
Influence of Spotted Lanternfly (Lycorma delicatula) on Multiple Maple (Acer spp.) Species Canopy Foliar Spectral and Chemical Profiles



{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Leaf Spectral and Tissue Collections
2.3. Spectral Indexes Calculated from Leaf Spectral Data
2.4. Chemical Quantification of Leaf Tissue
2.5. Statistical Analysis
3. Results
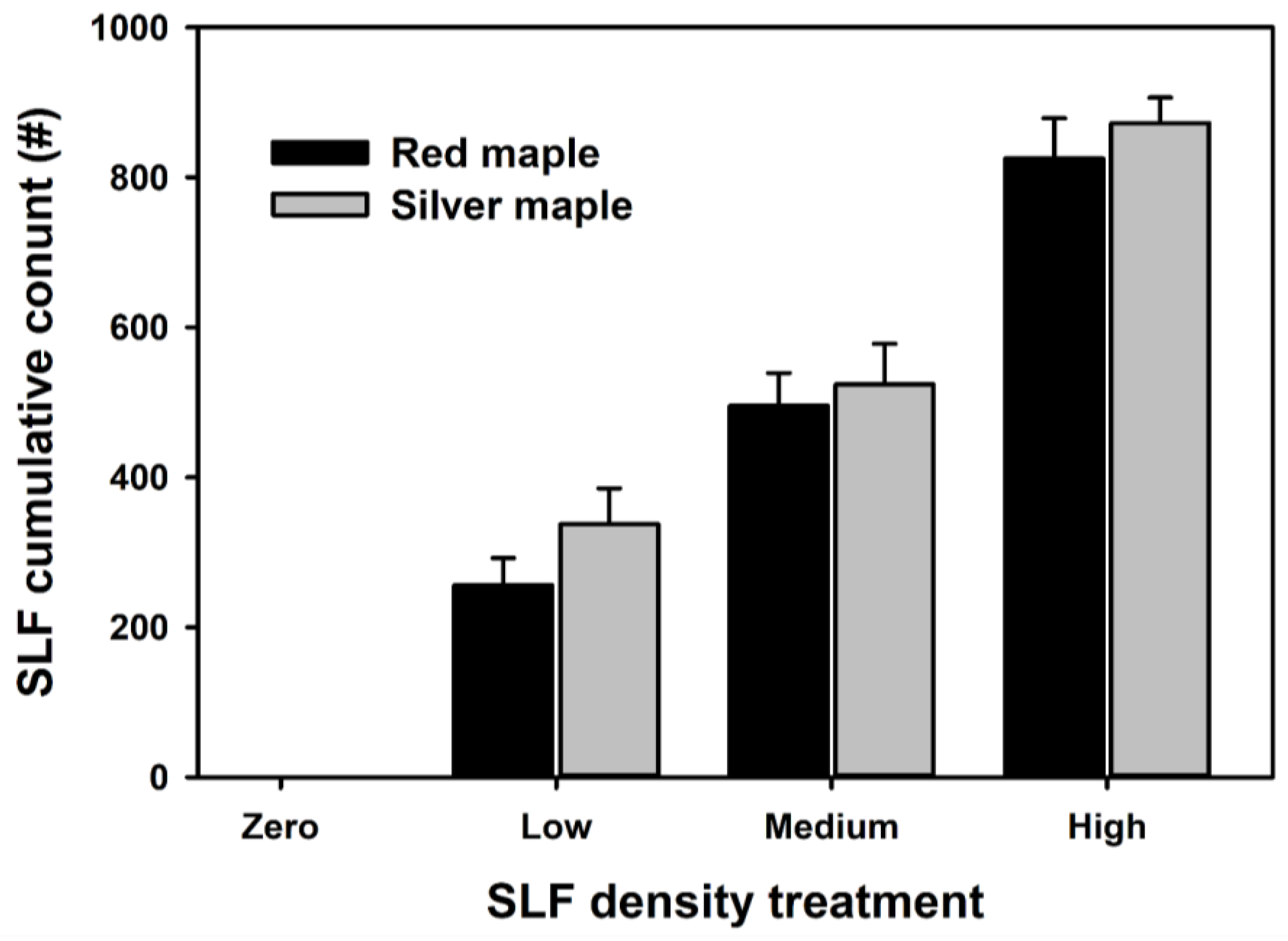
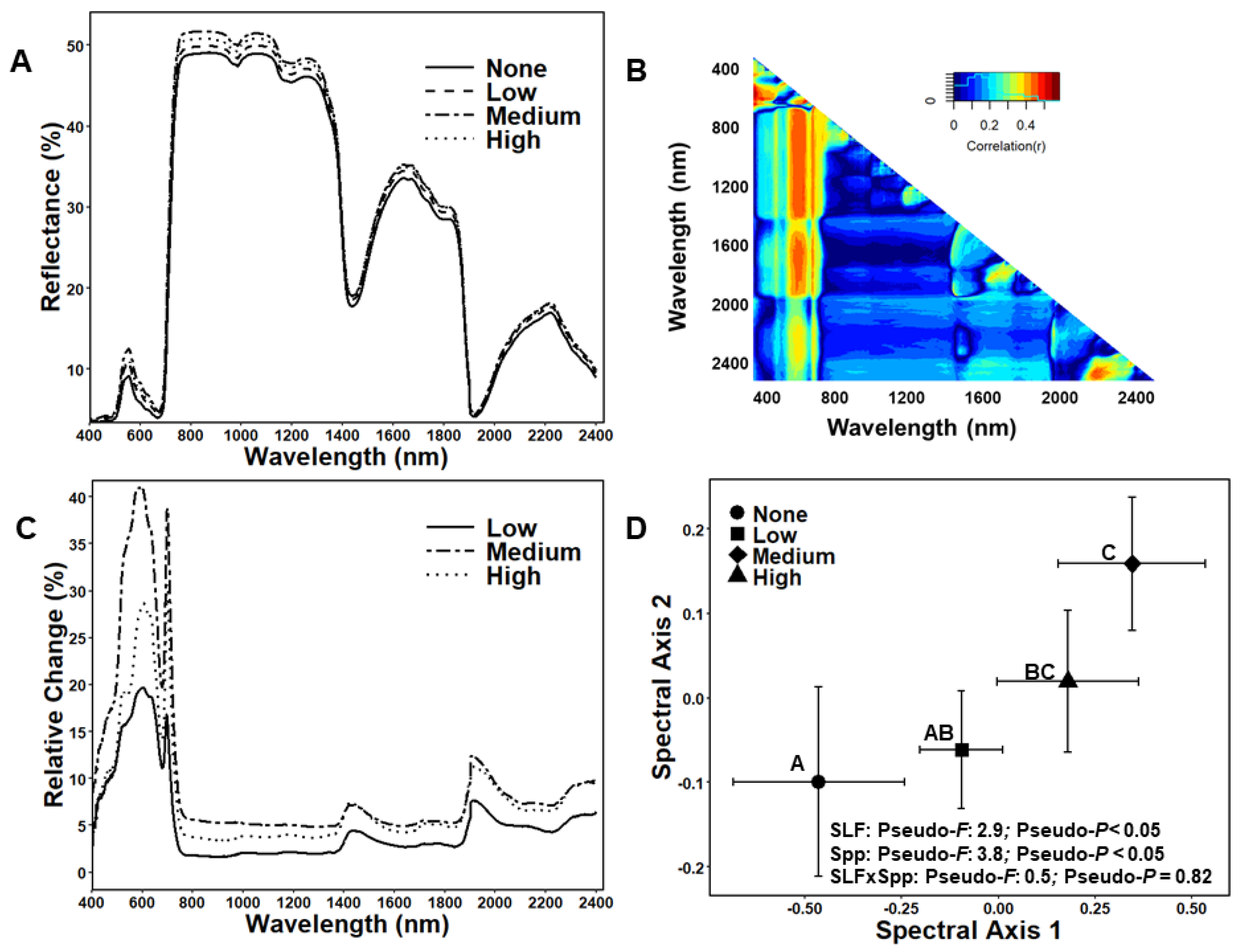
3.1. Foliar Spectral Responses to Different SLF Infestation Levels and Tree Species
3.2. Foliar Spectral Index and Chemical Responses to SLF Infestation Levels
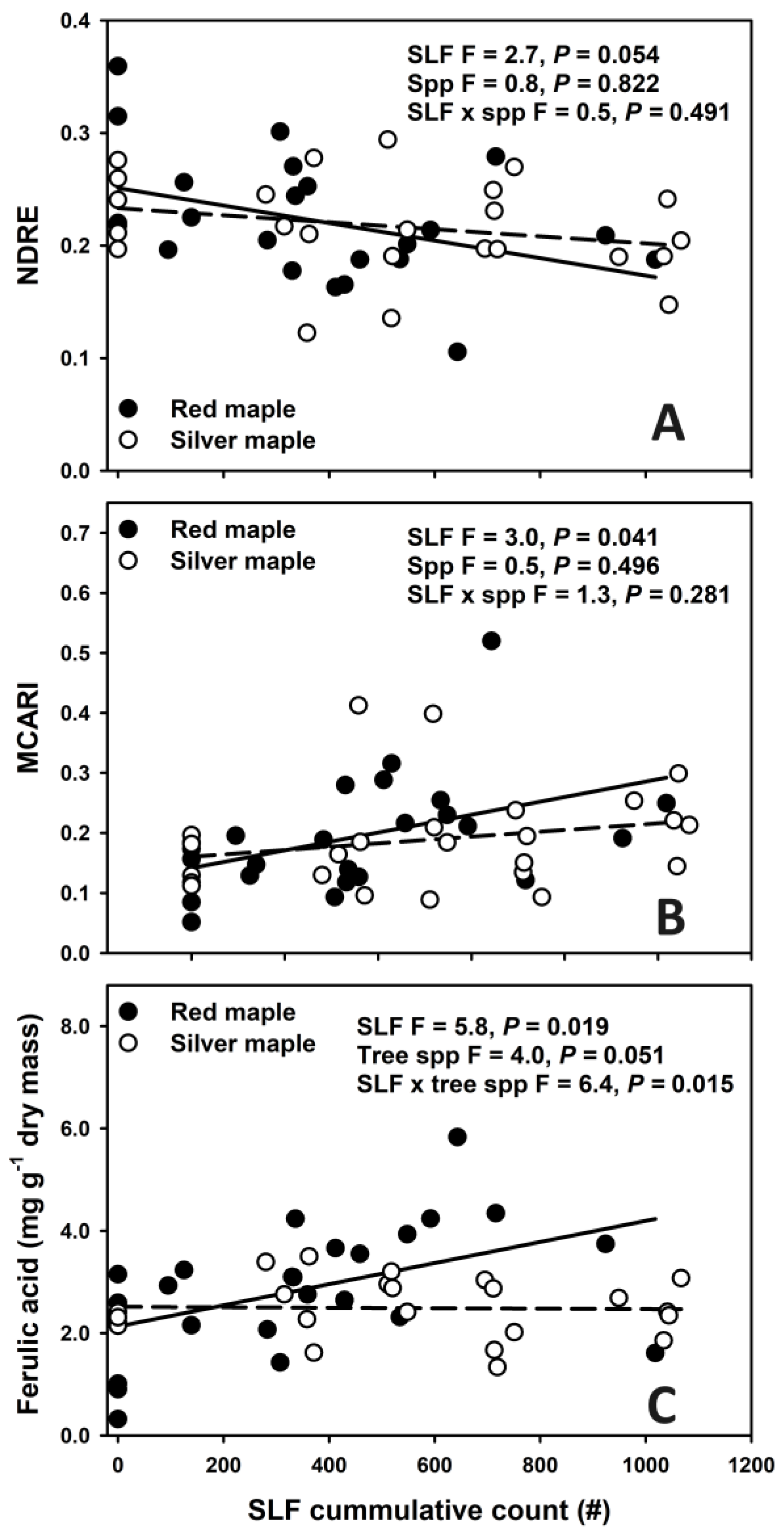
3.3. Relationships of Spectral Indexes and Foliar Chemistry with SLF Cumulative Densities
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Davidson, C.B.; Gottschalk, K.W.; Johnson, J.E. Tree mortality following defoliation by the European gypsy moth (Lymantria dispar L.) in the United States: A review. For. Sci. 1999, 45, 74–84. [Google Scholar] [CrossRef]
- McManus, M.; Csóka, G. History and impact of gypsy moth in North America and comparison to the recent outbreaks in Europe. Acta Silv. Lignaria Hung. 2007, 3, 47–64. [Google Scholar] [CrossRef]
- Bradshaw, C.J.; Leroy, B.; Bellard, C.; Roiz, D.; Albert, C.; Fournier, A.; Barbet-Massin, M.; Salles, J.-M.; Simard, F.; Courchamp, F. Massive yet grossly underestimated global costs of invasive insects. Nat. Commun. 2016, 7, 12986. [Google Scholar] [CrossRef] [PubMed]
- Ward, S.F.; Liebhold, A.M.; Morin, R.S.; Fei, S. Population dynamics of ash across the eastern USA following invasion by emerald ash borer. For. Ecol. Manag. 2021, 479, 118574. [Google Scholar] [CrossRef]
- Fantle-Lepczyk, J.E.; Haubrock, P.J.; Kramer, A.M.; Cuthbert, R.N.; Turbelin, A.J.; Crystal-Ornelas, R.; Diagne, C.; Courchamp, F. Economic costs of biological invasions in the United States. Sci. Total Environ. 2022, 806, 151318. [Google Scholar] [CrossRef]
- Fei, S.; Morin, R.S.; Oswalt, C.M.; Liebhold, A.M. Biomass losses resulting from insect and disease invasions in US forests. Proc. Natl. Acad. Sci. USA 2019, 116, 17371–17376. [Google Scholar] [CrossRef] [PubMed]
- Paini, D.R.; Sheppard, A.W.; Cook, D.C.; De Barro, P.J.; Worner, S.P.; Thomas, M.B. Global threat to agriculture from invasive species. Proc. Natl. Acad. Sci. USA 2016, 113, 7575–7579. [Google Scholar] [CrossRef]
- Du, Z.; Wu, Y.; Zhou, C.; Cao, L.; Ishikawa, T.; Kamitani, S.; Sota, T.; Song, F.; Tian, L.; Cai, W.; et al. Lobal phylogeny and invasion history of the spotted lanternfly revealed by mitochondrial phylogenomics. Evol. App. 2020, 14, 915–930. [Google Scholar] [CrossRef]
- Dara, S.K.; Barringer, L.; Arthurs, S.P. Lycorma delicatula (Hemiptera: Fulgoridae): A New Invasive Pest in the United States. J. Integr. Pest Manag. 2015, 6, 20. [Google Scholar] [CrossRef]
- Barringer, L.; Ciafré, C.M. Worldwide feeding host plants of spotted lanternfly, with significant additions from North America. Environ. Entomol. 2020, 49, 999–1011. [Google Scholar] [CrossRef]
- Urban, J.M. Perspective: Shedding light on spotted lanternfly impacts in the USA. Pest Manag. Sci. 2020, 76, 10–17. [Google Scholar] [CrossRef] [PubMed]
- Murman, K.; Setliff, G.P.; Pugh, C.V.; Toolan, M.J.; Canlas, I.; Cannon, S.; Abreu, L.; Fetchen, M.; Zhang, L.; Warden, M.L.; et al. Distribution, survival, and development of spotted lanternfly on host plants found in North America. Environ. Entomol. 2020, 49, 1270–1281. [Google Scholar] [CrossRef] [PubMed]
- Liu, H. Oviposition substrate selection, egg mass characteristics, host preference, and life history of the spotted lanternfly (Hemiptera: Fulgoridae) in North America. Environ. Entomol. 2019, 48, 1452–1468. [Google Scholar] [CrossRef] [PubMed]
- Spotted Lanternfly. Available online: https://www.aphis.usda.gov/aphis/resources/pests-diseases/hungry-pests/the-threat/spotted-lanternfly/spotted-lanternfly (accessed on 20 February 2024).
- Jung, J.; Jung, S.; Byeon, D.; Lee, W. Model-based prediction of potential distribution of the invasive insect pest, spotted lanternfly Lycorma delicatula (Hemiptera: Fulgoridae), by using CLIMEX. J. Asia-Pac. Biodivers. 2017, 10, 532–538. [Google Scholar] [CrossRef]
- Wakie, T.T.; Neven, L.G.; Yee, W.L.; Lu, Z. The establishment risk of Lycorma delicatula (Hemiptera: Fulgoridae) in the United States and globally. J. Econ. Entomol. 2019, 113, 306–314. [Google Scholar] [CrossRef] [PubMed]
- Harper, J.K.; Stone, W.; Kelsey, T.W.; Kime, L.F. Potential Economic Impact of the Spotted Lanternfly on Agriculture and Forestry in Pennsylvania; Center for Rural Pennsylvania: Harrisburg, PA, USA, 2019; pp. 1–84. [Google Scholar]
- Mahlein, A.K.; Steiner, U.; Hillnhütter, C.; Dehne, H.W.; Oerke, E.C. Hyperspectral imaging for small-scale analysis of symptoms caused by different sugar beet diseases. Plant Methods 2012, 8, 3. [Google Scholar] [CrossRef] [PubMed]
- Couture, J.J.; Serbin, S.P.; Townsend, P.A. Spectroscopic sensitivity of real-time, rapidly induced phytochemical change in response to damage. New Phytol. 2013, 198, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Couture, J.J.; Singh, A.; Rubert-Nason, K.F.; Serbin, S.P.; Lindroth, R.L.; Townsend, P.A. Spectroscopic determination of ecologically relevant plant secondary metabolites. Methods Ecol. Evol. 2016, 7, 1402–1412. [Google Scholar] [CrossRef]
- Mutka, A.M.; Bart, R.S. Image-based phenotyping of plant disease symptoms. Front. Plant Sci. 2015, 5, 734. [Google Scholar] [CrossRef] [PubMed]
- Zarco-Tejada, P.J.; Camino, C.; Beck, P.S.A.; Calderon, R.; Hornero, A.; Hernández-Clemente, R.; Kattenborn, T.; Montes-Borrego, M.; Susca, L.; Morelli, M.; et al. Previsual symptoms of Xylella fastidiosa infection revealed in spectral plant-trait alterations. Nat. Plants 2018, 4, 432–439. [Google Scholar] [CrossRef] [PubMed]
- Cotrozzi, L.; Couture, J.J. Hyperspectral assessment of plant responses to multi-stress environments: Prospects for managing protected agrosystems. Plants People Planet 2020, 2, 244–258. [Google Scholar] [CrossRef]
- Gold, K.M.; Townsend, P.A.; Chlus, A.; Herrmann, I.; Couture, J.J.; Larson, E.R.; Gevens, A.J. Hyperspectral measurements enable pre-symptomatic detection and differentiation of contrasting physiological effects of late blight and early blight in potato. Remote Sens. 2020, 12, 286. [Google Scholar] [CrossRef]
- Cotrozzi, L. Spectroscopic detection of forest diseases: A review (1970–2020). J. For. Res. 2022, 33, 21–38. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Townsend, P.A.; Pellegrini, E.; Nali, C.; Couture, J.J. Reflectance spectroscopy: A novel approach to better understand and monitor the impact of air pollution on Mediterranean plants. Environ. Sci. Pollut. Res. 2018, 25, 8249–8267. [Google Scholar] [CrossRef] [PubMed]
- Nay, J.; Burchfield, E.; Gilligan, J. A machine-learning approach to forecasting remotely sensed vegetation health. Int. J. Remote Sens. 2018, 39, 1800–1816. [Google Scholar] [CrossRef]
- Lavely, E.; Iavorivska, L.; Uyi, O.; Eissenstat, D.M.; Walsh, B.; Primka, E.J.; Harper, J.; Hoover, K. Impacts of short-term feeding by spotted lanternfly (Lycorma delicatula) on ecophysiology of young hardwood trees in a common garden. Front. Insect Sci. 2022, 2, 1080124. [Google Scholar] [CrossRef]
- Hoover, K.; Iavorivska, L.; Lavely, E.K.; Uyi, O.; Walsh, B.; Swackhamer, E.; Johnson, A.; Eissenstat, D.M. Effects of long-term feeding by spotted lanternfly (Hemiptera: Fulgoridae) on ecophysiology of common hardwood host trees. Environ. Entomol. 2023, 52, 888–899. [Google Scholar] [CrossRef] [PubMed]
- Mason, C.J.; Walsh, B.; Keller, J.; Couture, J.J.; Calvin, D.; Urban, J.M. Fidelity and timing of spotted lanternfly (Hemiptera: Fulgoridae) attack patterns on ornamental trees in the suburban landscape. Environ. Entomol. 2020, 49, 1427–1436. [Google Scholar] [CrossRef] [PubMed]
- Calvin, D.D.; Rost, J.; Keller, J.; Crawford, S.; Walsh, B.; Bosold, M.; Urban, J. Seasonal activity of spotted lanternfly (Hemiptera: Fulgoridae) in Southeast Pennsylvania. Environ. Entomol. 2023, 52, 1108–1125. [Google Scholar] [CrossRef]
- Colovic, M.; Yu, K.; Todorovic, M.; Cantore, V.; Hamze, M.; Albrizio, R.; Stellacci, A.M. Hyperspectral vegetation indices to assess water and nitrogen status of sweet maize crop. Agronomy 2022, 12, 2181. [Google Scholar] [CrossRef]
- Daughtry, C.S.; Walthall, C.L.; Kim, M.S.; De Colstoun, E.B.; McMurtrey, J.E., III. Estimating corn leaf chlorophyll concentration from leaf and canopy reflectance. Remote Sens. Environ. 2000, 74, 229–239. [Google Scholar] [CrossRef]
- Gamon, J.A.; Serrano, L.; Surfus, J.S. The photochemical reflectance index: An optical indicator of photosynthetic radiation use efficiency across species, functional types, and nutrient levels. Oecologia 1997, 112, 492–501. [Google Scholar] [CrossRef]
- Letts, M.G.; Phelan, C.A.; Johnson, D.R.E.; Rood, S.B. Seasonal photosynthetic gas exchange and leaf reflectance characteristics of male and female cottonwoods in a riparian woodland. Tree Physiol. 2008, 28, 1037–1048. [Google Scholar] [CrossRef] [PubMed]
- Nour, V.; Trandafir, I.; Cosmulescu, S. HPLC determination of phenolic acids, flavonoids and juglone in walnut leaves. J. Chromatogr. Sci. 2013, 51, 883–890. [Google Scholar] [CrossRef] [PubMed]
- Pellegrini, E.; Campanella, A.; Paolocci, M.; Trivellini, A.; Gennai, C.; Muganu, M.; Nali, C.; Lorenzini, G. Functional leaf traits and diurnal dynamics of photosynthetic parameters predict the behavior of grapevine varieties towards ozone. PLoS ONE 2015, 10, e0135056. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Hernandez, J.; Gonzalez-Castro, M.J.; Vazquez-Blanco, M.E.; Vazquez-Oderiz, M.L.; Simal-Lozano, J. HPLC determination of sugars and starch in green beans. J. Food Sci. 1994, 59, 1048–1049. [Google Scholar] [CrossRef]
- Smith, A.M.; Zeeman, S.C. Quantification of starch in plant tissues. Nat. Protoc. 2006, 1, 1342–1345. [Google Scholar] [CrossRef] [PubMed]
- Vu, J.C.V.; Gesch, R.W.; Pennanen, A.H.; Allen, L.H.; Boote, K.J.; Bowes, G. Soybean photosynthesis, Rubisco, and carbohydrate enzymes function at supraoptimal temperatures in elevated CO2. J. Plant Physiol. 2001, 158, 295–307. [Google Scholar] [CrossRef]
- Cotrozzi, L.; Remorini, D.; Pellegrini, E.; Landi, M.; Massai, R.; Nali, C.; Guidi, L.; Lorenzini, G. Variations in physiological and biochemical traits of oak seedlings grown under drought and ozone stress. Physiol. Plant. 2016, 157, 69–84. [Google Scholar] [CrossRef] [PubMed]
- Rock, B.N.; Hoshizaki, T.; Miller, J.R. Comparison of In Situ and Airborne Spectral Measurements of the Blue Shift Associated with Forest Decline. Remote Sens. Environ. 1988, 24, 109–127. [Google Scholar] [CrossRef]
- Filella, I.; Peñuelas, J. The red edge position and shape as indicators of plant chlorophyll content, biomass and hydric status. Int. J. Remote Sens. 1994, 15, 1459–1470. [Google Scholar] [CrossRef]
- Harner, A.D.; Leach, H.L.; Briggs, L.; Centinari, M. Prolonged phloem feeding by the spotted lanternfly, an invasive planthopper, alters resource allocation and inhibits gas exchange in grapevines. Plant Direct 2022, 6, e452. [Google Scholar] [CrossRef] [PubMed]
- Rehman, F.; Khan, F.A.; Badruddin, S.M.A. Role of Phenolics in Plant Defense Against Insect Herbivory. In Chemistry of Phytopotentials: Health, Energy and Environmental Perspectives; Khemani, L., Srivastava, M., Srivastava, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2012; pp. 309–313. [Google Scholar] [CrossRef]
- Walling, L.L. The myriad plant responses to herbivores. J. Plant Growth Regul. 2000, 19, 195–216. [Google Scholar] [CrossRef] [PubMed]
- Thompson, G.A.; Goggin, F.L. Transcriptomics and functional genomics of plant defence induction by phloem-feeding insects. J. Exp. Bot. 2006, 57, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Kerchev, P.I.; Fenton, B.; Foyer, C.H.; Hancock, R.D. Plant responses to insect herbivory: Interactions between photosynthesis, reactive oxygen species and hormonal signaling pathways. Plant Cell Environ. 2012, 35, 441–453. [Google Scholar] [CrossRef] [PubMed]
- Pincebourde, S.; Ngao, J. The impact of phloem feeding insects on leaf ecophysiology varies with leaf age. Front. Plant Sci. 2021, 12, 625689. [Google Scholar] [CrossRef]
- Wang, Y.; Cai, Q.N.; Zhang, Q.W.; Han, Y. Effect of the secondary substances from wheat on the growth and digestive physiology of cotton bollworm Helicoverpa armigera (Lepidoptera: Noctuidae). Eur. J. Entomol. 2006, 103, 255–258. [Google Scholar] [CrossRef]
- Czerniewicz, P.; Leszczynski, B.; Chrzanowski, G.; Sempruch, C.; Sytykiewicz, H. Effects of host plant phenolics on spring migration of bird cherry-oat aphid (Rhopalosiphum padi L.). Allelopath. J. 2011, 27, 309–316. [Google Scholar]
- Zhang, M.; Fang, T.; Pu, G.; Sun, X.; Zhou, X.; Cai, Q. Xenobiotic metabolism of plant secondary compounds in the English grain aphid, Sitobion avenae (F.) (Hemiptera: Aphididae). Pestic. Biochem. Physiol. 2013, 107, 44–49. [Google Scholar] [CrossRef]
- Abuley, I.K.; Pedersen, H.A.; Lekfeldt, J.D.S.; Fomsgaard, I.S.; Ravnskov, S. Metabolite profiling of Solanum tuberosum reveals a differential response to Phytophthora infestans dependent on host resistance and pathogen isolate. Plant Pathol. 2023, 72, 924–932. [Google Scholar] [CrossRef]
- Gao, P.; Qi, Y.; Li, L.; Yang, S.; Liu, J.; Wei, H.; Huang, F.; Yu, L. Amorphophallus muelleri activates ferulic acid and phenylpropane biosynthesis pathways to defend against Fusarium solani infection. Front. Plant Sci. 2023, 14, 1207970. [Google Scholar] [CrossRef] [PubMed]
- Kokaly, R.F.; Asner, G.P.; Ollinger, S.V.; Martin, M.E.; Wessman, C.A. Characterizing canopy biochemistry from imaging spectroscopy and its application to ecosystem studies. Remote Sens. Environ. 2009, 113, S78–S91. [Google Scholar] [CrossRef]
- Kokaly, R.F.; Skidmore, A.K. Plant phenolics and absorption features in vegetation reflectance spectra near 1.66 μm. Int. J. Appl. Earth Obs. Geoinf. 2015, 43, 55–83. [Google Scholar] [CrossRef]
- Fine, P.V.A.; Salazar, D.; Martin, R.E.; Metz, M.R.; Misiewicz, T.M.; Asner, G.P. Exploring the links between secondary metabolites and leaf spectral reflectance in a diverse genus of Amazonian trees. Ecosphere 2021, 12, e03362. [Google Scholar] [CrossRef]
- Pozdnyakova, L.; Oudemans, P.V.; Hughes, M.G.; Giménez, D. Estimation of spatial and spectral properties of phytophthora root rot and its effects on cranberry yield. Comput. Electron. Agric. 2002, 37, 57–70. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, M.; Zhu, J.; Geng, S. Spectral prediction of Phytophthora infestans infection on tomatoes using artificial neural network (ANN). Int. J. Remote Sens. 2008, 29, 1693–1706. [Google Scholar] [CrossRef]
- Reynolds, G.J.; Windels, C.E.; MacRae, I.V.; Laguette, S. Remote sensing for assessing rhizoctonia crown and root rot severity in sugar beet. Plant Dis. 2012, 96, 497–505. [Google Scholar] [CrossRef] [PubMed]
- Omer, M.; Locke, J.C.; Frantz, J.M. Using leaf temperature as a nondestructive procedure to detect root rot stress in geranium. HortTechnology 2015, 17, 532–536. [Google Scholar] [CrossRef]
- Calamita, F.; Imran, H.A.; Vescovo, L.; Mekhalfi, M.L.; La Porta, N. Early identification of root rot disease by using hyperspectral reflectance: The case of pathosystem grapevine/armillaria. Remote Sens. 2021, 13, 2436. [Google Scholar] [CrossRef]
- Weksler, S.; Rozenstein, O.; Haish, N.; Moshelion, M.; Wallach, R.; Ben-Dor, E. Pepper plants leaf spectral reflectance changes as a result of root rot damage. Remote Sens. 2021, 13, 980. [Google Scholar] [CrossRef]
- Peron-Danaher, R.; Cotrozzi, L.; Masjedi, A.; Enders, L.S.; Krupke, C.H.; Mickelbart, M.V.; Couture, J.J. Drought stress affects spectral separation of maize infested by western corn rootworm. Agronomy 2023, 13, 2562. [Google Scholar] [CrossRef]
- Lowe, A.; Harrison, N.; French, A.P. Hyperspectral image analysis techniques for the detection and classification of the early onset of plant disease and stress. Plant Methods 2017, 13, 80. [Google Scholar] [CrossRef] [PubMed]
- Susič, N.; Žibrat, U.; Širca, S.; Strajnar, P.; Razinger, J.; Knapič, M.; Vončina, A.; Urek, G.; Stare, B.G. Discrimination between abiotic and biotic drought stress in tomatoes using hyperspectral imaging. Sens. Actuators B Chem. 2018, 273, 842–852. [Google Scholar] [CrossRef]
- Williams, L.J.; Cavender-Bares, J.; Townsend, P.A.; Couture, J.J.; Wang, Z.; Stefanski, A.; Messier, C.; Reich, P.B. Remote spectral detection of biodiversity effects on forest biomass. Nat. Ecol. Evol. 2021, 5, 46–54. [Google Scholar] [CrossRef] [PubMed]
- Leach, H.; Walsh, B.; Urban, J. Evaluation of insecticides for control of spotted lanternfly in ornamental nursery crop, 2019. Arthropod Manag. Tests 2021, 46, tsab043. [Google Scholar] [CrossRef]
- Leach, H.; Urban, J. Management and control of the spotted lanternfly, Lycorma delicatula (Hemiptera: Fulgoridae), in the United States. Annu. Rev. Entomol. 2023, 68, 151–167. [Google Scholar]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Joll, E.G.; Ginzel, M.D.; Hoover, K.; Couture, J.J. Influence of Spotted Lanternfly (Lycorma delicatula) on Multiple Maple (Acer spp.) Species Canopy Foliar Spectral and Chemical Profiles. Remote Sens. 2024, 16, 2706. https://doi.org/10.3390/rs16152706
Joll EG, Ginzel MD, Hoover K, Couture JJ. Influence of Spotted Lanternfly (Lycorma delicatula) on Multiple Maple (Acer spp.) Species Canopy Foliar Spectral and Chemical Profiles. Remote Sensing. 2024; 16(15):2706. https://doi.org/10.3390/rs16152706
Chicago/Turabian StyleJoll, Elisabeth G., Matthew D. Ginzel, Kelli Hoover, and John J. Couture. 2024. "Influence of Spotted Lanternfly (Lycorma delicatula) on Multiple Maple (Acer spp.) Species Canopy Foliar Spectral and Chemical Profiles" Remote Sensing 16, no. 15: 2706. https://doi.org/10.3390/rs16152706
APA StyleJoll, E. G., Ginzel, M. D., Hoover, K., & Couture, J. J. (2024). Influence of Spotted Lanternfly (Lycorma delicatula) on Multiple Maple (Acer spp.) Species Canopy Foliar Spectral and Chemical Profiles. Remote Sensing, 16(15), 2706. https://doi.org/10.3390/rs16152706