
Phenology and Plant Functional Type Link Optical Properties of Vegetation Canopies to Patterns of Vertical Vegetation Complexity


Abstract

1. Introduction
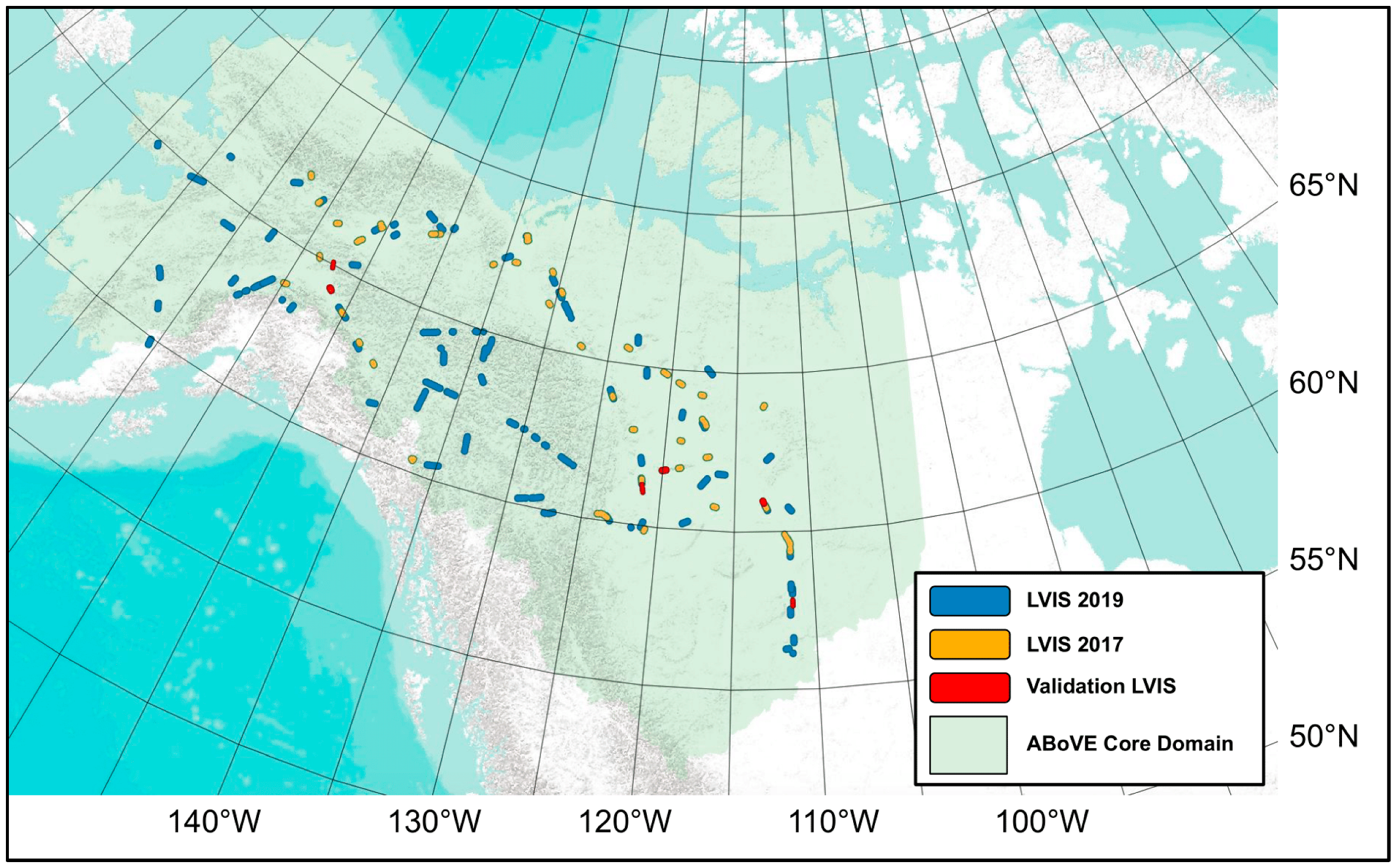
2. Data and Study Area
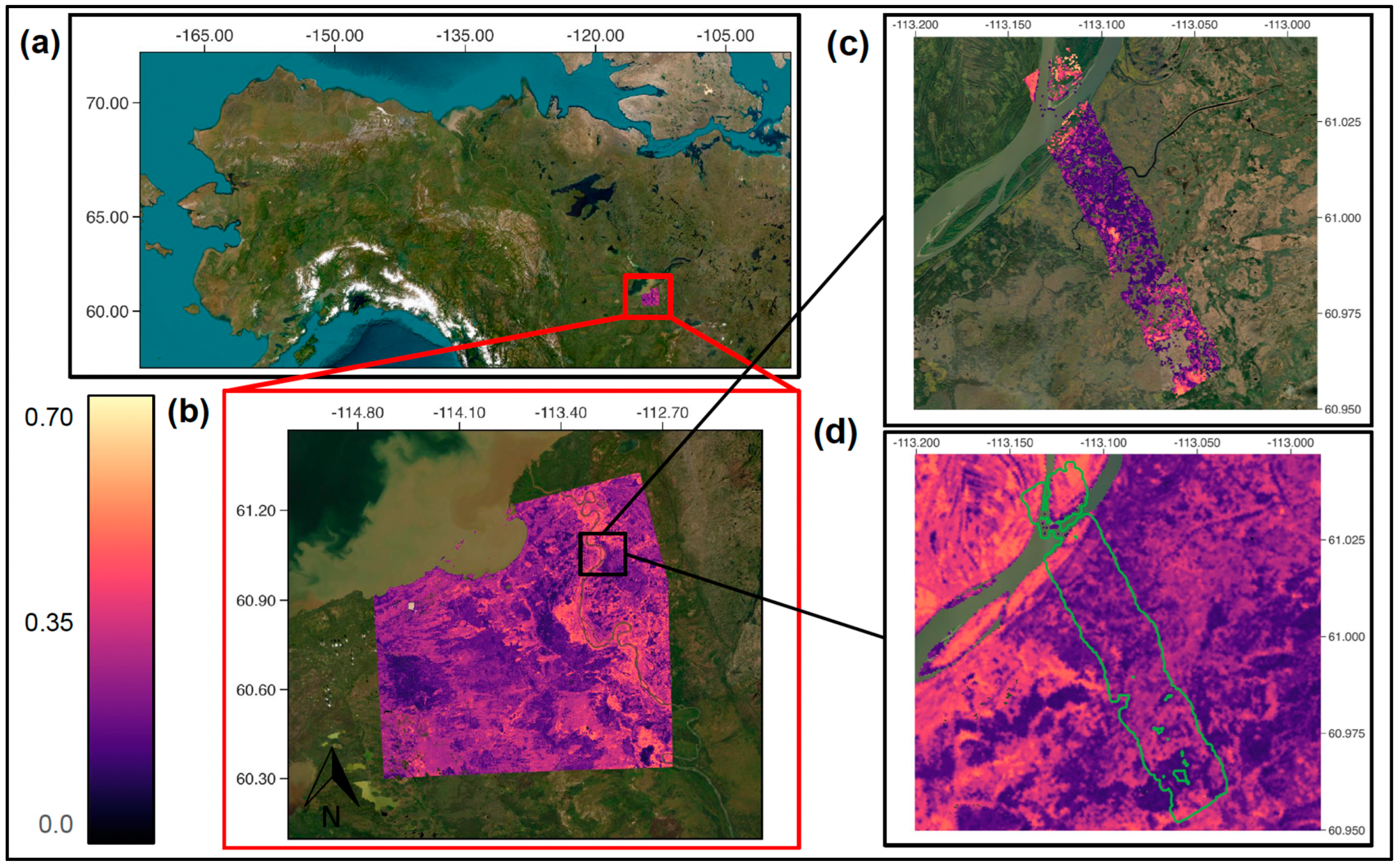
2.1. Study Area
2.2. In Situ Data
2.3. Full-Waveform LiDAR
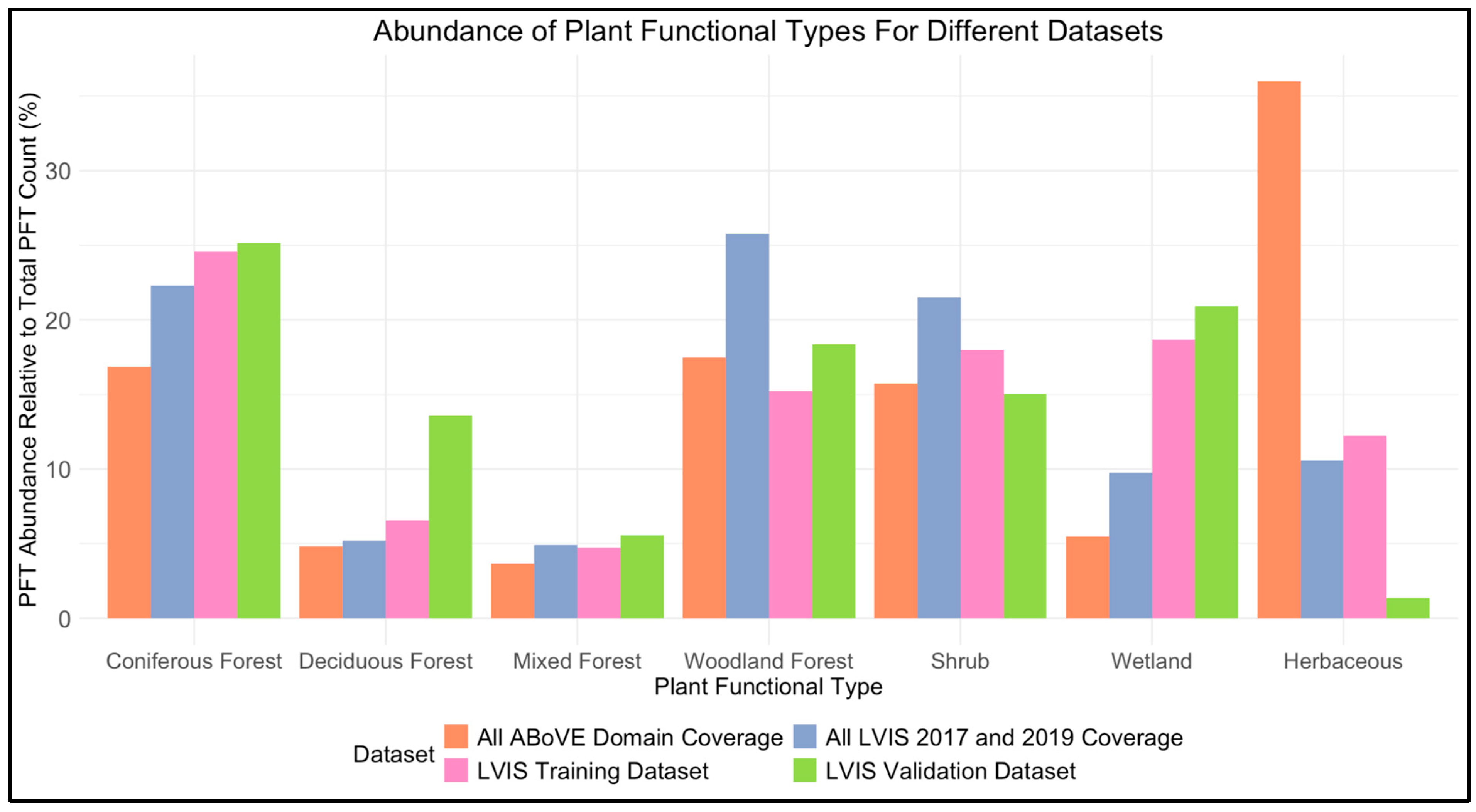
2.4. Plant Functional Type Data
2.5. Optical Data
2.6. Environmental Data
3. Methods
3.1. NEON In Situ Growth Form and Structure
3.2. Full-Waveform Processing
3.3. LiDAR Stratified Sampling
3.4. Plant Functional Type Groupings
3.5. Green Season Length
3.6. HLS Optical Data
3.7. Environmental Variables
3.8. Modeling, and Validation
4. Results
4.1. Plant Identity, Canopy Height, and Vertical Complexity
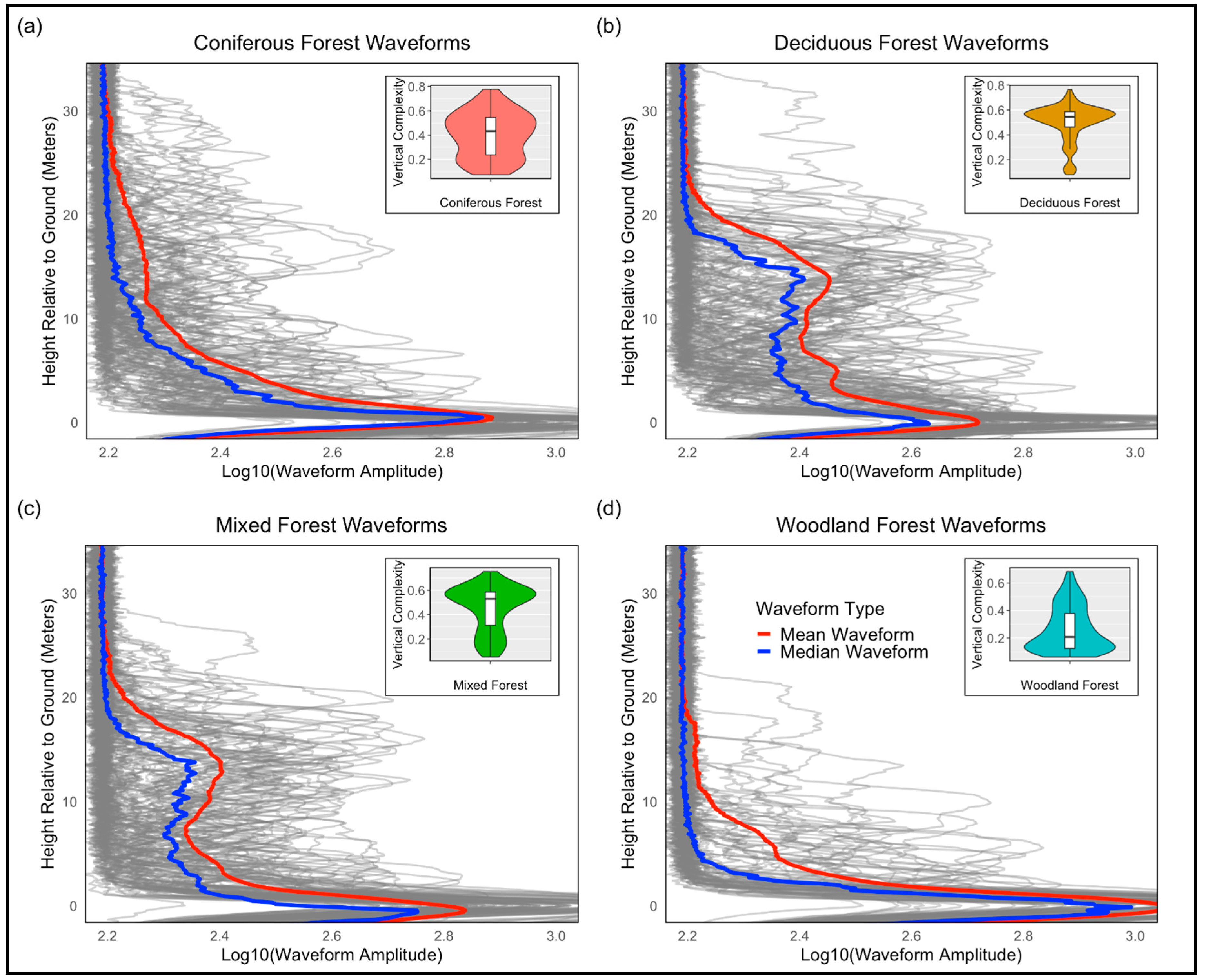
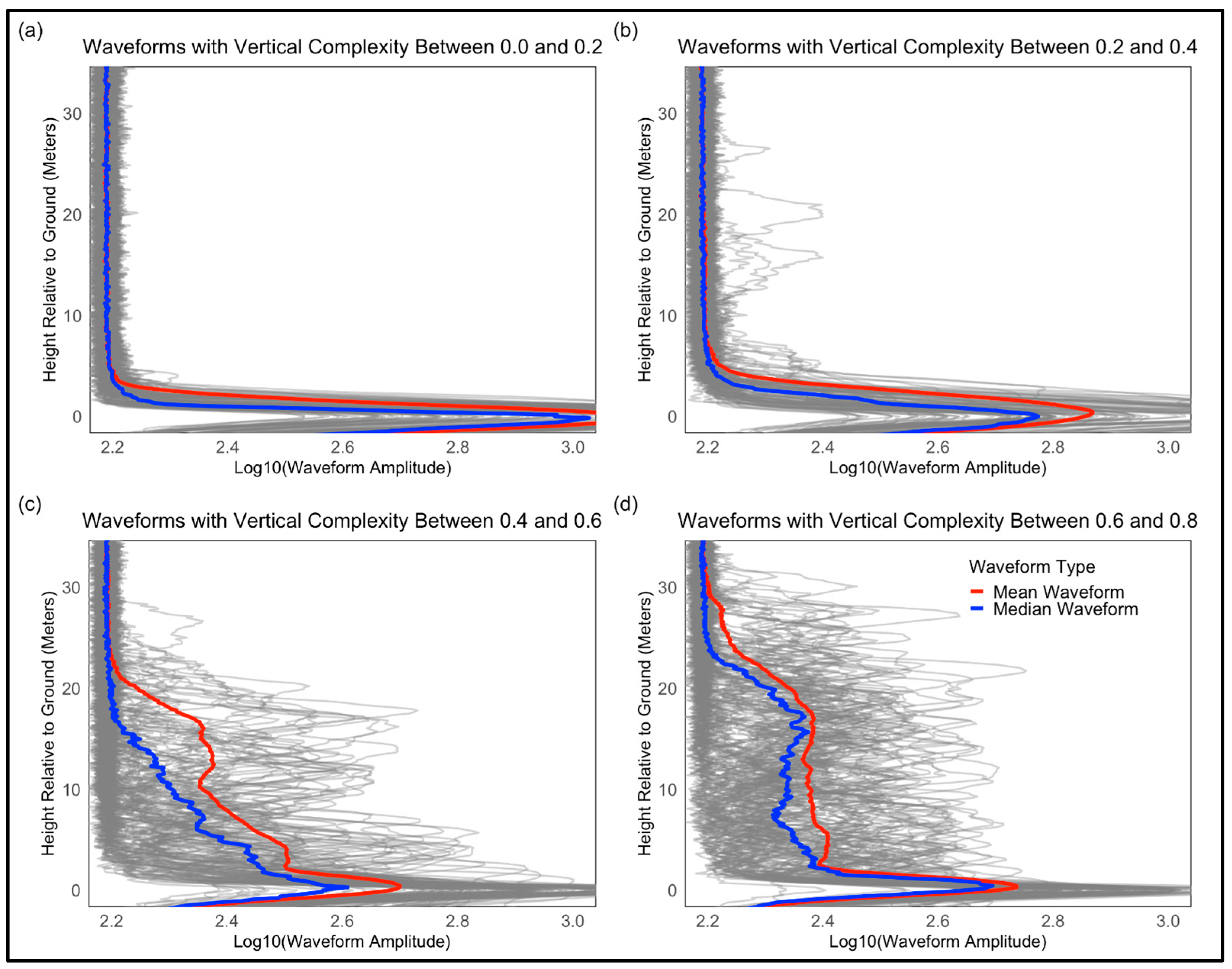
4.2. LiDAR Waveforms, Plant Functional Types, and Vertical Complexity
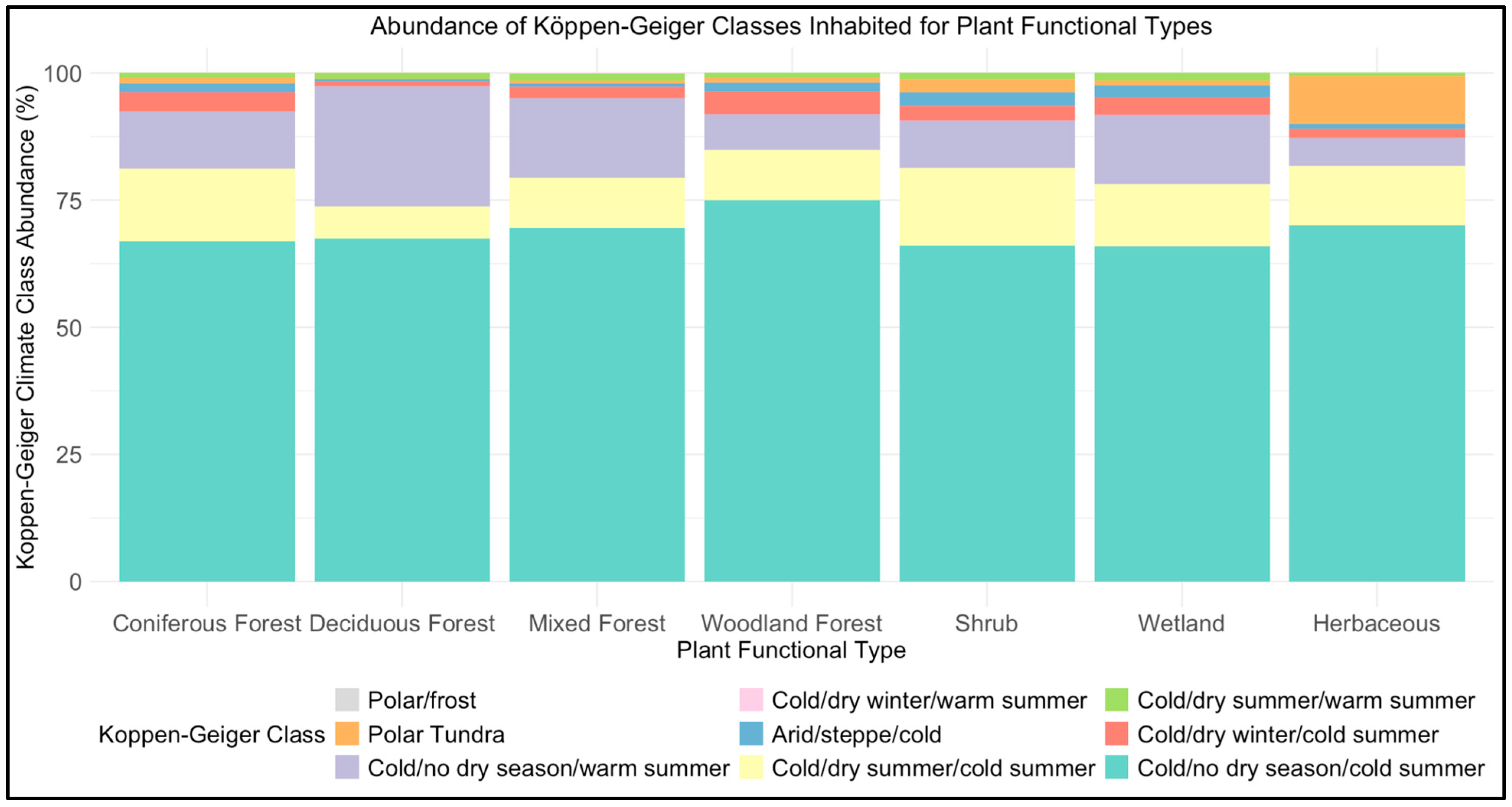
4.3. Environmental Sensitivities of Vertical Complexity
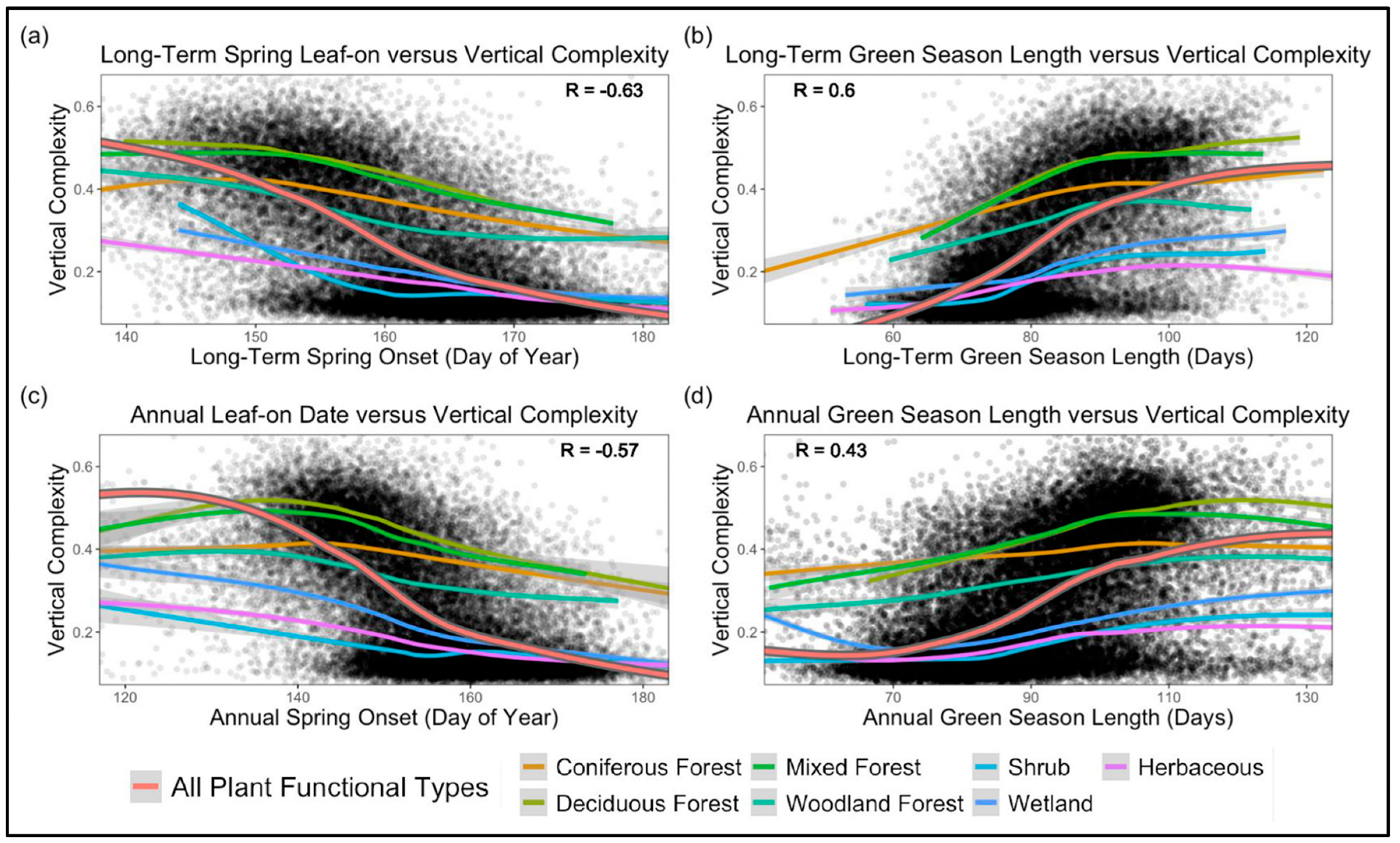
4.4. Phenology and Vertical Complexity
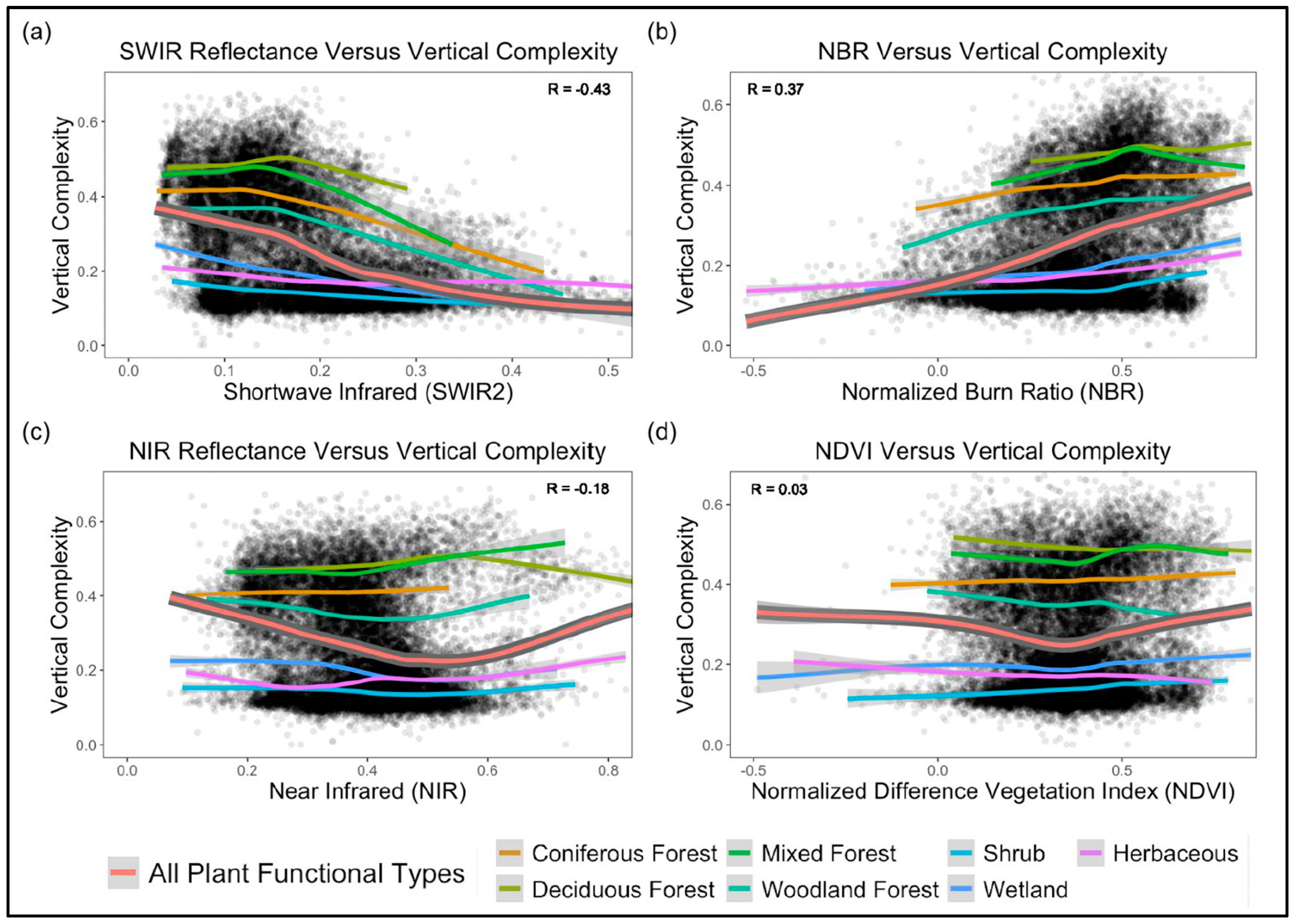
4.5. Canopy Optical Properties and Vertical Complexity
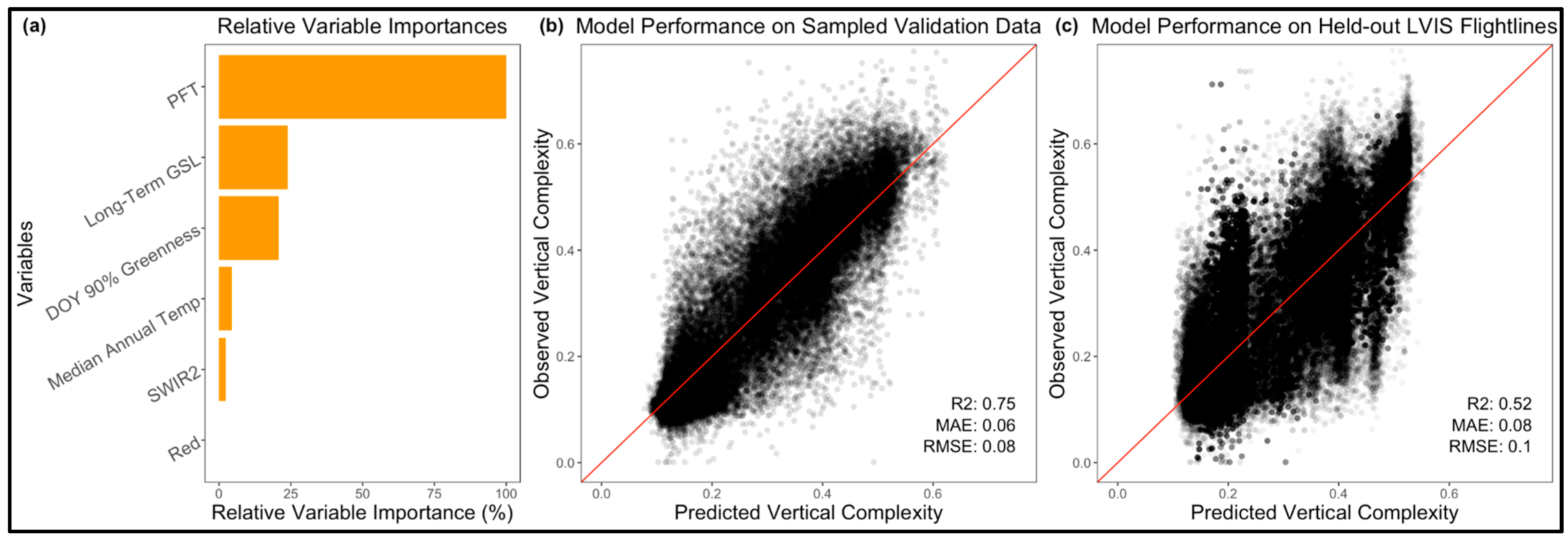
4.6. Random Forest Predictions
5. Discussion
- The relationship between canopy height and VC
- Waveform structure and vertical complexity
- Phenology and VC
- Climate and VC
- Random forest predictions of VC
5.1. Study Limitations
5.1.1. Spatial Sampling of LVIS Flightlines
5.1.2. Summer-Only LiDAR Observations
5.1.3. Temporal Misalignment between PFT and LiDAR Datasets
5.1.4. Drawbacks to Modeling at 30 m Resolution
5.2. Recommendations
5.2.1. Improved Spatial Sampling
5.2.2. Further Experiments with Optical Methods
5.2.3. Potential for SWIR Phenology
5.2.4. The Need for Multi-Scale Analysis
6. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McElhinny, C.; Gibbons, P.; Brack, C.; Bauhus, J. Forest and Woodland Stand Structural Complexity: Its Definition and Measurement. For. Ecol. Manag. 2005, 218, 1–24. [Google Scholar] [CrossRef]
- Gough, C.M.; Atkins, J.W.; Fahey, R.T.; Hardiman, B.S. High Rates of Primary Production in Structurally Complex Forests. Ecology 2019, 100, e02864. [Google Scholar] [CrossRef] [PubMed]
- Farwell, L.S.; Gudex-Cross, D.; Anise, I.E.; Bosch, M.J.; Olah, A.M.; Radeloff, V.C.; Razenkova, E.; Rogova, N.; Silveira, E.M.O.; Smith, M.M.; et al. Satellite Image Texture Captures Vegetation Heterogeneity and Explains Patterns of Bird Richness. Remote Sens. Environ. 2021, 253, 112175. [Google Scholar] [CrossRef]
- Goetz, S.J.; Steinberg, D.; Betts, M.G.; Holmes, R.T.; Doran, P.J.; Dubayah, R.; Hofton, M. Lidar Remote Sensing Variables Predict Breeding Habitat of a Neotropical Migrant Bird. Ecology 2010, 91, 1569–1576. [Google Scholar] [CrossRef] [PubMed]
- Murphy, B.A.; May, J.A.; Butterworth, B.J.; Andresen, C.G.; Desai, A.R. Unraveling Forest Complexity: Resource Use Efficiency, Disturbance, and the Structure-Function Relationship. J. Geophys. Res. Biogeosci. 2022, 127, e2021JG006748. [Google Scholar] [CrossRef]
- Fischer, R.; Knapp, N.; Bohn, F.; Shugart, H.H.; Huth, A. The Relevance of Forest Structure for Biomass and Productivity in Temperate Forests: New Perspectives for Remote Sensing. Surv. Geophys. 2019, 40, 709–734. [Google Scholar] [CrossRef]
- Box, E.O. Plant Functional Types and Climate at the Global Scale. J. Veg. Sci. 1996, 7, 309–320. [Google Scholar] [CrossRef]
- Fahey, R.T.; Atkins, J.W.; Gough, C.M.; Hardiman, B.S.; Nave, L.E.; Tallant, J.M.; Nadehoffer, K.J.; Vogel, C.; Scheuermann, C.M.; Stuart-Haëntjens, E. Defining a Spectrum of Integrative Trait-Based Vegetation Canopy Structural Types. Fahey-2019-Ecology Letters-Wiley Online Library. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/ele.13388 (accessed on 12 April 2024).
- Ustin, S.L.; Gamon, J.A. Remote Sensing of Plant Functional Types. New Phytol. 2010, 186, 795–816. [Google Scholar] [CrossRef]
- Givnish, T.J.; Wong, S.C.; Stuart-Williams, H.; Holloway-Phillips, M.; Farquhar, G.D. Determinants of Maximum Tree Height in Eucalyptus Species along a Rainfall Gradient in Victoria, Australia. Ecology 2014, 95, 2991–3007. [Google Scholar] [CrossRef]
- Tao, S.; Guo, Q.; Li, C.; Givnish, Z.; Fang, J. Global Patterns and Determinants of Forest Canopy Height. Ecology 2016, 97, 3265–3270. [Google Scholar] [CrossRef]
- Jucker, T.; Bongalov, B.; Burslem, D.F.R.P.; Nilus, R.; Dalponte, M.; Lewis, S.L.; Phillips, O.L.; Qie, L.; Coomes, D.A. Topography Shapes the Structure, Composition and Function of Tropical Forest Landscapes. Ecol. Lett. 2018, 21, 989–1000. [Google Scholar] [CrossRef] [PubMed]
- Ehbrecht, M.; Seidel, D.; Annighöfer, P.; Kreft, H.; Köhler, M.; Zemp, D.C.; Puettmann, K.; Nilus, R.; Babweteera, F.; Willim, K.; et al. Global Patterns and Climatic Controls of Forest Structural Complexity. Nat. Commun. 2021, 12, 519. [Google Scholar] [CrossRef] [PubMed]
- Ryan, M.G.; Phillips, N.; Bond, B.J. The Hydraulic Limitation Hypothesis Revisited. Plant Cell Environ. 2006, 29, 367–381. [Google Scholar] [CrossRef] [PubMed]
- Intergovernmental Panel on Climate Change (IPCC) Climate Change 2021—The Physical Science Basis: Working Group I Contribution to the Sixth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2023.
- Andreu-Hayles, L.; D’Arrigo, R.; Anchukaitis, K.J.; Beck, P.S.A.; Frank, D.; Goetz, S. Varying Boreal Forest Response to Arctic Environmental Change at the Firth River, Alaska. Environ. Res. Lett. 2011, 6, 045503. [Google Scholar] [CrossRef]
- Welp, L.R.; Randerson, J.T.; Liu, H.P. The Sensitivity of Carbon Fluxes to Spring Warming and Summer Drought Depends on Plant Functional Type in Boreal Forest Ecosystems. Agric. For. Meteorol. 2007, 147, 172–185. [Google Scholar] [CrossRef]
- Stinziano, J.R.; Way, D.A. Combined Effects of Rising [CO2] and Temperature on Boreal Forests: Growth, Physiology and Limitations. Botany 2014, 92, 425–436. [Google Scholar] [CrossRef]
- Dial, R.J.; Maher, C.T.; Hewitt, R.E.; Sullivan, P.F. Sufficient Conditions for Rapid Range Expansion of a Boreal Conifer. Nature 2022, 608, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Maher, C.T.; Dial, R.J.; Pastick, N.J.; Hewitt, R.E.; Jorgenson, M.T.; Sullivan, P.F. The Climate Envelope of Alaska’s Northern Treelines: Implications for Controlling Factors and Future Treeline Advance. Ecography 2021, 44, 1710–1722. [Google Scholar] [CrossRef]
- Scheffer, M.; Hirota, M.; Holmgren, M.; Van Nes, E.H.; Chapin, F.S. Thresholds for Boreal Biome Transitions. Proc. Natl. Acad. Sci. USA 2012, 109, 21384–21389. [Google Scholar] [CrossRef]
- Reich, P.B.; Bermudez, R.; Montgomery, R.A.; Rich, R.L.; Rice, K.E.; Hobbie, S.E.; Stefanski, A. Even Modest Climate Change May Lead to Major Transitions in Boreal Forests. Nature 2022, 608, 540–545. [Google Scholar] [CrossRef]
- Whitman, E.; Parisien, M.-A.; Thompson, D.K.; Flannigan, M.D. Short-Interval Wildfire and Drought Overwhelm Boreal Forest Resilience. Sci. Rep. 2019, 9, 18796. [Google Scholar] [CrossRef] [PubMed]
- Alexander, H.D.; Mack, M.C.; Goetz, S.; Beck, P.S.A.; Belshe, E.F. Implications of Increased Deciduous Cover on Stand Structure and Aboveground Carbon Pools of Alaskan Boreal Forests. Ecosphere 2012, 3, art45. [Google Scholar] [CrossRef]
- Mekonnen, Z.A.; Riley, W.J.; Randerson, J.T.; Grant, R.F.; Rogers, B.M. Expansion of High-Latitude Deciduous Forests Driven by Interactions between Climate Warming and Fire. Nat. Plants 2019, 5, 952–958. [Google Scholar] [CrossRef] [PubMed]
- Mack, M.C.; Walker, X.J.; Johnstone, J.F.; Alexander, H.D.; Melvin, A.M.; Jean, M.; Miller, S.N. Carbon Loss from Boreal Forest Wildfires Offset by Increased Dominance of Deciduous Trees. Science 2021, 372, 280–283. [Google Scholar] [CrossRef] [PubMed]
- Fisher, J.I.; Mustard, J.F.; Vadeboncoeur, M.A. Green Leaf Phenology at Landsat Resolution: Scaling from the Field to the Satellite. Remote Sens. Environ. 2006, 100, 265–279. [Google Scholar] [CrossRef]
- Melaas, E.K.; Sulla-Menashe, D.; Gray, J.M.; Black, T.A.; Morin, T.H.; Richardson, A.D.; Friedl, M.A. Multisite Analysis of Land Surface Phenology in North American Temperate and Boreal Deciduous Forests from Landsat. Remote Sens. Environ. 2016, 186, 452–464. [Google Scholar] [CrossRef]
- Linkosalo, T.; Häkkinen, R.; Terhivuo, J.; Tuomenvirta, H.; Hari, P. The Time Series of Flowering and Leaf Bud Burst of Boreal Trees (1846–2005) Support the Direct Temperature Observations of Climatic Warming. Agric. For. Meteorol. 2009, 149, 453–461. [Google Scholar] [CrossRef]
- Jeganathan, C.; Dash, J.; Atkinson, P.M. Remotely Sensed Trends in the Phenology of Northern High Latitude Terrestrial Vegetation, Controlling for Land Cover Change and Vegetation Type. Remote Sens. Environ. 2014, 143, 154–170. [Google Scholar] [CrossRef]
- Buermann, W.; Bikash, P.R.; Jung, M.; Burn, D.H.; Reichstein, M. Earlier Springs Decrease Peak Summer Productivity in North American Boreal Forests. Environ. Res. Lett. 2013, 8, 024027. [Google Scholar] [CrossRef]
- Chen, X.; Wang, W.; Chen, J.; Zhu, X.; Shen, M.; Gan, L.; Cao, X. Does Any Phenological Event Defined by Remote Sensing Deserve Particular Attention? An Examination of Spring Phenology of Winter Wheat in Northern China. Ecol. Indic. 2020, 116, 106456. [Google Scholar] [CrossRef]
- Keenan, T.F.; Gray, J.; Friedl, M.A.; Toomey, M.; Bohrer, G.; Hollinger, D.Y.; Munger, J.W.; O’Keefe, J.; Schmid, H.P.; Wing, I.S.; et al. Net Carbon Uptake Has Increased through Warming-Induced Changes in Temperate Forest Phenology. Nat. Clim. Chang. 2014, 4, 598–604. [Google Scholar] [CrossRef]
- Park, T.; Ganguly, S.; Tømmervik, H.; Euskirchen, E.S.; Høgda, K.-A.; Karlsen, S.R.; Brovkin, V.; Nemani, R.R.; Myneni, R.B. Changes in Growing Season Duration and Productivity of Northern Vegetation Inferred from Long-Term Remote Sensing Data. Environ. Res. Lett. 2016, 11, 084001. [Google Scholar] [CrossRef]
- Sulla-Menashe, D.; Woodcock, C.E.; Friedl, M.A. Canadian Boreal Forest Greening and Browning Trends: An Analysis of Biogeographic Patterns and the Relative Roles of Disturbance versus Climate Drivers. Environ. Res. Lett. 2018, 13, 014007. [Google Scholar] [CrossRef]
- Guo, W.; Liu, H.; Wu, X. Vegetation Greening Despite Weakening Coupling between Vegetation Growth and Temperature over the Boreal Region. J. Geophys. Res. Biogeosci. 2018, 123, 2376–2387. [Google Scholar] [CrossRef]
- Melaas, E.K.; Sulla-Menashe, D.; Friedl, M.A. Multidecadal Changes and Interannual Variation in Springtime Phenology of North American Temperate and Boreal Deciduous Forests. Geophys. Res. Lett. 2018, 45, 2679–2687. [Google Scholar] [CrossRef]
- LaRue, E.A.; Fahey, R.T.; Alveshere, B.C.; Atkins, J.W.; Bhatt, P.; Buma, B.; Chen, A.; Cousins, S.; Elliott, J.M.; Elmore, A.J.; et al. A Theoretical Framework for the Ecological Role of Three-Dimensional Structural Diversity. Front. Ecol. Environ. 2023, 21, 4–13. [Google Scholar] [CrossRef]
- Atkins, J.W.; Bhatt, P.; Carrasco, L.; Francis, E.; Garabedian, J.E.; Hakkenberg, C.R.; Hardiman, B.S.; Jung, J.; Koirala, A.; LaRue, E.A.; et al. Integrating Forest Structural Diversity Measurement into Ecological Research. Ecosphere 2023, 14, e4633. [Google Scholar] [CrossRef]
- Marselis, S.M.; Tang, H.; Armston, J.D.; Calders, K.; Labrière, N.; Dubayah, R. Distinguishing Vegetation Types with Airborne Waveform Lidar Data in a Tropical Forest-Savanna Mosaic: A Case Study in Lopé National Park, Gabon. Remote Sens. Environ. 2018, 216, 626–634. [Google Scholar] [CrossRef]
- Gu, Z.; Cao, S.; Sanchez-Azofeifa, G.A. Using LiDAR Waveform Metrics to Describe and Identify Successional Stages of Tropical Dry Forests. Int. J. Appl. Earth Obs. Geoinf. 2018, 73, 482–492. [Google Scholar] [CrossRef]
- Sun, C.; Cao, S.; Sanchez-Azofeifa, G.A. Mapping Tropical Dry Forest Age Using Airborne Waveform LiDAR and Hyperspectral Metrics. Int. J. Appl. Earth Obs. Geoinf. 2019, 83, 101908. [Google Scholar] [CrossRef]
- Dubayah, R.; Armston, J.; Healey, S.P.; Bruening, J.M.; Patterson, P.L.; Kellner, J.R.; Duncanson, L.; Saarela, S.; Ståhl, G.; Yang, Z.; et al. GEDI Launches a New Era of Biomass Inference from Space. Environ. Res. Lett. 2022, 17, 095001. [Google Scholar] [CrossRef]
- Schneider, F.D.; Ferraz, A.; Hancock, S.; Duncanson, L.I.; Dubayah, R.O.; Pavlick, R.P.; Schimel, D.S. Towards Mapping the Diversity of Canopy Structure from Space with GEDI. Environ. Res. Lett. 2020, 15, 115006. [Google Scholar] [CrossRef]
- Diaz-Kloch, N.; Murray, D.L. Bridging the Gap: Comprehensive Boreal Forest Complexity Mapping through LVIS Full-Waveform LiDAR, Single-Year and Time Series Landsat Imagery. Remote Sens. 2023, 15, 5274. [Google Scholar] [CrossRef]
- Dube, T.; Mutanga, O. Investigating the Robustness of the New Landsat-8 Operational Land Imager Derived Texture Metrics in Estimating Plantation Forest Aboveground Biomass in Resource Constrained Areas. ISPRS J. Photogramm. Remote Sens. 2015, 108, 12–32. [Google Scholar] [CrossRef]
- Jansen, V.S.; Kolden, C.A.; Schmalz, H.J. The Development of Near Real-Time Biomass and Cover Estimates for Adaptive Rangeland Management Using Landsat 7 and Landsat 8 Surface Reflectance Products. Remote Sens. 2018, 10, 1057. [Google Scholar] [CrossRef]
- Matasci, G.; Hermosilla, T.; Wulder, M.A.; White, J.C.; Coops, N.C.; Hobart, G.W.; Zald, H.S.J. Large-Area Mapping of Canadian Boreal Forest Cover, Height, Biomass and Other Structural Attributes Using Landsat Composites and Lidar Plots. Remote Sens. Environ. 2018, 209, 90–106. [Google Scholar] [CrossRef]
- Li, Z.; Bi, S.; Hao, S.; Cui, Y. Aboveground Biomass Estimation in Forests with Random Forest and Monte Carlo-Based Uncertainty Analysis. Ecol. Indic. 2022, 142, 109246. [Google Scholar] [CrossRef]
- Lang, N.; Schindler, K.; Wegner, J.D. Country-Wide High-Resolution Vegetation Height Mapping with Sentinel-2. Remote Sens. Environ. 2019, 233, 111347. [Google Scholar] [CrossRef]
- Guo, Y.; Chen, S.; Li, X.; Cunha, M.; Jayavelu, S.; Cammarano, D.; Fu, Y. Machine Learning-Based Approaches for Predicting SPAD Values of Maize Using Multi-Spectral Images. Remote Sens. 2022, 14, 1337. [Google Scholar] [CrossRef]
- Kearney, S.P.; Porensky, L.M.; Augustine, D.J.; Gaffney, R.; Derner, J.D. Monitoring Standing Herbaceous Biomass and Thresholds in Semiarid Rangelands from Harmonized Landsat 8 and Sentinel-2 Imagery to Support within-Season Adaptive Management. Remote Sens. Environ. 2022, 271, 112907. [Google Scholar] [CrossRef]
- Lang, N.; Jetz, W.; Schindler, K.; Wegner, J.D. A High-Resolution Canopy Height Model of the Earth. Nat. Ecol. Evol. 2023, 7, 1778–1789. [Google Scholar] [CrossRef] [PubMed]
- Guo, Y.; Xiao, Y.; Hao, F.; Zhang, X.; Chen, J.; de Beurs, K.; He, Y.; Fu, Y.H. Comparison of Different Machine Learning Algorithms for Predicting Maize Grain Yield Using UAV-Based Hyperspectral Images. Int. J. Appl. Earth Obs. Geoinf. 2023, 124, 103528. [Google Scholar] [CrossRef]
- Wang, C.; Feng, M.-C.; Yang, W.-D.; Ding, G.-W.; Sun, H.; Liang, Z.-Y.; Xie, Y.-K.; Qiao, X.-X. Impact of Spectral Saturation on Leaf Area Index and Aboveground Biomass Estimation of Winter Wheat. Spectrosc. Lett. 2016, 49, 241–248. [Google Scholar] [CrossRef]
- Zhao, P.; Lu, D.; Wang, G.; Wu, C.; Huang, Y.; Yu, S. Examining Spectral Reflectance Saturation in Landsat Imagery and Corresponding Solutions to Improve Forest Aboveground Biomass Estimation. Remote Sens. 2016, 8, 469. [Google Scholar] [CrossRef]
- Magiera, A.; Feilhauer, H.; Waldhardt, R.; Wiesmair, M.; Otte, A. Modelling Biomass of Mountainous Grasslands by Including a Species Composition Map. Ecol. Indic. 2017, 78, 8–18. [Google Scholar] [CrossRef]
- Guo, Y.; Fu, Y.; Hao, F.; Zhang, X.; Wu, W.; Jin, X.; Robin Bryant, C.; Senthilnath, J. Integrated Phenology and Climate in Rice Yields Prediction Using Machine Learning Methods. Ecol. Indic. 2021, 120, 106935. [Google Scholar] [CrossRef]
- Meyer, H.; Reudenbach, C.; Wöllauer, S.; Nauss, T. Importance of Spatial Predictor Variable Selection in Machine Learning Applications—Moving from Data Reproduction to Spatial Prediction. Ecol. Model. 2019, 411, 108815. [Google Scholar] [CrossRef]
- Loboda, T.V.; Carroll, M.L. ABoVE: Study Domain and Standard Reference Grids; Version 2; ORNL DAAC: Oak Ridge, TN, USA, 2017. [Google Scholar] [CrossRef]
- Wang, J.A.; Sulla-Menashe, D.; Woodcock, C.E.; Sonnentag, O.; Keeling, R.F.; Friedl, M.A. Extensive Land Cover Change across Arctic–Boreal Northwestern North America from Disturbance and Climate Forcing. Glob. Chang. Biol. 2020, 26, 807–822. [Google Scholar] [CrossRef]
- NEON (National Ecological Observatory Network). Vegetation Structure (DP1.10098.001), RELEASE-2024. Available online: https://data.neonscience.org/data-products/DP1.10098.001/RELEASE-2024 (accessed on 18 April 2024). [CrossRef]
- Blair, J.B.; Rabine, D.L.; Hofton, M.A. The Laser Vegetation Imaging Sensor: A Medium-Altitude, Digitisation-Only, Airborne Laser Altimeter for Mapping Vegetation and Topography. ISPRS J. Photogramm. Remote Sens. 1999, 54, 115–122. [Google Scholar] [CrossRef]
- Blair, J.B.; Hofton, M. ABoVE LVIS L1B Geolocated Return Energy Waveforms, Version 1 [Data Set]. Boulder, Colorado USA. NASA National Snow and Ice Data Center Distributed Active Archive Center. 2018. Available online: https://nsidc.org/data/ablvis1b/versions/1 (accessed on 18 April 2024). [CrossRef]
- Blair, J.B.; Hofton, M. LVIS Facility L1B Geolocated Return Energy Waveforms, Version 1 [Data Set]. Boulder, Colorado USA. NASA National Snow and Ice Data Center Distributed Active Archive Center. 2020. Available online: https://nsidc.org/data/lvisf1b/versions/1 (accessed on 18 April 2024). [CrossRef]
- Blair, J.B.; Hofton, M. ABoVE LVIS L2 Geolocated Surface Elevation Product, Version 1 [Data Set]. Boulder, Colorado USA. NASA National Snow and Ice Data Center Distributed Active Archive Center. 2018. Available online: https://nsidc.org/data/ablvis2/versions/1 (accessed on 18 April 2024). [CrossRef]
- Blair, J.B.; Hofton, M. LVIS Facility L2 Geolocated Surface Elevation and Canopy Height Product, Version 1 [Data Set]. Boulder, Colorado USA. NASA National Snow and Ice Data Center Distributed Active Archive Center. 2020. Available online: https://nsidc.org/data/lvisf2/versions/1 (accessed on 18 April 2024). [CrossRef]
- Wang, J.A.; Sulla-Menashe, D.; Woodcock, C.E.; Sonnentag, O.; Keeling, R.F.; Friedl, M.A. ABoVE: Landsat-Derived Annual Dominant Land Cover Across ABoVE Core Domain, 1984–2014; ORNL DAAC: Oak Ridge, TN, USA, 2019. [Google Scholar] [CrossRef]
- Zhu, Z.; Woodcock, C.E. Continuous Change Detection and Classification of Land Cover Using All Available Landsat Data. Remote Sens. Environ. 2014, 144, 152–171. [Google Scholar] [CrossRef]
- Friedl, M. MuSLI Multi-Source Land Surface Phenology Yearly North America 30 m V011 [Data Set]. NASA EOSDIS Land Processes Distributed Active Archive Center. 2021. Available online: https://lpdaac.usgs.gov/products/mslsp30nav011/ (accessed on 18 April 2024). [CrossRef]
- Masek, J.; Ju, J.J.; Roger, S.; Skakun, E.; Vermote, M.; Claverie, J.; Dungan, Z.; Yin, B.; Freitag, C.J. HLS Operational Land Imager Surface Reflectance and TOA Brightness Daily Global 30 m v2.0. 2021, Distributed by NASA EOSDIS Land Processes DAAC. Available online: https://doi.org/10.5067/HLS/HLSL30.002 (accessed on 18 April 2024).
- Claverie, M.; Ju, J.; Masek, J.G.; Dungan, J.L.; Vermote, E.F.; Roger, J.-C.; Skakun, S.V.; Justice, C. The Harmonized Landsat and Sentinel-2 Surface Reflectance Data Set. Remote Sens. Environ. 2018, 219, 145–161. [Google Scholar] [CrossRef]
- Thornton, M.M.; Shrestha, R.; Wei, Y.; Thornton, P.E.; Kao, S.-C.; Wilson, B.E. Daymet: Daily Surface Weather Data on a 1-Km Grid for North America; Version 4 R1; ORNL DAAC: Oak Ridge, TN, USA, 2022. [Google Scholar] [CrossRef]
- Present and Future Köppen-Geiger Climate Classification Maps at 1-Km Resolution|Scientific Data. Available online: https://www.nature.com/articles/sdata2018214 (accessed on 3 June 2024).
- Qiu, S.; Zhu, Z.; He, B. Fmask 4.0: Improved Cloud and Cloud Shadow Detection in Landsats 4–8 and Sentinel-2 Imagery. Remote Sens. Environ. 2019, 231, 111205. [Google Scholar] [CrossRef]
- Liaw, A.; Wiener, M. Classification and Regression by randomForest. R News 2002, 2, 18–22. Available online: https://journal.r-project.org/articles/RN-2002-022/RN-2002-022.pdf (accessed on 18 April 2024).
- Kuhn, M. Building Predictive Models in R Using the Caret Package. J. Stat. Softw. 2008, 28, 1–26. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2023; Available online: https://Www.R-Project.Org/ (accessed on 18 April 2024).
- Probst, P.; Wright, M.N.; Boulesteix, A.-L. Hyperparameters and Tuning Strategies for Random Forest. WIREs Data Min. Knowl. Discov. 2019, 9, e1301. [Google Scholar] [CrossRef]
- Goodbody, T.R.H.; Coops, N.C.; Queinnec, M.; White, J.C.; Tompalski, P.; Hudak, A.T.; Auty, D.; Valbuena, R.; LeBoeuf, A.; Sinclair, I.; et al. sgsR: A Structurally Guided Sampling Toolbox for LiDAR-Based Forest Inventories. For. Int. J. For. Res. 2023, 96, 411–424. [Google Scholar] [CrossRef]
- Atkins, J.W.; Walter, J.A.; Stovall, A.E.L.; Fahey, R.T.; Gough, C.M. Power Law Scaling Relationships Link Canopy Structural Complexity and Height across Forest Types. Funct. Ecol. 2022, 36, 713–726. [Google Scholar] [CrossRef]
- Kattenborn, T.; Schiefer, F.; Frey, J.; Feilhauer, H.; Mahecha, M.D.; Dormann, C.F. Spatially Autocorrelated Training and Validation Samples Inflate Performance Assessment of Convolutional Neural Networks. ISPRS Open J. Photogramm. Remote Sens. 2022, 5, 100018. [Google Scholar] [CrossRef]
- Ploton, P.; Mortier, F.; Réjou-Méchain, M.; Barbier, N.; Picard, N.; Rossi, V.; Dormann, C.; Cornu, G.; Viennois, G.; Bayol, N.; et al. Spatial Validation Reveals Poor Predictive Performance of Large-Scale Ecological Mapping Models. Nat. Commun. 2020, 11, 4540. [Google Scholar] [CrossRef]
- Lee, B.R.; Ibáñez, I. Spring Phenological Escape Is Critical for the Survival of Temperate Tree Seedlings. Funct. Ecol. 2021, 35, 1848–1861. [Google Scholar] [CrossRef]
- Gao, B.-C.; Goetz, A.F.H. Extraction of Dry Leaf Spectral Features from Reflectance Spectra of Green Vegetation. Remote Sens. Environ. 1994, 47, 369–374. [Google Scholar] [CrossRef]
- Atkins, J.W.; Costanza, J.; Dahlin, K.M.; Dannenberg, M.P.; Elmore, A.J.; Fitzpatrick, M.C.; Hakkenberg, C.R.; Hardiman, B.S.; Kamoske, A.; LaRue, E.A.; et al. Scale Dependency of Lidar-Derived Forest Structural Diversity. Methods Ecol. Evol. 2023, 14, 708–723. [Google Scholar] [CrossRef]
- Juchheim, J.; Ehbrecht, M.; Schall, P.; Ammer, C.; Seidel, D. Effect of Tree Species Mixing on Stand Structural Complexity. For. Int. J. For. Res. 2020, 93, 75–83. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Dataset | Variable Name | Description | Units |
|---|---|---|---|
| ABoVE-Pheno | Annual spring onset (ABoVE-Pheno ASO) | The year-specific date that a pixel achieves half-maximum EVI amplitude | Day of year (DOY) |
| ABoVE-Pheno | Long-term spring onset (LTSO) | The multi-year mean of the date that a pixel achieves half-maximum EVI amplitude | Day of year (DOY) |
| ABoVE-Pheno | Annual autumn onset (ABoVE-Pheno AAO) | The year-specific date that a pixel drops below half-maximum EVI amplitude | Day of year (DOY) |
| ABoVE-Pheno | Long-term autumn onset (LTAO) | Multi-year mean of the date that a pixel drops below half-maximum EVI amplitude | Day of year (DOY) |
| ABoVE-Pheno | maxEVI | Annual maximum EVI signal | |
| MS-LSP | ASO | The year-specific date that a pixel achieves 50% EVI amplitude | Day of year (DOY) |
| MS-LSP | AAO | The year-specific date that a pixel drops below 50% EVI amplitude | Day of year (DOY) |
| MS-LSP | OGMx | The year-specific date that a pixel achieves 90% EVI amplitude | Day of year (DOY) |
| MS-LSP | OGMn | The year-specific date that a pixel drops below 90% EVI amplitude | Day of year (DOY) |
| MS-LSP | Peak | The year-specific date that a pixel drops below 10% EVI amplitude | Day of year (DOY) |
| HLS | SWIR1 | Landsat 8 shortwave infrared band 1 | |
| HLS | SWIR2 | Landsat 8 shortwave infrared band 2 | |
| HLS | NIR | Landsat 8 near-infrared band | |
| HLS | NBR | Normalized burn ratio | |
| HLS | NDVI | Normalized difference vegetation index | |
| Daymet | MAT | Annual median of maximum daily temperature | °C |
| Daymet | MnAT | Annual mean of maximum daily temperature | °C |
| Daymet | MdMnAT | Annual median of minimum daily temperature | °C |
| Daymet | AMT | Annual minimum of daily minimum temperature | °C |
| Daymet | PRCP | Total annual precipitation | mm |
| ABoVE-LULC Class | PFT Groups | NEON Growth Group | Height Range |
|---|---|---|---|
| Coniferous Forest | Coniferous Forest | Mature Tree | >3 m |
| Deciduous Forest | Deciduous Forest | Mature Tree | >3 m |
| Mixed Forest | Mixed Forest | Mature Tree | >3 m |
| Woodland | Woodland | Mature Tree | >3 m |
| Low Shrub | Shrub | Shrub Form | 5–30 cm |
| Tall Shrub | Shrub | Shrub Form | Between 50 cm and 3 m |
| Open Shrub | Shrub | Shrub Form | Less than 3 m |
| Herbaceous | Herbaceous | n/a | n/a |
| Tussock Tundra | Herbaceous | n/a | n/a |
| Sparsely Vegetated | Herbaceous | n/a | n/a |
| Fen | Wetland | n/a | n/a |
| Bog | Wetland | n/a | n/a |
| Shallows/littoral | n/a | n/a | n/a |
| Barren | n/a | n;a | n/a |
| Water | n/a | n/a | n/a |
| Predictor Variable | Model A | Model B | Model C | Model D |
|---|---|---|---|---|
| PFT | 100% | 100% | 100% | 100% |
| ASO | 26.67% | 26.38% | ||
| LTSO | 48% | |||
| AGSL | 15.40% | 3.29% | ||
| LTGSL | 46.85% | 23.87% | ||
| ABoVE-Pheno ASO | 14.40% | |||
| MaxEVI | 18.17% | |||
| OGMx | 23.59% | 25.52% | 20.81% | |
| OGI | 25.70% | 5.96% | ||
| Peak | 15.91% | 16.43% | ||
| SWIR1 | 10.13% | 11.22% | ||
| SWIR2 | 10.80% | 14.27% | 2.258% | 16.23% |
| NIR | 0.00% | |||
| Red | 0.000% | |||
| NBR | 2.78% | 5.64% | ||
| NDVI | ||||
| PRCP | 0.000% | 0.00% | 2.19% | |
| MAT | 6.73% | 4.39% | 4.48% | |
| MnAT | 13.11% | 5.62% | 7.56% | |
| MdMnAT | 7.37% | 4.22% | ||
| AMT |
| Validation Method | Model A | Model B | Model C | Model D |
|---|---|---|---|---|
| K-fold cross-validation | R2: 0.68 MAE: 0.07 RMSE: 0.09 | R2: 0.73 MAE: 0.06 RMSE: 0.08 | R2: 0.67 MAE: 0.06 RMSE: 0.08 | R2: 0.68 MAE: 0.06 RMSE: 0.09 |
| SampVal | R2: 0.77 MAE: 0.06 RMSE: 0.07 | R2: 0.80 MAE: 0.05 RMSE: 0.07 | R2: 0.75 MAE: 0.06 RMSE: 0.08 | R2: 0.78 MAE: 0.05 RMSE: 0.08 |
| FLVal | R2: 0.49 MAE: 0.08 RMSE: 0.10 | R2: 0.50 MAE: 0.08 RMSE: 0.10 | R2: 0.52 MAE: 0.08 RMSE: 0.10 | R2: 0.38 MAE: 0.09 RMSE: 0.11 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jurayj, D.; Bowers, R.; Fayne, J.V. Phenology and Plant Functional Type Link Optical Properties of Vegetation Canopies to Patterns of Vertical Vegetation Complexity. Remote Sens. 2024, 16, 2577. https://doi.org/10.3390/rs16142577
Jurayj D, Bowers R, Fayne JV. Phenology and Plant Functional Type Link Optical Properties of Vegetation Canopies to Patterns of Vertical Vegetation Complexity. Remote Sensing. 2024; 16(14):2577. https://doi.org/10.3390/rs16142577
Chicago/Turabian StyleJurayj, Duncan, Rebecca Bowers, and Jessica V. Fayne. 2024. "Phenology and Plant Functional Type Link Optical Properties of Vegetation Canopies to Patterns of Vertical Vegetation Complexity" Remote Sensing 16, no. 14: 2577. https://doi.org/10.3390/rs16142577
APA StyleJurayj, D., Bowers, R., & Fayne, J. V. (2024). Phenology and Plant Functional Type Link Optical Properties of Vegetation Canopies to Patterns of Vertical Vegetation Complexity. Remote Sensing, 16(14), 2577. https://doi.org/10.3390/rs16142577