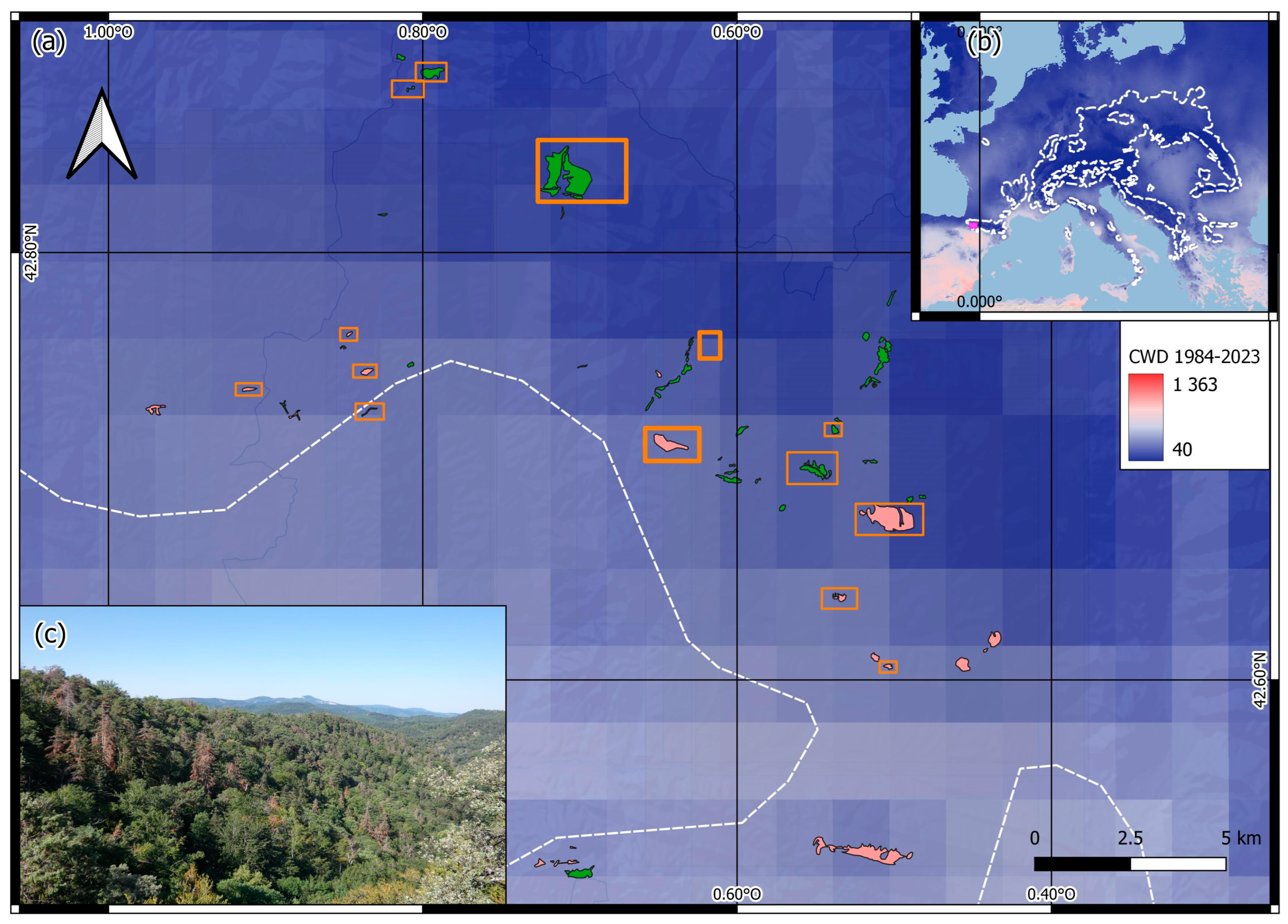
2.1. Study Sites and Field Sampling
2.2. Climatological Data
2.3. Remote Sensing Data and Vegetation Indices
2.4. Statistical Analyses
3. Results
3.1. Characteristics of Declining and Non-Declining Silver Fir Stands
3.2. Climate Trends and Drought Variability
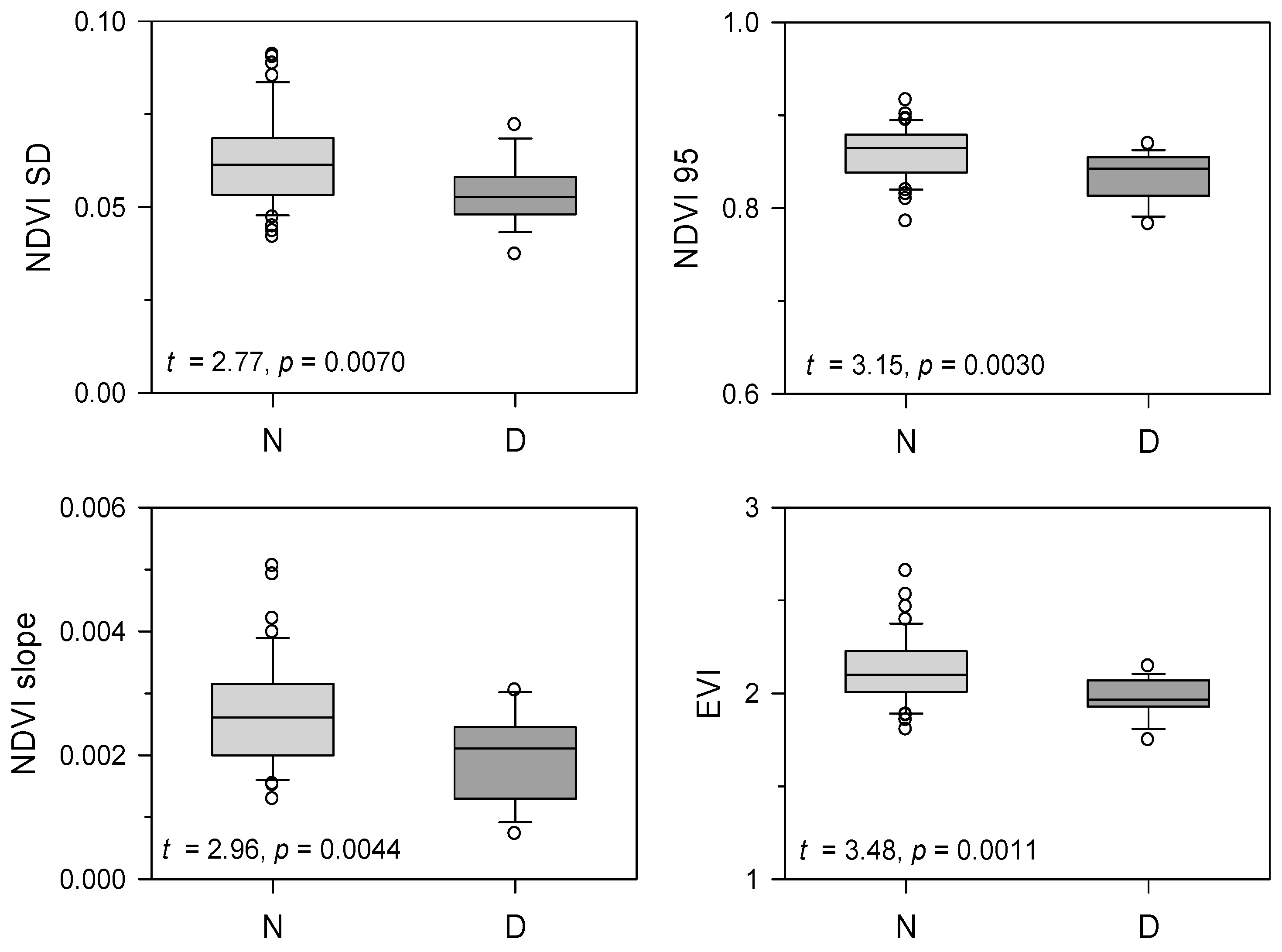
3.3. Comparing Vegetation Indices Between Declining and Non-Declining Stands
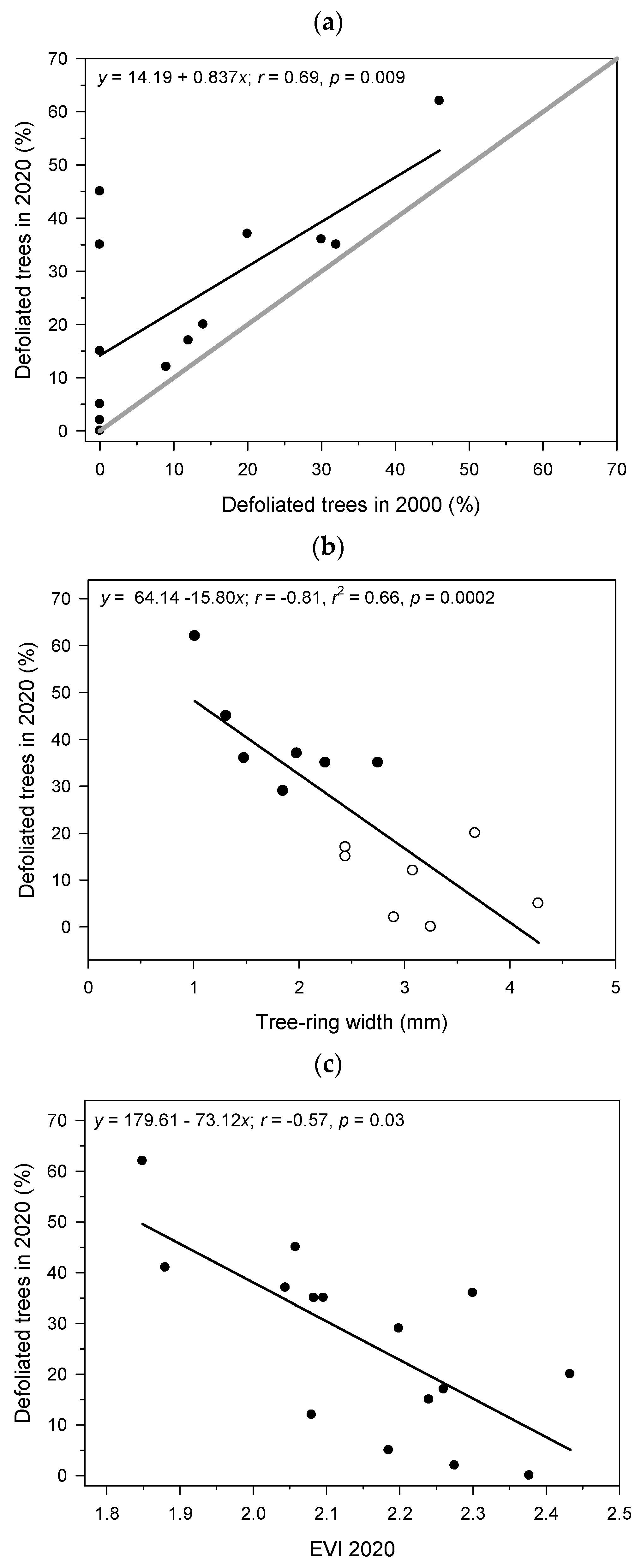
3.4. Relationships Between EVI and Stand Defoliation
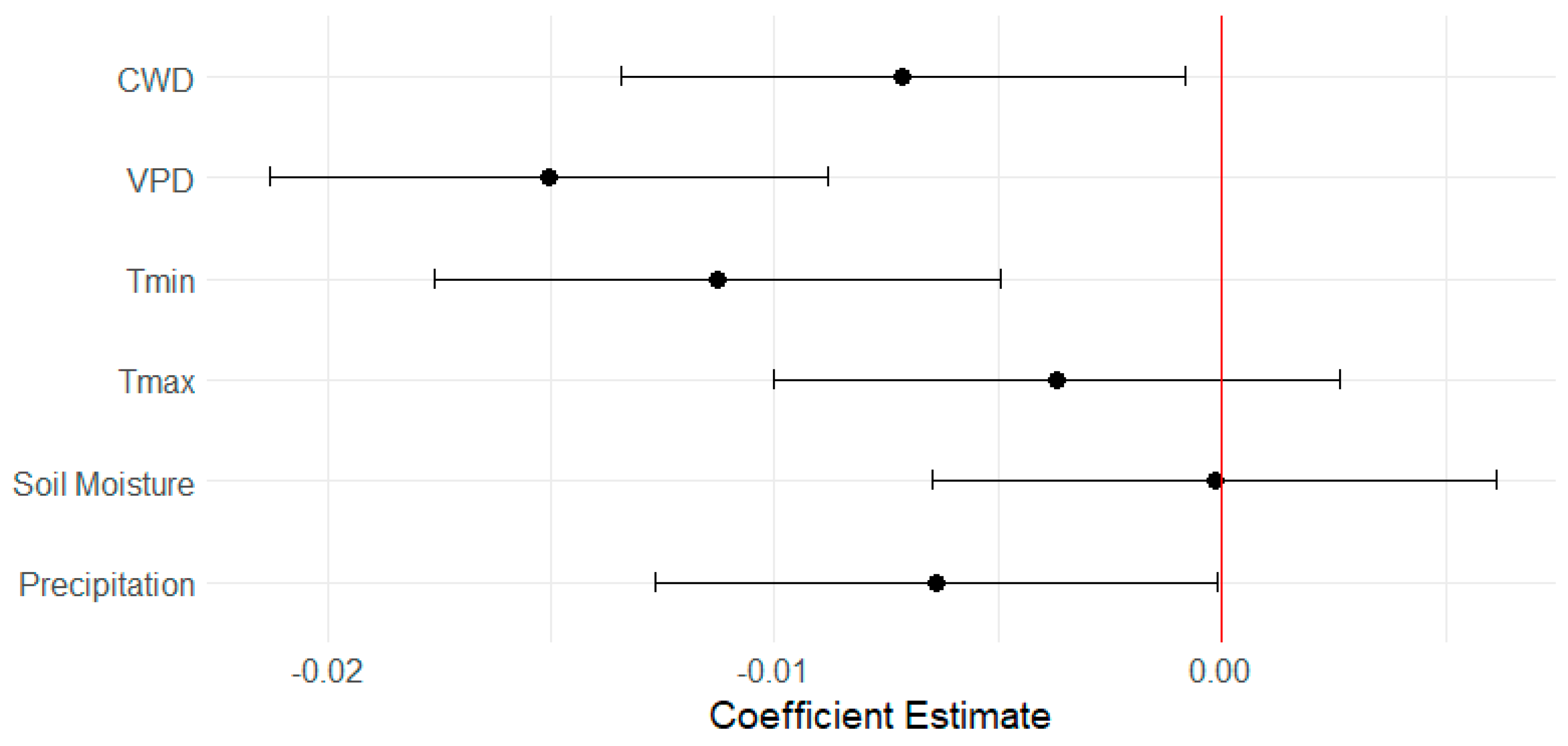
3.5. Responses of EVI to Climate Variables
4. Discussion
4.1. Comparison of NDVI and EVI Between Stands of Different Vigor
4.2. Relationship of NDVI and EVI with Vigor Field Data
4.3. Climatic Factors Influencing Vegetation Indices
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Buras, A.; Rammig, A.; Zang, C.S. The European Forest Condition Monitor: Using Remotely Sensed Forest Greenness to Identify Hot Spots of Forest Decline. Front. Plant Sci. 2021, 12, 689220. [Google Scholar] [CrossRef] [PubMed]
- Gazol, A.; Camarero, J.J. Compound Climate Events Increase Tree Drought Mortality across European Forests. Sci. Total Environ. 2022, 816, 151604. [Google Scholar] [CrossRef] [PubMed]
- Senf, C.; Buras, A.; Zang, C.S.; Rammig, A.; Seidl, R. Excess Forest Mortality Is Consistently Linked to Drought across Europe. Nat. Commun. 2020, 11, 6200. [Google Scholar] [CrossRef]
- Hartmann, H.; Moura, C.F.; Anderegg, W.R.L.; Ruehr, N.K.; Salmon, Y.; Allen, C.D.; Arndt, S.K.; Breshears, D.D.; Davi, H.; Galbraith, D.; et al. Research Frontiers for Improving Our Understanding of Drought-induced Tree and Forest Mortality. New Phytol. 2018, 218, 15–28. [Google Scholar] [CrossRef]
- Thonfeld, F.; Gessner, U.; Holzwarth, S.; Kriese, J.; Da Ponte, E.; Huth, J.; Kuenzer, C. A First Assessment of Canopy Cover Loss in Germany’s Forests after the 2018–2020 Drought Years. Remote Sens. 2022, 14, 562. [Google Scholar] [CrossRef]
- Alibakhshi, S. A Robust Approach and Analytical Tool for Identifying Early Warning Signals of Forest Mortality. Ecol. Indic. 2023, 155, 110983. [Google Scholar] [CrossRef]
- Yan, Y.; Piao, S.; Hammond, W.M.; Chen, A.; Hong, S.; Xu, H.; Munson, S.M.; Myneni, R.B.; Allen, C.D. Climate-Induced Tree-Mortality Pulses Are Obscured by Broad-Scale and Long-Term Greening. Nat. Ecol. Evol. 2024, 8, 912–923. [Google Scholar] [CrossRef]
- Hammond, W.M.; Johnson, D.M.; Meinzer, F.C. A Thin Line between Life and Death: Radial Sap Flux Failure Signals Trajectory to Tree Mortality. Plant Cell Environ. 2021, 44, 1311–1314. [Google Scholar] [CrossRef]
- Rogers, B.M.; Solvik, K.; Hogg, E.H.; Ju, J.; Masek, J.G.; Michaelian, M.; Berner, L.T.; Goetz, S.J. Detecting Early Warning Signals of Tree Mortality in Boreal North America Using Multiscale Satellite Data. Glob. Chang. Biol. 2018, 24, 2284–2304. [Google Scholar] [CrossRef]
- Camarero, J.J.; Bigler, C.; Linares, J.C.; Gil-Pelegrín, E. Synergistic Effects of Past Historical Logging and Drought on the Decline of Pyrenean Silver Fir Forests. For. Ecol. Manag. 2011, 262, 759–769. [Google Scholar] [CrossRef]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological Maps for the Main European Woody Species. Data Brief. 2017, 12, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Balao, F.; Rios, M.; Sanchez-Robles, J.M.; Lorenzo, M.T.; Linares, J.C.; Terrab, A. Postglacial Genetic Legacies, Demography, and Climate Change: Setting Conservation Priorities for Silver Fir. bioRxiv 2024. [Google Scholar] [CrossRef]
- Vitasse, Y.; Bottero, A.; Cailleret, M.; Bigler, C.; Fonti, P.; Gessler, A.; Lévesque, M.; Rohner, B.; Weber, P.; Rigling, A.; et al. Contrasting Resistance and Resilience to Extreme Drought and Late Spring Frost in Five Major European Tree Species. Glob. Chang. Biol. 2019, 25, 3781–3792. [Google Scholar] [CrossRef] [PubMed]
- Gazol, A.; Camarero, J.J.; Gutiérrez, E.; Popa, I.; Andreu-Hayles, L.; Motta, R.; Nola, P.; Ribas, M.; Sangüesa-Barreda, G.; Urbinati, C.; et al. Distinct Effects of Climate Warming on Populations of Silver Fir (Abies alba) across Europe. J. Biogeogr. 2015, 42, 1150–1162. [Google Scholar] [CrossRef]
- Camarero, J.J. The Multiple Factors Explaining Decline in Mountain Forests: Historical Logging and Warming-Related Drought Stress Is Causing Silver-Fir Dieback in the Aragón Pyrenees. In High Mountain Conservation in a Changing World; Catalan, J., Ninot, J.M., Aniz, M.M., Eds.; Advances in Global Change Research; Springer International Publishing: Cham, Switzerland, 2017; Volume 62, pp. 131–154. [Google Scholar] [CrossRef]
- Dobrowolska, D.; Bončina, A.; Klumpp, R. Ecology and Silviculture of Silver Fir (Abies alba Mill.): A Review. J. For. Res. 2017, 22, 326–335. [Google Scholar] [CrossRef]
- Gazol, A.; González De Andrés, E.; Colangelo, M.; Valeriano, C.; Camarero, J.J. Pyrenean Silver Fir Forests Retain Legacies of Past Disturbances and Climate Change in Their Growth, Structure and Composition. Forests 2023, 14, 713. [Google Scholar] [CrossRef]
- Sangüesa-Barreda, G.; Camarero, J.J.; Oliva, J.; Montes, F.; Gazol, A. Past Logging, Drought and Pathogens Interact and Contribute to Forest Dieback. Agric. For. Meteorol. 2015, 208, 85–94. [Google Scholar] [CrossRef]
- Valeriano, C.; Tumajer, J.; Gazol, A.; González De Andrés, E.; Sánchez-Salguero, R.; Colangelo, M.; Linares, J.C.; Valor, T.; Sangüesa-Barreda, G.; Julio Camarero, J. Delineating Vulnerability to Drought Using a Process-Based Growth Model in Pyrenean Silver Fir Forests. For. Ecol. Manag. 2023, 541, 121069. [Google Scholar] [CrossRef]
- Vejpustková, M.; Čihák, T.; Fišer, P. The Increasing Drought Sensitivity of Silver Fir (Abies alba Mill.) Is Evident in the Last Two Decades. J. For. Sci. 2023, 69, 67–79. [Google Scholar] [CrossRef]
- Wang, J.; Sammis, T.W.; Gutschick, V.P.; Gebremichael, M.; Dennis, S.O.; Harrison, R.E. Review of Satellite Remote Sensing Use in Forest Health Studies. TOGEOGJ 2010, 3, 28–42. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the Radiometric and Biophysical Performance of the MODIS Vegetation Indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Khoury, S.; Coomes, D.A. Resilience of Spanish Forests to Recent Droughts and Climate Change. Glob. Chang. Biol. 2020, 26, 7079–7098. [Google Scholar] [CrossRef] [PubMed]
- Rouse, J.W.; Haas, R.H.; Schell, J.A.; Deering, D.W. Monitoring Vegetation Systems in the Great Plains with ERTS. NASA Spec. Publ. 1974, 1, 309. [Google Scholar]
- Huete, A. A Comparison of Vegetation Indices over a Global Set of TM Images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Xiao, X.; Braswell, B.; Zhang, Q.; Boles, S.; Frolking, S.; Moore, B. Sensitivity of Vegetation Indices to Atmospheric Aerosols: Continental-Scale Observations in Northern Asia. Remote Sens. Environ. 2003, 84, 385–392. [Google Scholar] [CrossRef]
- Fiore, N.M.; Goulden, M.L.; Czimczik, C.I.; Pedron, S.A.; Tayo, M.A. Do Recent NDVI Trends Demonstrate Boreal Forest Decline in Alaska? Environ. Res. Lett. 2020, 15, 095007. [Google Scholar] [CrossRef]
- Parent, M.B.; Verbyla, D. The Browning of Alaska’s Boreal Forest. Remote Sens. 2010, 2, 2729–2747. [Google Scholar] [CrossRef]
- Aubard, V.; Paulo, J.A.; Silva, J.M.N. Long-Term Monitoring of Cork and Holm Oak Stands Productivity in Portugal with Landsat Imagery. Remote Sens. 2019, 11, 525. [Google Scholar] [CrossRef]
- Vicente Serrano, S.M.; Lasanta, T.; Romo, A. Diferencias Espaciales En La Evolución Del NDVI En La Cuenca Alta Del Aragón: Efectos de Los Caminos En El Uso Del Suelo. CIG 2003, 29, 51–66. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; Lasanta, T. Diversidad Espacial de La Actividad Vegetal En Campos Abandonados Del Pirineo Central Español: Análisis de Los Procesos de Sucesión Mediante Imágenes Landsat (1984-2001). Pirineos 2006, 161, 59–84. [Google Scholar] [CrossRef]
- Alatorre, L.C.; Beguería, S.; Vicente-Serrano, S.M. Análisis de La Evolución Espacio-Temporal Del NDVI Sobre Áreas Vegetadas y Zonas de Riesgo de Erosión En El Pirineo Central. Pirineos 2010, 165, 7–27. [Google Scholar] [CrossRef]
- Xiao, X.; Hollinger, D.; Aber, J.; Goltz, M.; Davidson, E.A.; Zhang, Q.; Moore, B. Satellite-Based Modeling of Gross Primary Production in an Evergreen Needleleaf Forest. Remote Sens. Environ. 2004, 89, 519–534. [Google Scholar] [CrossRef]
- Lausch, A.; Erasmi, S.; King, D.; Magdon, P.; Heurich, M. Understanding Forest Health with Remote Sensing -Part I—A Review of Spectral Traits, Processes and Remote-Sensing Characteristics. Remote Sens. 2016, 8, 1029. [Google Scholar] [CrossRef]
- Barka, I.; Bucha, T.; Molnár, T.; Móricz, N.; Somogyi, Z.; Koreň, M. Suitability of MODIS-Based NDVI Index for Forest Monitoring and Its Seasonal Applications in Central Europe. Cent. Eur. For. J. 2019, 65, 206–217. [Google Scholar] [CrossRef]
- Berner, L.T.; Beck, P.S.A.; Bunn, A.G.; Lloyd, A.H.; Goetz, S.J. High-Latitude Tree Growth and Satellite Vegetation Indices: Correlations and Trends in Russia and Canada (1982–2008). J. Geophys. Res. 2011, 116, G01015. [Google Scholar] [CrossRef]
- Bunn, A.G.; Hughes, M.K.; Kirdyanov, A.V.; Losleben, M.; Shishov, V.V.; Berner, L.T.; Oltchev, A.; Vaganov, E.A. Comparing Forest Measurements from Tree Rings and a Space-Based Index of Vegetation Activity in Siberia. Environ. Res. Lett. 2013, 8, 035034. [Google Scholar] [CrossRef]
- Gallardo, V.B.; Hadad, M.A.; Roig, F.A.; Gatica, G.; Chen, F. Spatio-Temporal Linkage Variations between NDVI and Tree Rings on the Leeward Side of the Northern Patagonian Andes. For. Ecol. Manag. 2024, 553, 121593. [Google Scholar] [CrossRef]
- Montoya-Jiménez, L.R.; Gómez-Guerrero, A.; Pedraza-Oropeza, F.J.A.; González-Martínez, T.M.; Correa-Díaz, A. Short-Term Tree-Ring Series of Pinus Hartwegii Lindl. Taken at Ground Level Correlate to Normalized Difference Vegetation Index Series. Forests 2024, 15, 324. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Camarero, J.J.; Olano, J.M.; Martín-Hernández, N.; Peña-Gallardo, M.; Tomás-Burguera, M.; Gazol, A.; Azorin-Molina, C.; Bhuyan, U.; El Kenawy, A. Diverse Relationships between Forest Growth and the Normalized Difference Vegetation Index at a Global Scale. Remote Sens. Environ. 2016, 187, 14–29. [Google Scholar] [CrossRef]
- Gazol, A.; Camarero, J.J.; Vicente-Serrano, S.M.; Sánchez-Salguero, R.; Gutiérrez, E.; De Luis, M.; Sangüesa-Barreda, G.; Novak, K.; Rozas, V.; Tíscar, P.A.; et al. Forest Resilience to Drought Varies across Biomes. Glob. Chang. Biol. 2018, 24, 2143–2158. [Google Scholar] [CrossRef]
- Wang, Z.; Lyu, L.; Liu, W.; Liang, H.; Huang, J.; Zhang, Q.-B. Topographic Patterns of Forest Decline as Detected from Tree Rings and NDVI. CATENA 2021, 198, 105011. [Google Scholar] [CrossRef]
- Zhang, T.; Song, J.; Fan, Y.; Liu, Y.; Yu, S.; Guo, D.; Hou, T.; Guo, K. Vegetation Index Research on the Basis of Tree-Ring Data: Current Status and Prospects. Forests 2023, 14, 2016. [Google Scholar] [CrossRef]
- Mašek, J.; Tumajer, J.; Lange, J.; Kaczka, R.; Fišer, P.; Treml, V. Variability in Tree-Ring Width and NDVI Responses to Climate at a Landscape Level. Ecosystems 2023, 26, 1144–1157. [Google Scholar] [CrossRef]
- Norman, S.P.; Koch, F.H.; Hargrove, W.W. Review of Broad-Scale Drought Monitoring of Forests: Toward an Integrated Data Mining Approach. For. Ecol. Manag. 2016, 380, 346–358. [Google Scholar] [CrossRef]
- Gazol, A.; Sangüesa-Barreda, G.; Camarero, J.J. Forecasting Forest Vulnerability to Drought in Pyrenean Silver Fir Forests Showing Dieback. Front. For. Glob. Chang. 2020, 3, 36. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Rings and Climate; Academic Press: London, UK; New York, NY, USA, 1976. [Google Scholar]
- Cottam, G.; Curtis, J.T. The Use of Distance Measures in Phytosociological Sampling. Ecology 1956, 37, 451–460. [Google Scholar] [CrossRef]
- Abatzoglou, J.T.; Dobrowski, S.Z.; Parks, S.A.; Hegewisch, K.C. TerraClimate, a High-Resolution Global Dataset of Monthly Climate and Climatic Water Balance from 1958–2015. Sci Data 2018, 5, 1–12. [Google Scholar] [CrossRef]
- Beguería, S.; Vicente-Serrano, S.M. SPEI: Calculation of the Standardized Precipitation-Evapotranspiration Index. 2023. Available online: https://CRAN.R-project.org/package=SPEI (accessed on 20 October 2024).
- Abu Arra, A.; Şişman, E. A Comprehensive Analysis and Comparison of SPI and SPEI for Spatiotemporal Drought Evaluation. Env. Monit. Assess. 2024, 196, 980. [Google Scholar] [CrossRef]
- Peña-Gallardo, M.; Vicente-Serrano, S.M.; Camarero, J.J.; Gazol, A.; Sánchez-Salguero, R.; Domínguez-Castro, F.; El Kenawy, A.; Beguería-Portugés, S.; Gutiérrez, E.; De Luis, M.; et al. Drought Sensitiveness on Forest Growth in Peninsular Spain and the Balearic Islands. Forests 2018, 9, 524. [Google Scholar] [CrossRef]
- Yuan, W.; Zheng, Y.; Piao, S.; Ciais, P.; Lombardozzi, D.; Wang, Y.; Ryu, Y.; Chen, G.; Dong, W.; Hu, Z.; et al. Increased Atmospheric Vapor Pressure Deficit Reduces Global Vegetation Growth. Sci. Adv. 2019, 5, eaax1396. [Google Scholar] [CrossRef]
- Liu, X.; He, B.; Guo, L.; Huang, L.; Yuan, W.; Chen, X.; Hao, X.; Xie, X.; Zhang, Y.; Zhong, Z.; et al. European Carbon Uptake Has Not Benefited from Vegetation Greening. Geophys. Res. Lett. 2021, 48, e2021GL094870. [Google Scholar] [CrossRef]
- Grossiord, C.; Buckley, T.N.; Cernusak, L.A.; Novick, K.A.; Poulter, B.; Siegwolf, R.T.W.; Sperry, J.S.; McDowell, N.G. Plant Responses to Rising Vapor Pressure Deficit. New Phytol. 2020, 226, 1550–1566. [Google Scholar] [CrossRef]
- Luo, Y.; Liang, W.; Yan, J.; Zhang, W.; Gou, F.; Wang, C.; Liang, X. Vegetation Growth Response and Trends after Water Deficit Exposure in the Loess Plateau, China. Remote Sens. 2023, 15, 2593. [Google Scholar] [CrossRef]
- Vicente-Serrano, S.M.; Beguería, S.; López-Moreno, J.I. A Multiscalar Drought Index Sensitive to Global Warming: The Standardized Precipitation Evapotranspiration Index. J. Clim. 2010, 23, 1696–1718. [Google Scholar] [CrossRef]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-Scale Geospatial Analysis for Everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- White, J.C.; Wulder, M.A.; Hobart, G.W.; Luther, J.E.; Hermosilla, T.; Griffiths, P.; Coops, N.C.; Hall, R.J.; Hostert, P.; Dyk, A.; et al. Pixel-Based Image Compositing for Large-Area Dense Time Series Applications and Science. Can. J. Remote Sens. 2014, 40, 192–212. [Google Scholar] [CrossRef]
- Zhu, Z.; Woodcock, C.E. Object-Based Cloud and Cloud Shadow Detection in Landsat Imagery. Remote Sens. Environ. 2012, 118, 83–94. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing. 2024. Available online: https://www.R-project.org/ (accessed on 20 September 2024).
- Fox, J.; Weisberg, S. An R Companion to Applied Regression. 2023. Available online: https://CRAN.R-project.org/package=car (accessed on 20 October 2024).
- Torres, P.; Rodes-Blanco, M.; Viana-Soto, A.; Nieto, H.; García, M. The Role of Remote Sensing for the Assessment and Monitoring of Forest Health: A Systematic Evidence Synthesis. Forests 2021, 12, 1134. [Google Scholar] [CrossRef]
- Loranty, M.M.; Davydov, S.P.; Kropp, H.; Alexander, H.D.; Mack, M.C.; Natali, S.M.; Zimov, N.S. Vegetation Indices Do Not Capture Forest Cover Variation in Upland Siberian Larch Forests. Remote Sens. 2018, 10, 1686. [Google Scholar] [CrossRef]
- Novillo, C.J.; Arrogante-Funes, P.; Romero-Calcerrada, R. Recent NDVI Trends in Mainland Spain: Land-Cover and Phytoclimatic-Type Implications. IJGI 2019, 8, 43. [Google Scholar] [CrossRef]
- Astigarraga, J.; Andivia, E.; Zavala, M.A.; Gazol, A.; Cruz-Alonso, V.; Vicente-Serrano, S.M.; Ruiz-Benito, P. Evidence of Non-stationary Relationships between Climate and Forest Responses: Increased Sensitivity to Climate Change in Iberian Forests. Glob. Chang. Biol. 2020, 26, 5063–5076. [Google Scholar] [CrossRef] [PubMed]
- Matsushita, B.; Yang, W.; Chen, J.; Onda, Y.; Qiu, G. Sensitivity of the Enhanced Vegetation Index (EVI) and Normalized Difference Vegetation Index (NDVI) to Topographic Effects: A Case Study in High-Density Cypress Forest. Sensors 2007, 7, 2636–2651. [Google Scholar] [CrossRef] [PubMed]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To Die or Not to Die: Early Warnings of Tree Dieback in Response to a Severe Drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef]
- Sánchez-Salguero, R.; Camarero, J.; Grau, J.; De La Cruz, A.; Gil, P.; Minaya, M.; Fernández-Cancio, Á. Analysing Atmospheric Processes and Climatic Drivers of Tree Defoliation to Determine Forest Vulnerability to Climate Warming. Forests 2016, 8, 13. [Google Scholar] [CrossRef]
- Wei, M.; Jiao, L.; Zhang, P.; Wu, X.; Xue, R.; Du, D. Spatio-Temporal Diversity in the Link between Tree Radial Growth and Remote Sensing Vegetation Index of Qinghai Spruce on the Northeastern Margin of the Tibetan Plateau. Forests 2023, 14, 260. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Status | Declining (D) | Non-Declining (N) |
|---|---|---|
| N° stands | 20 | 42 |
| Total area (km2) | 6.96 | 7.63 |
| Elevation (m a.s.l.) | 1222 ± 211 | 1314 ± 160 |
| Maximum temperature (°C) | 14.76 ± 0.04 a | 12.75 ± 0.04 b |
| Minimum temperature (°C) | 5.04 ± 0.03 a | 3.52 ± 0.02 b |
| Precipitation (mm) | 829.7 ± 4.9 a | 1024.8 ± 4.6 b |
| Vapor pressure deficit (kPa) | 0.566 ± 0.002 a | 0.482 ± 0.001 b |
| Climatic water deficit (mm) | 343.0 ± 3.5 a | 220.0 ± 2.2 b |
| Soil moisture (mm) | 85.8 ± 0.7 a | 111.9 ± 0.5 b |
| NDVI | 0.766 ± 0.005 a | 0.779 ± 0.005 a |
| EVI | 1.916 ± 0.060 a | 2.154 ± 0.038 b |
| Tree-ring width (mm) * [10,19,46] | 1.74 ± 0.17 a | 2.57 ± 0.51 b |
| Defoliation 2000 (%) * [10,46] | 18 | 4 |
| Defoliation 2020 (%) * [19] | 40 | 9 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Crespo-Antia, J.P.; Gazol, A.; Pizarro, M.; González de Andrés, E.; Valeriano, C.; Rubio Cuadrado, Á.; Linares, J.C.; Camarero, J.J. Matching Vegetation Indices and Tree Vigor in Pyrenean Silver Fir Stands. Remote Sens. 2024, 16, 4564. https://doi.org/10.3390/rs16234564
Crespo-Antia JP, Gazol A, Pizarro M, González de Andrés E, Valeriano C, Rubio Cuadrado Á, Linares JC, Camarero JJ. Matching Vegetation Indices and Tree Vigor in Pyrenean Silver Fir Stands. Remote Sensing. 2024; 16(23):4564. https://doi.org/10.3390/rs16234564
Chicago/Turabian StyleCrespo-Antia, Juan Pablo, Antonio Gazol, Manuel Pizarro, Ester González de Andrés, Cristina Valeriano, Álvaro Rubio Cuadrado, Juan Carlos Linares, and Jesús Julio Camarero. 2024. "Matching Vegetation Indices and Tree Vigor in Pyrenean Silver Fir Stands" Remote Sensing 16, no. 23: 4564. https://doi.org/10.3390/rs16234564
APA StyleCrespo-Antia, J. P., Gazol, A., Pizarro, M., González de Andrés, E., Valeriano, C., Rubio Cuadrado, Á., Linares, J. C., & Camarero, J. J. (2024). Matching Vegetation Indices and Tree Vigor in Pyrenean Silver Fir Stands. Remote Sensing, 16(23), 4564. https://doi.org/10.3390/rs16234564