
Mapping Malaria Vector Habitats in West Africa: Drone Imagery and Deep Learning Analysis for Targeted Vector Surveillance

 , , , , , , , , , and
, , , , , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
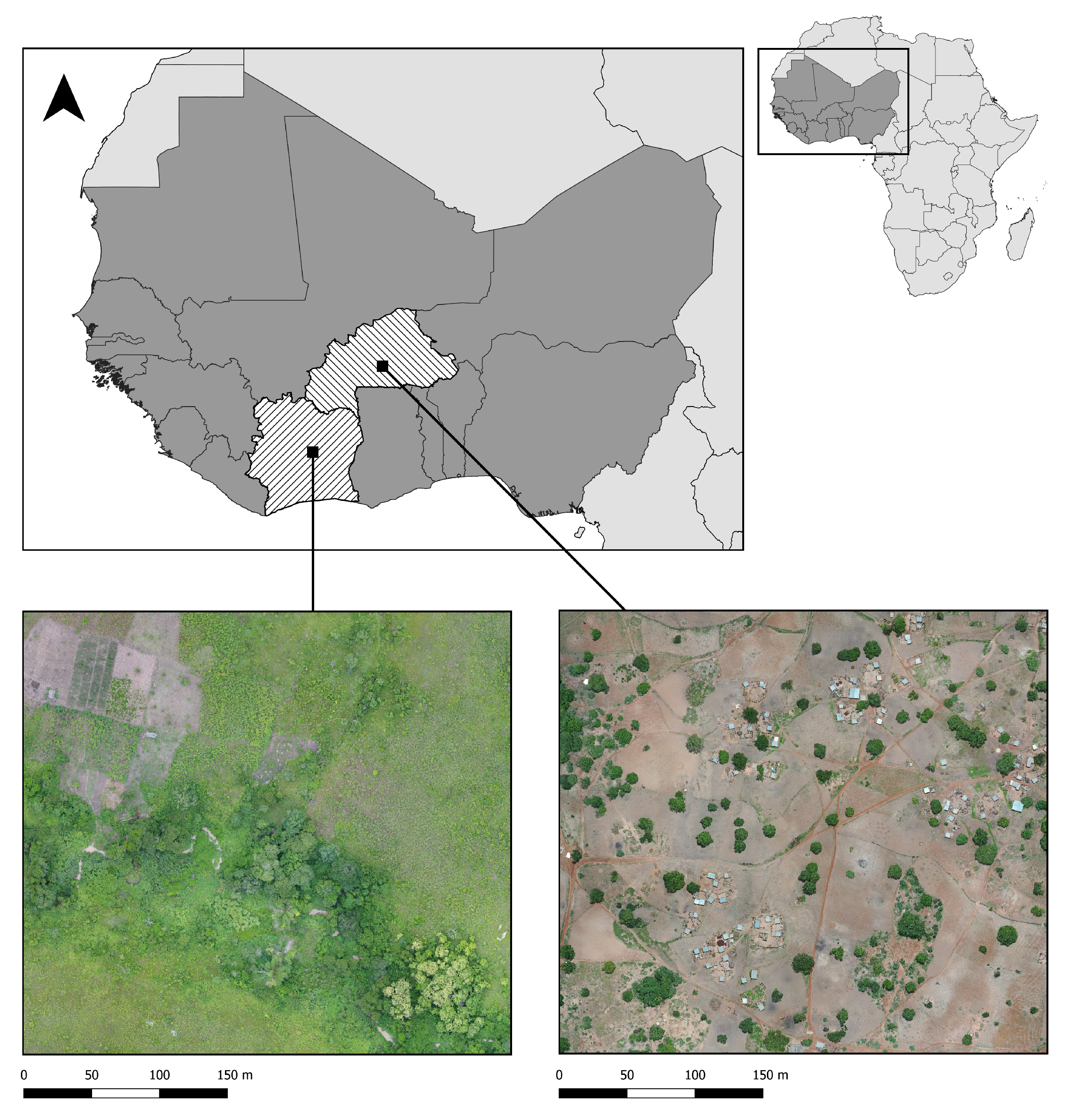
2.1. Drone Mapping
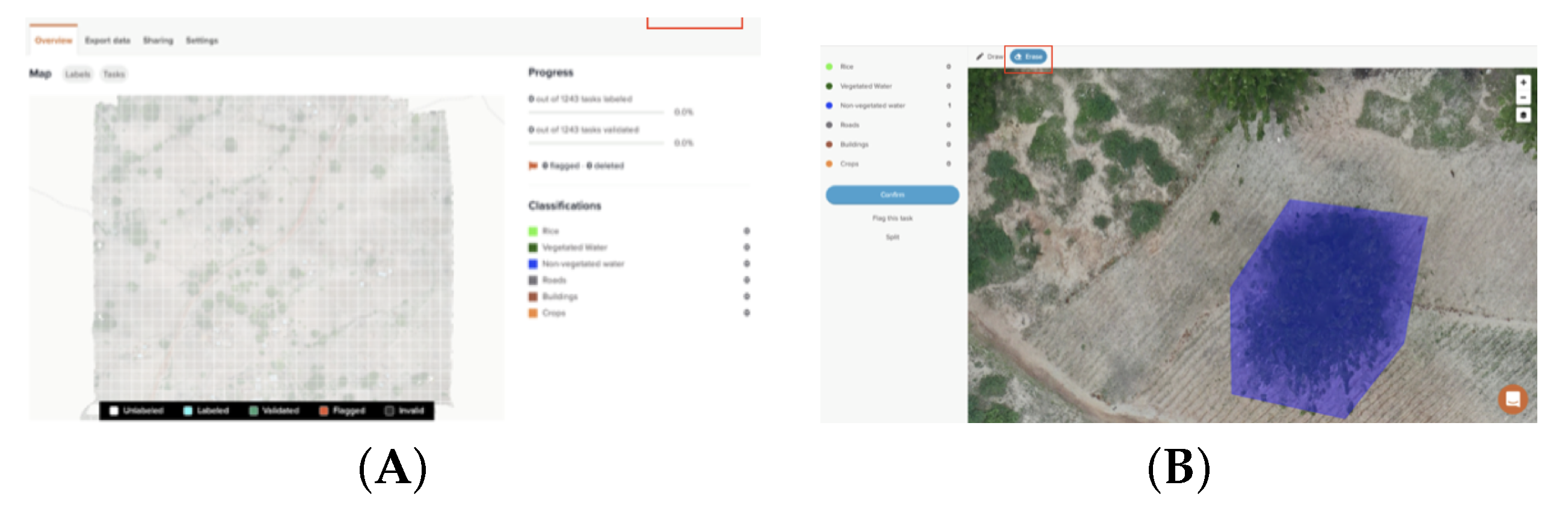
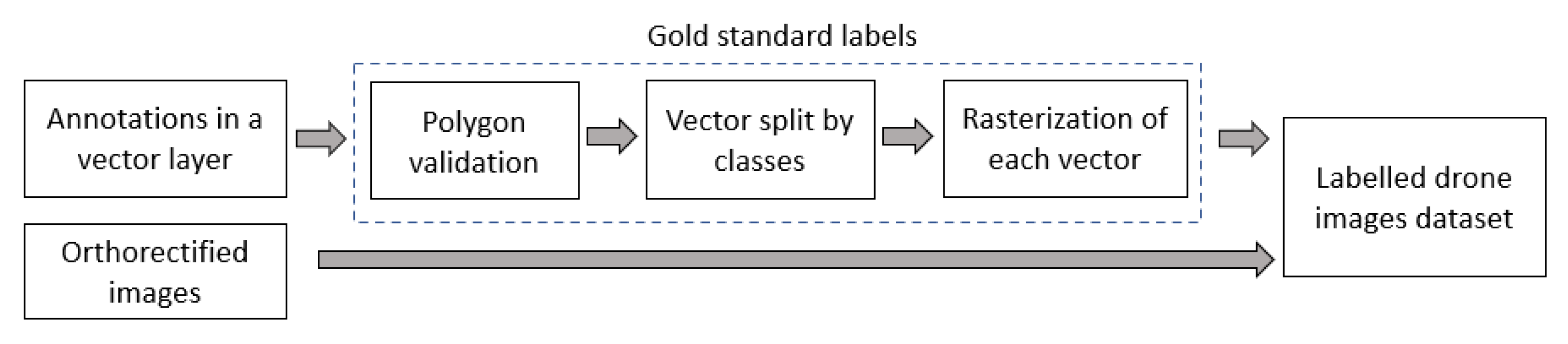
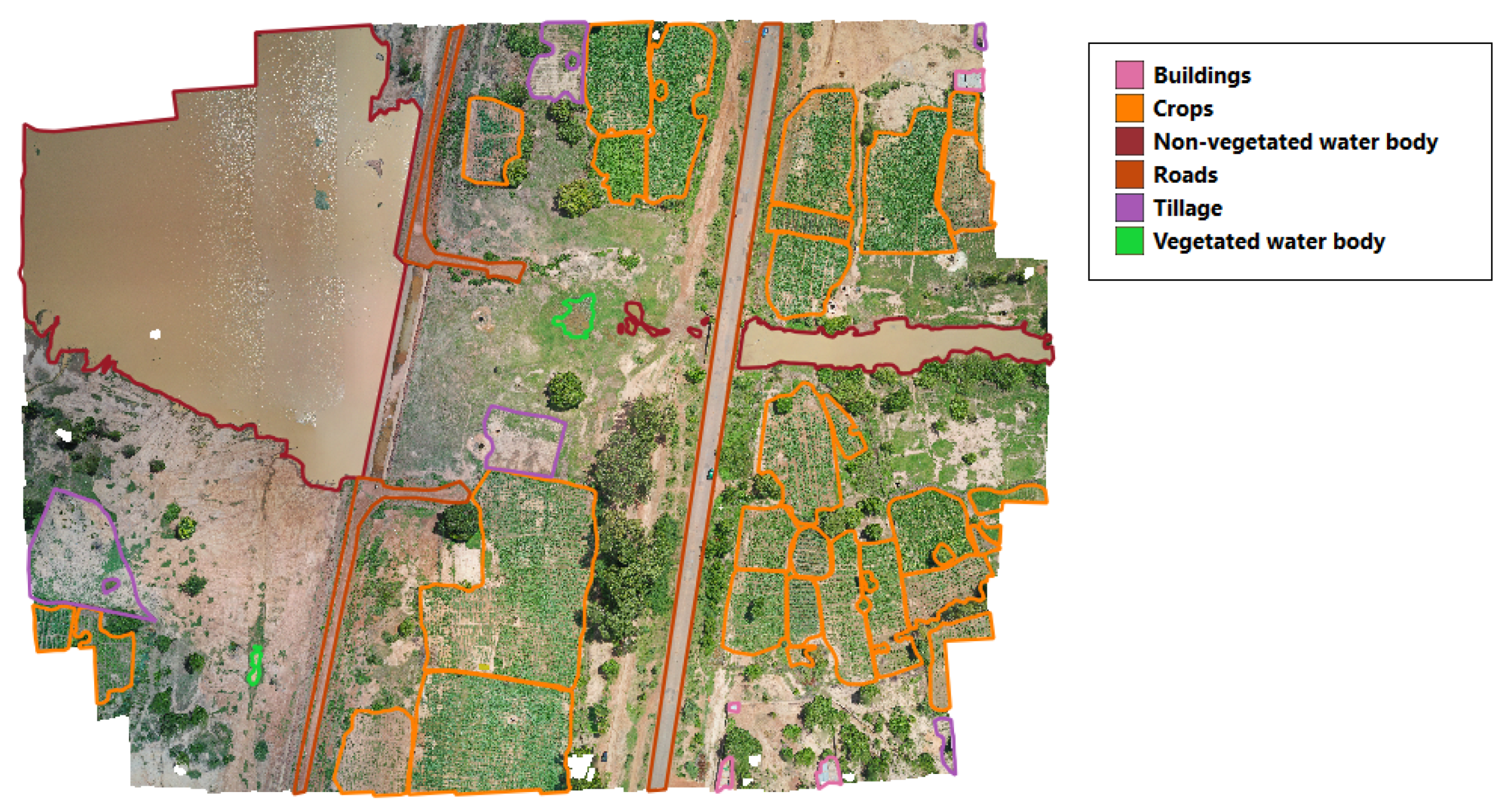
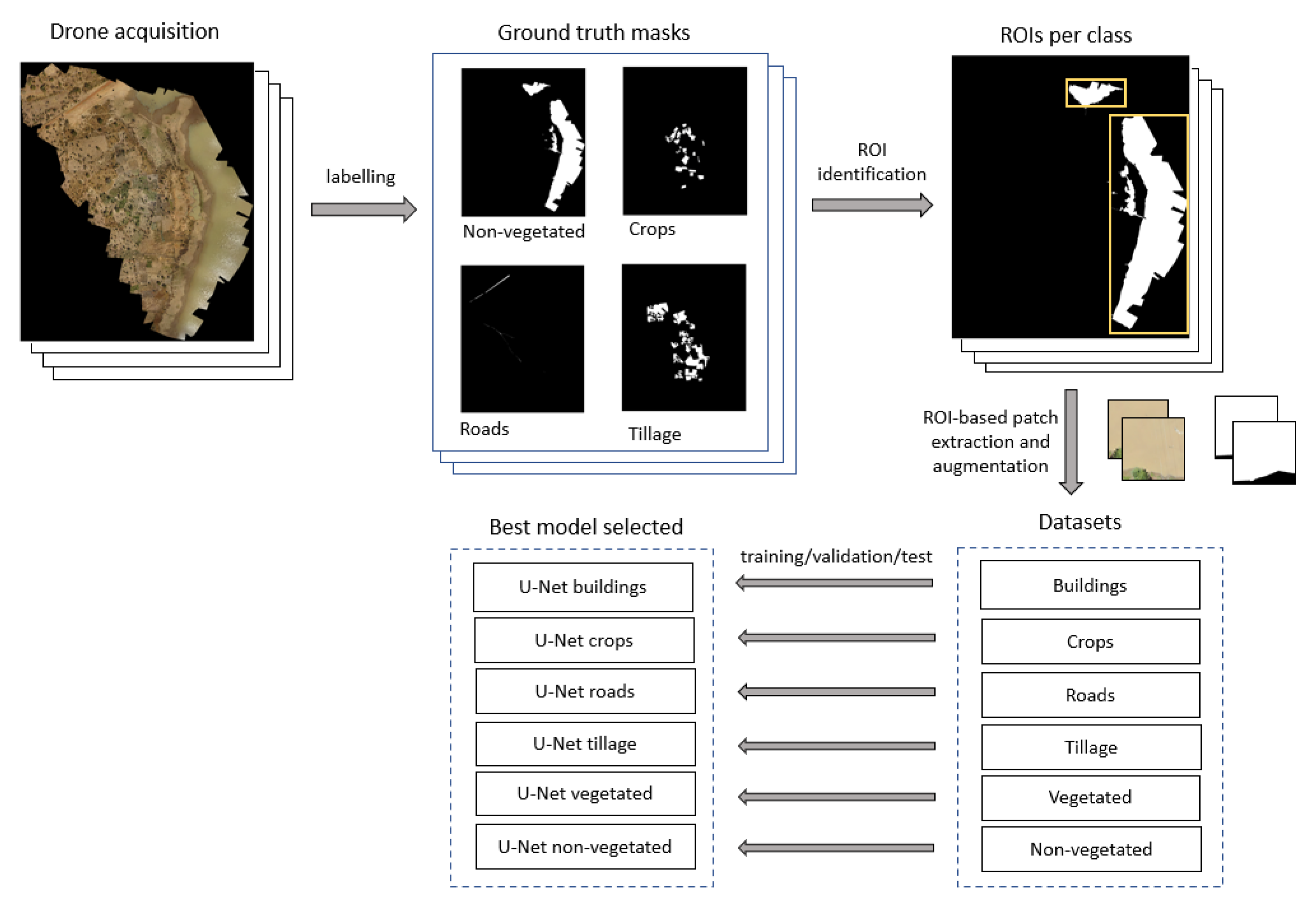
2.2. Image Labeling and Development of Labeled Dataset
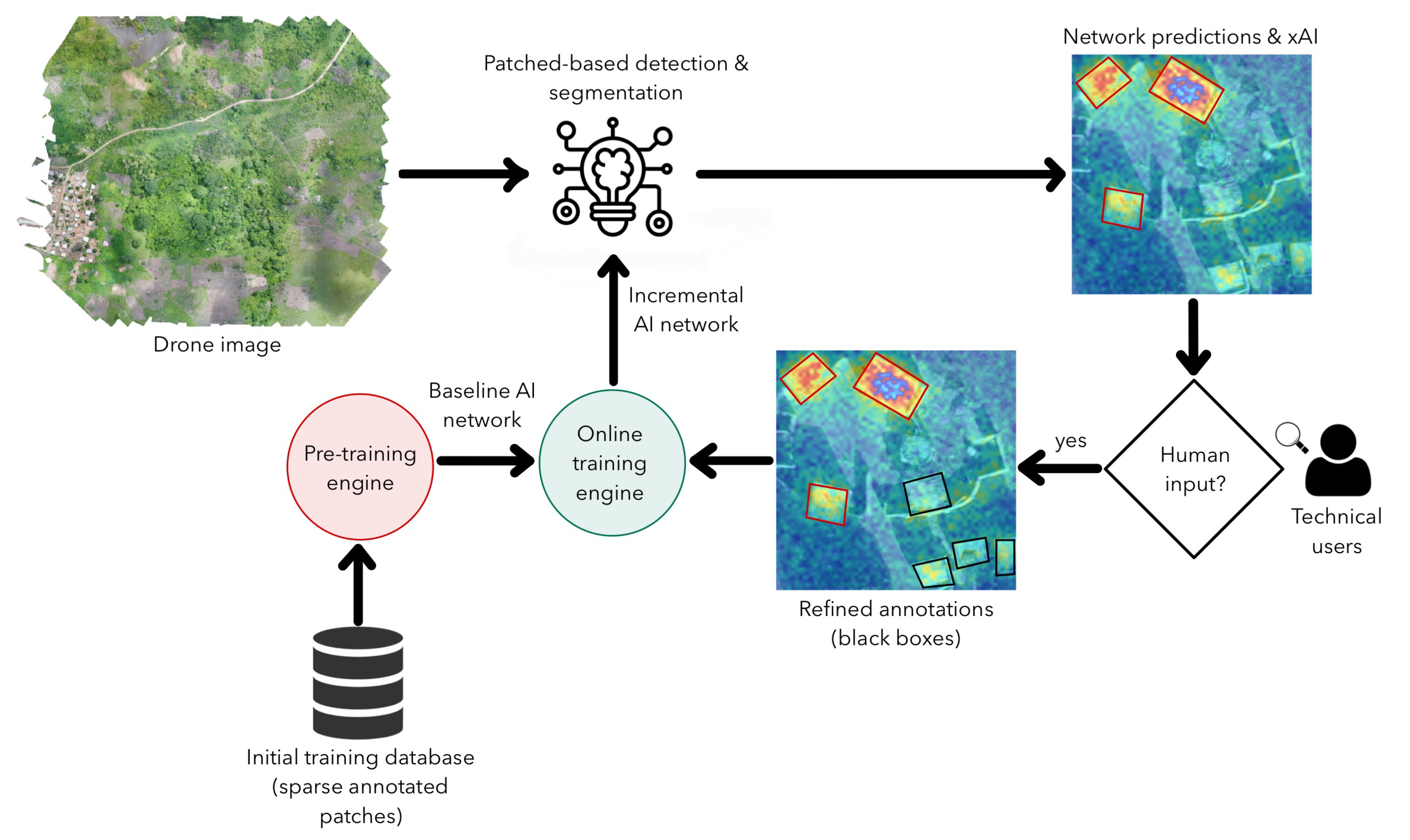
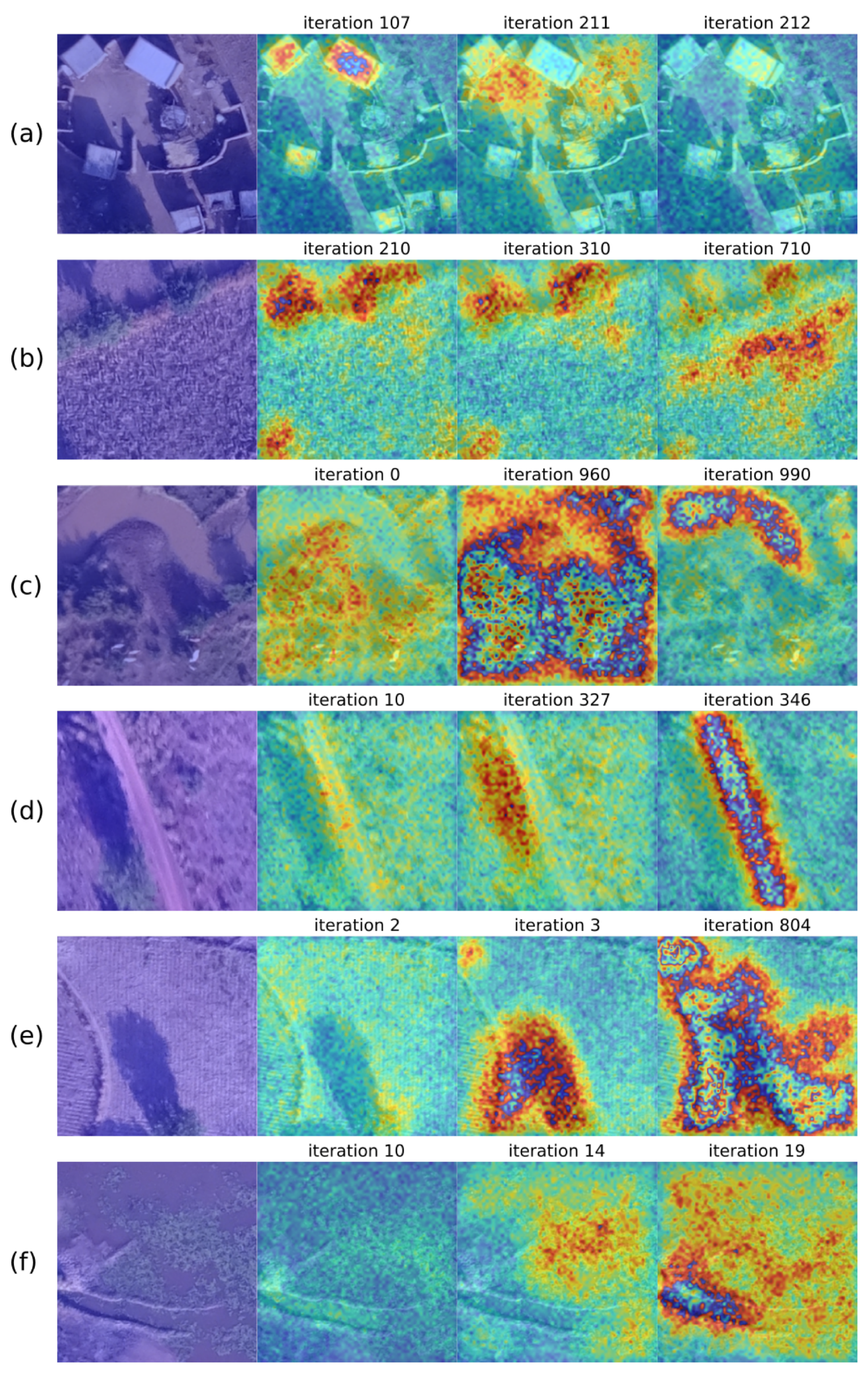
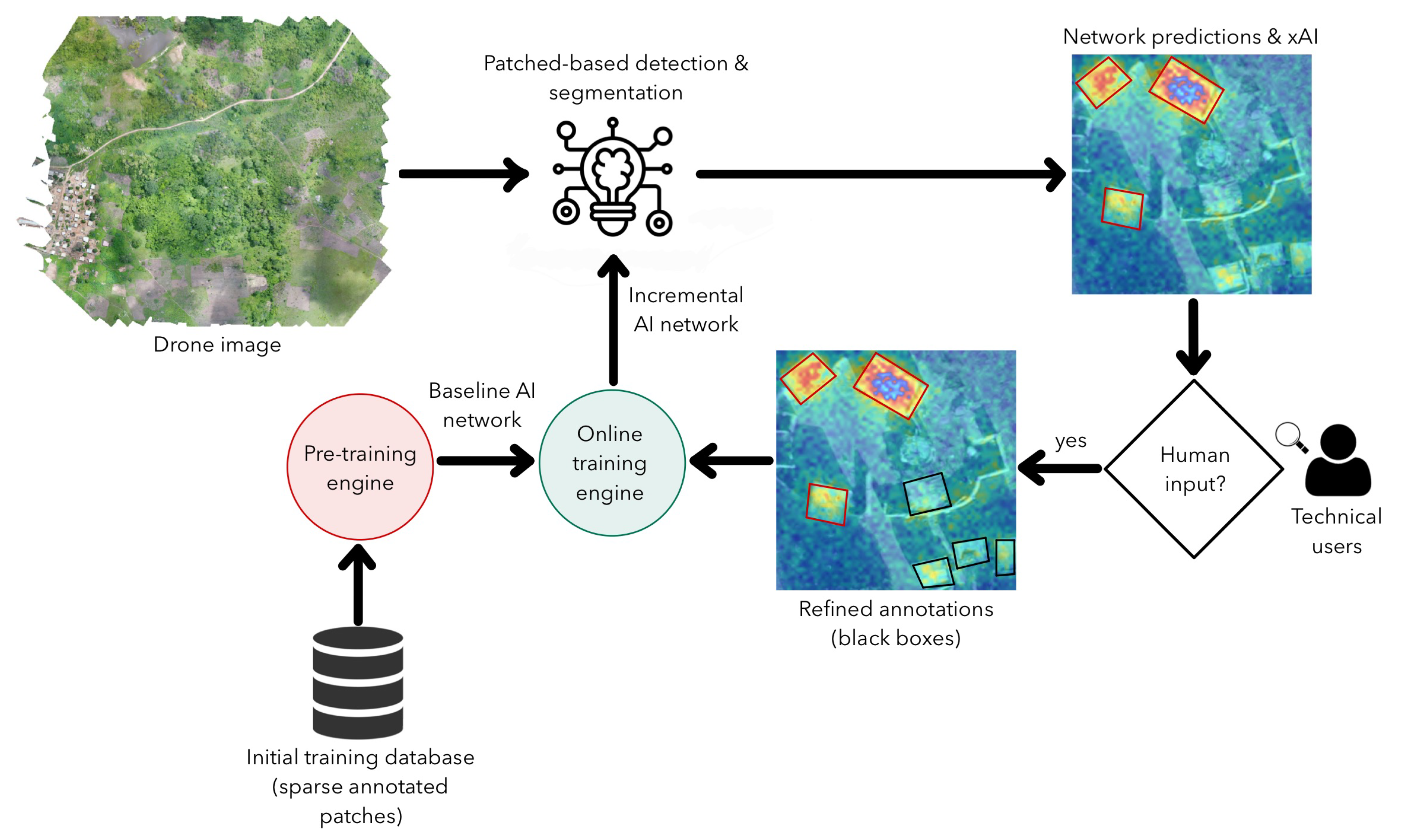
2.3. Algorithm Development
2.4. Evaluation Metrics
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| EO | Earth observation |
| UAV | unmanned aerial vehicle |
| CNN | convolutional neural network |
| DP | deep learning |
| TAD | technology-assisted digitizing |
| GLCM | gray-level co-occurrence matrix |
| RGB | red, green and blue |
| RMSE | root mean square error |
| MCSMA | multiple-criteria spectral mixture analysis |
| RNN | recurrent neural network |
| ReLU | rectified linear |
| CPU | central processing unit |
| RAM | random access memory |
References
- Patz, J.A.; Daszak, P.; Tabor, G.M.; Aguirre, A.A.; Pearl, M.; Epstein, J.; Wolfe, N.D.; Kilpatrick, A.M.; Foufopoulos, J.; Molyneux, D.; et al. Unhealthy landscapes: Policy recommendations on land use change and infectious disease emergence. Environ. Health Perspect. 2004, 112, 1092–1098. [Google Scholar] [CrossRef] [PubMed]
- Byrne, I.; Chan, K.; Manrique, E.; Lines, J.; Wolie, R.Z.; Trujillano, F.; Garay, G.J.; Del Prado Cortez, M.N.; Alatrista-Salas, H.; Sternberg, E.; et al. Technical Workflow Development for Integrating Drone Surveys and Entomological Sampling to Characterise Aquatic Larval Habitats of Anopheles funestus in Agricultural Landscapes in Côte d’Ivoire. J. Environ. Public Health 2021, 2021, 3220244. [Google Scholar] [CrossRef] [PubMed]
- Stanton, M.C.; Kalonde, P.; Zembere, K.; Spaans, R.H.; Jones, C.M. The application of drones for mosquito larval habitat identification in rural environments: A practical approach for malaria control? Malar. J. 2021, 20, 244. [Google Scholar] [CrossRef] [PubMed]
- Hardy, A.; Makame, M.; Cross, D.; Majambere, S.; Msellem, M. Using low-cost drones to map malaria vector habitats. Parasites Vectors 2017, 10, 29. [Google Scholar] [CrossRef]
- Hardy, A.; Ettritch, G.; Cross, D.E.; Bunting, P.; Liywalii, F.; Sakala, J.; Silumesii, A.; Singini, D.; Smith, M.; Willis, T.; et al. Automatic detection of open and vegetated water bodies using Sentinel 1 to map African malaria vector mosquito breeding habitats. Remote Sens. 2019, 11, 593. [Google Scholar] [CrossRef]
- Carrasco-Escobar, G.; Manrique, E.; Ruiz-Cabrejos, J.; Saavedra, M.; Alava, F.; Bickersmith, S.; Prussing, C.; Vinetz, J.M.; Conn, J.E.; Moreno, M.; et al. High-accuracy detection of malaria vector larval habitats using drone-based multispectral imagery. PLoS Neglected Trop. Dis. 2019, 13, e0007105. [Google Scholar] [CrossRef]
- Fornace, K.M.; Diaz, A.V.; Lines, J.; Drakeley, C.J. Achieving global malaria eradication in changing landscapes. Malar. J. 2021, 20, 69. [Google Scholar] [CrossRef]
- Lacey, L.A.; Lacey, C.M. The medical importance of riceland mosquitoes and their control using alternatives to chemical insecticides. J. Am. Mosq. Control. Assoc. Suppl. 1990, 2, 1–93. [Google Scholar]
- Tusting, L.S.; Thwing, J.; Sinclair, D.; Fillinger, U.; Gimnig, J.; Bonner, K.E.; Bottomley, C.; Lindsay, S.W. Mosquito larval source management for controlling malaria. Cochrane Database Syst. Rev. 2013, 2013, CD008923. [Google Scholar] [CrossRef]
- Ndiaye, A.; Niang, E.H.A.; Diène, A.N.; Nourdine, M.A.; Sarr, P.C.; Konaté, L.; Faye, O.; Gaye, O.; Sy, O. Mapping the breeding sites of Anopheles gambiae sl in areas of residual malaria transmission in central western Senegal. PLoS ONE 2020, 15, e0236607. [Google Scholar] [CrossRef]
- Kalluri, S.; Gilruth, P.; Rogers, D.; Szczur, M. Surveillance of arthropod vector-borne infectious diseases using remote sensing techniques: A review. PLoS Pathog. 2007, 3, e116. [Google Scholar] [CrossRef]
- Getzin, S.; Wiegand, K.; Schöning, I. Assessing biodiversity in forests using very high-resolution images and unmanned aerial vehicles. Methods Ecol. Evol. 2012, 3, 397–404. [Google Scholar] [CrossRef]
- Wimberly, M.C.; de Beurs, K.M.; Loboda, T.V.; Pan, W.K. Satellite observations and malaria: New opportunities for research and applications. Trends Parasitol. 2021, 37, 525–537. [Google Scholar] [CrossRef]
- Fornace, K.M.; Herman, L.S.; Abidin, T.R.; Chua, T.H.; Daim, S.; Lorenzo, P.J.; Grignard, L.; Nuin, N.A.; Ying, L.T.; Grigg, M.J.; et al. Exposure and infection to Plasmodium knowlesi in case study communities in Northern Sabah, Malaysia and Palawan, The Philippines. PLoS Neglected Trop. Dis. 2018, 12, e0006432. [Google Scholar] [CrossRef]
- Brock, P.M.; Fornace, K.M.; Grigg, M.J.; Anstey, N.M.; William, T.; Cox, J.; Drakeley, C.J.; Ferguson, H.M.; Kao, R.R. Predictive analysis across spatial scales links zoonotic malaria to deforestation. Proc. R. Soc. B 2019, 286, 20182351. [Google Scholar] [CrossRef]
- Byrne, I.; Aure, W.; Manin, B.O.; Vythilingam, I.; Ferguson, H.M.; Drakeley, C.J.; Chua, T.H.; Fornace, K.M. Environmental and spatial risk factors for the larval habitats of plasmodium knowlesi vectors in sabah, Malaysian borneo. Sci. Rep. 2021, 11, 11810. [Google Scholar] [CrossRef]
- Johnson, E.; Sharma, R.S.K.; Cuenca, P.R.; Byrne, I.; Salgado-Lynn, M.; Shahar, Z.S.; Lin, L.C.; Zulkifli, N.; Saidi, N.D.M.; Drakeley, C.; et al. Forest fragmentation drives zoonotic malaria prevalence in non-human primate hosts. bioRxiv 2022. [Google Scholar] [CrossRef]
- Gimnig, J.E.; Ombok, M.; Kamau, L.; Hawley, W.A. Characteristics of larval anopheline (Diptera: Culicidae) habitats in Western Kenya. J. Med. Entomol. 2001, 38, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Himeidan, Y.E.; Zhou, G.; Yakob, L.; Afrane, Y.; Munga, S.; Atieli, H.; El-Rayah, E.A.; Githeko, A.K.; Yan, G. Habitat stability and occurrences of malaria vector larvae in western Kenya highlands. Malar. J. 2009, 8, 234. [Google Scholar] [CrossRef]
- Kibret, S.; Wilson, G.G.; Ryder, D.; Tekie, H.; Petros, B. Malaria impact of large dams at different eco-epidemiological settings in Ethiopia. Trop. Med. Health 2017, 45, 4. [Google Scholar] [CrossRef]
- Nambunga, I.H.; Ngowo, H.S.; Mapua, S.A.; Hape, E.E.; Msugupakulya, B.J.; Msaky, D.S.; Mhumbira, N.T.; Mchwembo, K.R.; Tamayamali, G.Z.; Mlembe, S.V.; et al. Aquatic habitats of the malaria vector Anopheles funestus in rural south-eastern Tanzania. Malar. J. 2020, 19, 219. [Google Scholar] [CrossRef] [PubMed]
- Diakité, N.R.; Guindo-Coulibaly, N.; Adja, A.M.; Ouattara, M.; Coulibaly, J.T.; Utzinger, J.; N’Goran, E.K. Spatial and temporal variation of malaria entomological parameters at the onset of a hydro-agricultural development in central Côte d’Ivoire. Malar. J. 2015, 14, 340. [Google Scholar] [CrossRef] [PubMed]
- Zahouli, J.B.Z.; Koudou, B.G.; Müller, P.; Malone, D.; Tano, Y.; Utzinger, J. Effect of land-use changes on the abundance, distribution, and host-seeking behavior of Aedes arbovirus vectors in oil palm-dominated landscapes, southeastern Côte d’Ivoire. PLoS ONE 2017, 12, e0189082. [Google Scholar] [CrossRef] [PubMed]
- Dida, G.O.; Anyona, D.N.; Abuom, P.O.; Akoko, D.; Adoka, S.O.; Matano, A.S.; Owuor, P.O.; Ouma, C. Spatial distribution and habitat characterization of mosquito species during the dry season along the Mara River and its tributaries, in Kenya and Tanzania. Infect. Dis. Poverty 2018, 7, 2. [Google Scholar] [CrossRef] [PubMed]
- Mendis, C.; Jacobsen, J.L.; Gamage-Mendis, A.; Bule, E.; Dgedge, M.; Thompson, R.; Cuamba, N.; Barreto, J.; Begtrup, K.; Sinden, R.E.; et al. Anopheles arabiensis and An. funestus are equally important vectors of malaria in Matola coastal suburb of Maputo, southern Mozambique. Med. Vet. Entomol. 2000, 14, 171–180. [Google Scholar] [CrossRef]
- Omukunda, E.; Githeko, A.; Ndong’a, M.F.; Mushinzimana, E.; Yan, G. Effect of swamp cultivation on distribution of anopheline larval habitats in Western Kenya. J. Vector Borne Dis. 2012, 49, 61–71. [Google Scholar]
- Kweka, E.J.; Kamau, L.; Munga, S.; Lee, M.C.; Githeko, A.K.; Yan, G. A first report of Anopheles funestus sibling species in western Kenya highlands. Acta Trop. 2013, 128, 158–161. [Google Scholar] [CrossRef]
- Fornace, K.M.; Drakeley, C.J.; William, T.; Espino, F.; Cox, J. Mapping infectious disease landscapes: Unmanned aerial vehicles and epidemiology. Trends Parasitol. 2014, 30, 514–519. [Google Scholar] [CrossRef]
- Carrasco-Escobar, G.; Moreno, M.; Fornace, K.; Herrera-Varela, M.; Manrique, E.; Conn, J.E. The use of drones for mosquito surveillance and control. Parasites Vectors 2022, 15, 473. [Google Scholar] [CrossRef]
- Lu, D.; Weng, Q. A survey of image classification methods and techniques for improving classification performance. Int. J. Remote Sens. 2007, 28, 823–870. [Google Scholar] [CrossRef]
- Šiljeg, A.; Panđa, L.; Domazetović, F.; Marić, I.; Gašparović, M.; Borisov, M.; Milošević, R. Comparative Assessment of Pixel and Object-Based Approaches for Mapping of Olive Tree Crowns Based on UAV Multispectral Imagery. Remote Sens. 2022, 14, 757. [Google Scholar] [CrossRef]
- Hodgson, J.C.; Mott, R.; Baylis, S.M.; Pham, T.T.; Wotherspoon, S.; Kilpatrick, A.D.; Raja Segaran, R.; Reid, I.; Terauds, A.; Koh, L.P. Drones count wildlife more accurately and precisely than humans. Methods Ecol. Evol. 2018, 9, 1160–1167. [Google Scholar] [CrossRef]
- Gray, P.C.; Fleishman, A.B.; Klein, D.J.; McKown, M.W.; Bezy, V.S.; Lohmann, K.J.; Johnston, D.W. A convolutional neural network for detecting sea turtles in drone imagery. Methods Ecol. Evol. 2019, 10, 345–355. [Google Scholar] [CrossRef]
- Kattenborn, T.; Eichel, J.; Fassnacht, F.E. Convolutional Neural Networks enable efficient, accurate and fine-grained segmentation of plant species and communities from high-resolution UAV imagery. Sci. Rep. 2019, 9, 17656. [Google Scholar] [CrossRef]
- Liu, Z.Y.C.; Chamberlin, A.J.; Tallam, K.; Jones, I.J.; Lamore, L.L.; Bauer, J.; Bresciani, M.; Wolfe, C.M.; Casagrandi, R.; Mari, L.; et al. Deep Learning Segmentation of Satellite Imagery Identifies Aquatic Vegetation Associated with Snail Intermediate Hosts of Schistosomiasis in Senegal, Africa. Remote Sens. 2022, 14, 1345. [Google Scholar] [CrossRef]
- Hardy, A.; Oakes, G.; Hassan, J.; Yussuf, Y. Improved Use of Drone Imagery for Malaria Vector Control through Technology-Assisted Digitizing (TAD). Remote Sens. 2022, 14, 317. [Google Scholar] [CrossRef]
- Kwak, G.H.; Park, N.W. Impact of texture information on crop classification with machine learning and UAV images. Appl. Sci. 2019, 9, 643. [Google Scholar] [CrossRef]
- Hu, P.; Chapman, S.C.; Zheng, B. Coupling of machine learning methods to improve estimation of ground coverage from unmanned aerial vehicle (UAV) imagery for high-throughput phenotyping of crops. Funct. Plant Biol. 2021, 48, 766–779. [Google Scholar] [CrossRef]
- Komarkova, J.; Jech, J.; Sedlak, P. Comparison of Vegetation Spectral Indices Based on UAV Data: Land Cover Identification Near Small Water Bodies. In Proceedings of the 2020 15th Iberian Conference on Information Systems and Technologies (CISTI), Sevilla, Spain, 24–27 June 2020; pp. 1–4. [Google Scholar]
- Cao, S.; Xu, W.; Sanchez-Azofeif, A.; Tarawally, M. Mapping Urban Land Cover Using Multiple Criteria Spectral Mixture Analysis: A Case Study in Chengdu, China. In Proceedings of the IGARSS 2018–2018 IEEE International Geoscience and Remote Sensing Symposium, Valencia, Spain, 22–27 July 2018; pp. 2701–2704. [Google Scholar]
- Rustowicz, R.; Cheong, R.; Wang, L.; Ermon, S.; Burke, M.; Lobell, D. Semantic segmentation of crop type in Africa: A novel dataset and analysis of deep learning methods. In Proceedings of the IEEE/CVF Conference on Computer Vision and Pattern Recognition Workshops, Long Beach, CA, USA, 16–17 June 2019; pp. 75–82. [Google Scholar]
- Collins, K.A.; Ouedraogo, A.; Guelbeogo, W.M.; Awandu, S.S.; Stone, W.; Soulama, I.; Ouattara, M.S.; Nombre, A.; Diarra, A.; Bradley, J.; et al. Investigating the impact of enhanced community case management and monthly screening and treatment on the transmissibility of malaria infections in Burkina Faso: Study protocol for a cluster-randomised trial. BMJ Open 2019, 9, e030598. [Google Scholar] [CrossRef]
- Jimenez, G.; Kar, A.; Ounissi, M.; Ingrassia, L.; Boluda, S.; Delatour, B.; Stimmer, L.; Racoceanu, D. Visual Deep Learning-Based Explanation for Neuritic Plaques Segmentation in Alzheimer’s Disease Using Weakly Annotated Whole Slide Histopathological Images. In Lecture Notes in Computer Science, Proceedings of the Medical Image Computing and Computer Assisted Intervention (MICCAI), Singapore, 18–22 September 2022; Wang, L., Dou, Q., Fletcher, P.T., Speidel, S., Li, S., Eds.; Springer: Cham, Switzerland, 2022; Volume 13432, pp. 336–344. [Google Scholar] [CrossRef]
- Ronneberger, O.; Fischer, P.; Brox, T. U-net: Convolutional networks for biomedical image segmentation. In Proceedings of the International Conference on Medical image computing and computer-assisted intervention, Munich, Germany, 5–9 October 2015; Springer: Berlin/Heidelberg, Germany, 2015; pp. 234–241. [Google Scholar]
- Vaswani, A.; Shazeer, N.; Parmar, N.; Uszkoreit, J.; Jones, L.; Gomez, A.N.; Kaiser, Ł.; Polosukhin, I. Attention is all you need. In Proceedings of the Advances in Neural Information Processing Systems, Long Beach, CA, USA, 16–17 June 2017; Volume 30.
- Oktay, O.; Schlemper, J.; Folgoc, L.L.; Lee, M.; Heinrich, M.; Misawa, K.; Mori, K.; McDonagh, S.; Hammerla, N.Y.; Kainz, B.; et al. Attention u-net: Learning where to look for the pancreas. arXiv 2018, arXiv:1804.03999. [Google Scholar]
- Jimenez, G.; Kar, A.; Ounissi, M.; Stimmer, L.; Delatour, B.; Racoceanu, D. Interpretable Deep Learning in Computational Histopathology for refined identification of Alzheimer’s Disease biomarkers. In Proceedings of the Alzheimer’s & Dementia: Alzheimer’s Association International Conference (AAIC), San Diego, CA, USA, 2–4 August 2022; Wiley: Hoboken, NJ, USA, 2022. [Google Scholar]
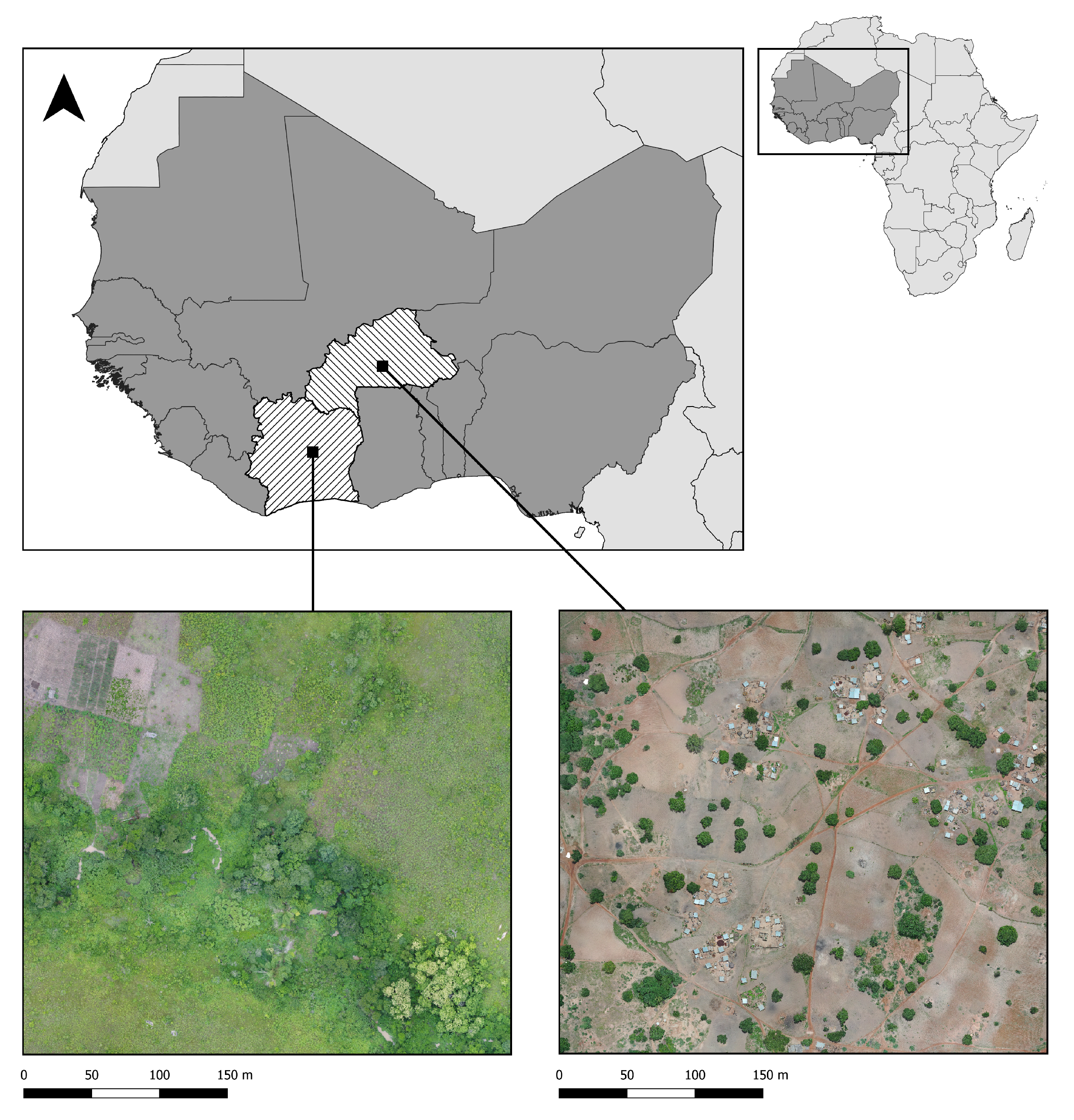


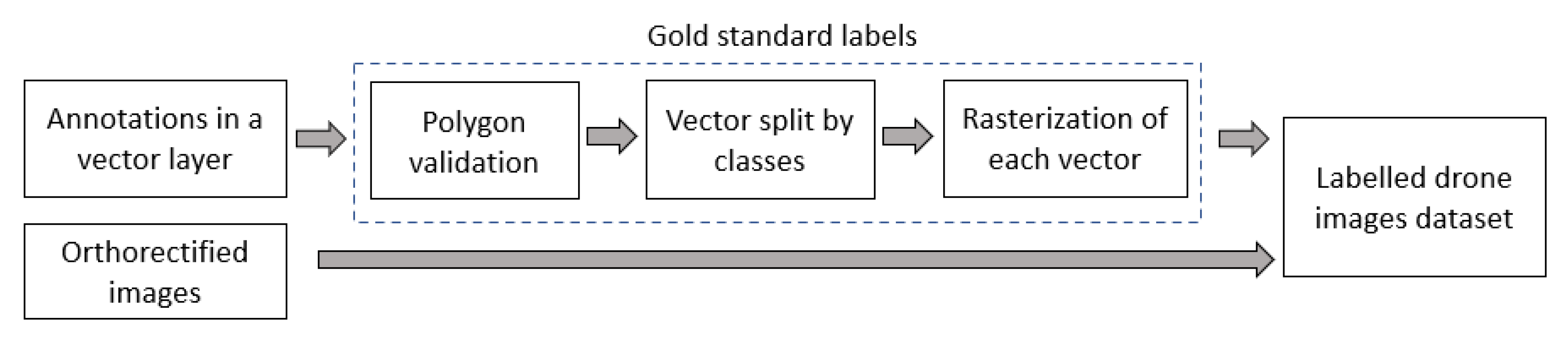

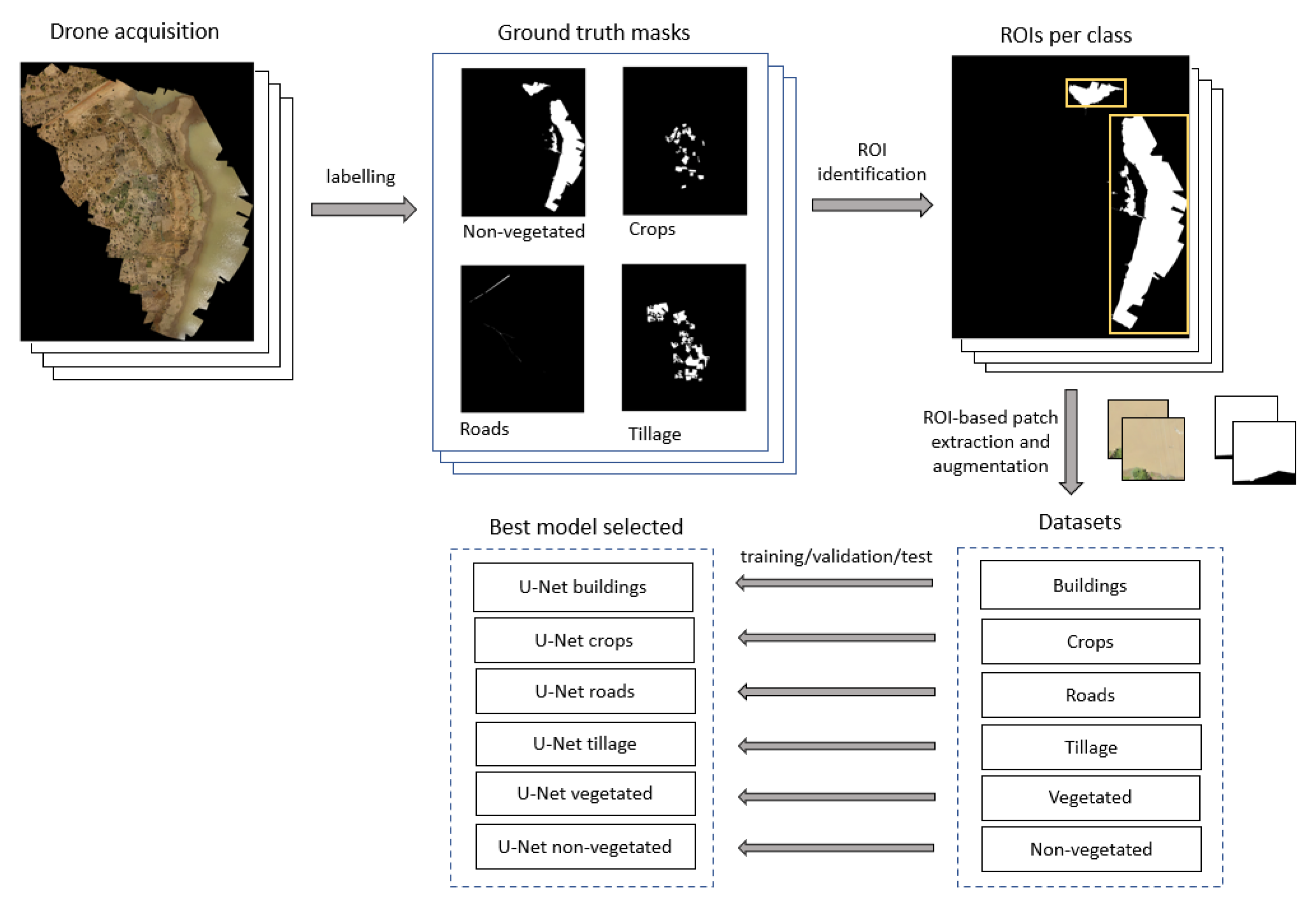
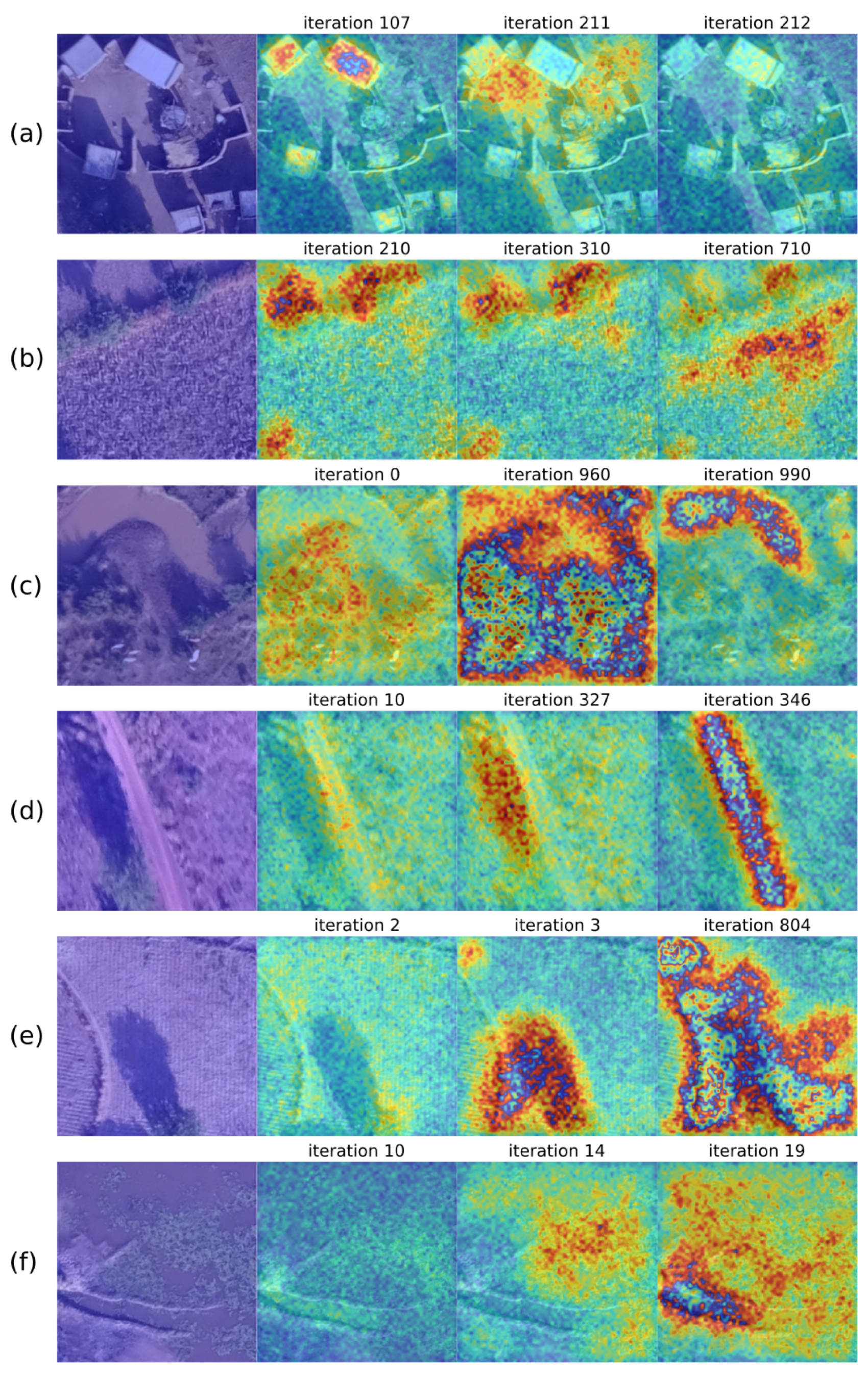


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Application | Imaging Source | Method | Result |
|---|---|---|---|---|
| Senegal River, West Africa [35] | Mapping snails’ aquatic habitats | Satellite: 8-band World View 2 for training UAV: used to assess labeling | Semantic segmentation using U-Net 8-band + GLCM features | Accuracy 4 classes Test: 82.7% 4 classes hold-out validation: 96.5% |
| Anbandegi, Korea [37] | Crop classification Kimchi cabbage | UAV: green, red, NIR bands | SVM and RF. Using GLCM features to reduce noise | Overall accuracy 4 classes: 98.72% |
| Queensland, Australia [38] | Ground coverage Wheat crops | UAV: RGB Real image set (RISs) Synthetic image set (SISs) | Two-step approach: per-pixel segmentation, sub-pixel segmentation using regression tree classifier | RMSE RISs: <6% SISs: <5% |
| Pardubice, Czech Republic [39] | Land cover identification near small water body | UAV: RGB | Comparing 8 different vegetation indexes | Visual comparison Best performance: NGRDI, GLI2, VARI |
| Chengdu, China [40] | Mapping vegetation, impervious surface and soil in urban environment | Satellite: Landsat-8 Operational Land Imager (OLI) | Applied multiple criteria spectral mixture analysis (MCSMA) with multi-step approach for spectral unmixing | RMSE Vegetation: 0.143 Soil: 0.170 Impervious: 0.151 |
| Ghana and South Sudan [41] | Semantic segmentation of crops in Africa | Satellite: Sentinel-1 (VV and VH), Sentinel-2 (10 bands) and Planet Scope (RGB + NIR) | Compared: 2D U-Net + CLSTM and 3D CNN using multi-temporal images | Accuracy South Sudan: 2D U-Net 88.7%, 3D 90% Ghana: 2D U-Net 65.7%, 3D 63.5% |
| Class | # Drone | # Train/Val | # Train/Val Patches | # Test | # Test Patches | ||
|---|---|---|---|---|---|---|---|
| Images | Images | 256 × 256 | 512 × 512 | Images | 256 × 256 | 512 × 512 | |
| Buildings | 48 | 36 | 7441 | 1324 | 12 | 4835 | 714 |
| Crops | 93 | 69 | 229,522 | 58,738 | 24 | 64,357 | 16,440 |
| Roads | 60 | 45 | 38,799 | 3908 | 15 | 14,330 | 2106 |
| Tillage | 42 | 31 | 86,988 | 22,817 | 11 | 39,025 | 10,264 |
| Non-vegetated | 37 | 27 | 9194 | 2232 | 10 | 79 | 1 |
| Vegetated | 20 | 15 | 4660 | 1107 | 5 | 564 | 125 |
| Burkina Faso | ||||||||
| Patch size | 256 × 256 | 512 × 512 | ||||||
| Class | patches | min (%) | mean (%) | max (%) | patches | min (%) | mean (%) | max (%) |
| Buildings | 6584 | 10.01 | 33.72 | 100 | 778 | 20.05 | 35.81 | 87.9 |
| Crops | 14,440 | 10.00 | 76.70 | 100 | 3662 | 20.00 | 71.23 | 100.0 |
| Roads | 42,810 | 10.00 | 29.51 | 100 | 4513 | 20.00 | 36.72 | 100.0 |
| Tillage | 121,952 | 10.00 | 75.49 | 100 | 32,089 | 20.00 | 70.35 | 100.0 |
| Non-vegetated | 7734 | 10.09 | 91.73 | 100 | 1900 | 20.08 | 89.51 | 100.0 |
| Vegetated | 251 | 10.17 | 65.89 | 100 | 58 | 20.49 | 61.03 | 100.0 |
| Côte d’Ivoire | ||||||||
| Patch size | 256 × 256 | 512 × 512 | ||||||
| Class | patches | min (%) | mean (%) | max (%) | patches | min (%) | mean (%) | max (%) |
| Buildings | 5692 | 10.0 | 46.5 | 100 | 1260 | 20.0 | 35.6 | 82 |
| Crops | 279,439 | 10.0 | 79.5 | 100 | 71,516 | 20.0 | 74.2 | 100 |
| Roads | 10,319 | 10.0 | 31.4 | 100 | 1501 | 20.0 | 26.7 | 96 |
| Tillage | 4061 | 10.0 | 69.4 | 100 | 992 | 20.1 | 62.3 | 100 |
| Non-vegetated | 1539 | 10.1 | 55.3 | 100 | 333 | 20.0 | 56.3 | 100 |
| Vegetated | 5646 | 10.0 | 62.3 | 100 | 1107 | 20.1 | 58.5 | 100 |
| Class | CV | FP | FN | TN | TP | Precision | Recall | Dice |
|---|---|---|---|---|---|---|---|---|
| Vegetated water body | 1 | 0.17 | 0.15 | 0.15 | 0.53 | 0.75 | 0.78 | 0.68 |
| 2 | 0.25 | 0.19 | 0.20 | 0.37 | 0.59 | 0.66 | 0.56 | |
| 3 | 0.21 | 0.12 | 0.22 | 0.45 | 0.68 | 0.78 | 0.65 | |
| Avg. | 0.21 | 0.15 | 0.19 | 0.45 | 0.67 | 0.74 | 0.63 | |
| Tillage | 1 | 0.10 | 0.03 | 0.13 | 0.74 | 0.88 | 0.96 | 0.88 |
| 2 | 0.10 | 0.09 | 0.15 | 0.66 | 0.87 | 0.88 | 0.82 | |
| 3 | 0.11 | 0.05 | 0.13 | 0.71 | 0.87 | 0.93 | 0.86 | |
| Avg. | 0.10 | 0.06 | 0.14 | 0.70 | 0.87 | 0.92 | 0.85 | |
| Roads | 1 | 0.09 | 0.07 | 0.63 | 0.21 | 0.70 | 0.74 | 0.70 |
| 2 | 0.06 | 0.06 | 0.71 | 0.17 | 0.73 | 0.75 | 0.71 | |
| 3 | 0.16 | 0.15 | 0.51 | 0.17 | 0.52 | 0.53 | 0.43 | |
| Avg. | 0.10 | 0.09 | 0.62 | 0.18 | 0.65 | 0.67 | 0.61 | |
| Non-vegetated water body | 1 | 0.02 | 0.54 | 0.06 | 0.38 | 0.95 | 0.41 | 0.50 |
| 2 | 0.21 | 0.26 | 0.26 | 0.27 | 0.57 | 0.51 | 0.48 | |
| 3 | 0.22 | 0.03 | 0.18 | 0.57 | 0.72 | 0.96 | 0.75 | |
| Avg. | 0.15 | 0.28 | 0.17 | 0.41 | 0.74 | 0.63 | 0.58 | |
| Crops | 1 | 0.10 | 0.06 | 0.10 | 0.74 | 0.87 | 0.93 | 0.86 |
| 2 | 0.13 | 0.04 | 0.09 | 0.75 | 0.85 | 0.95 | 0.86 | |
| 3 | 0.09 | 0.04 | 0.10 | 0.76 | 0.89 | 0.95 | 0.88 | |
| Avg. | 0.11 | 0.05 | 0.10 | 0.75 | 0.87 | 0.94 | 0.86 | |
| Building | 1 | 0.04 | 0.07 | 0.51 | 0.38 | 0.89 | 0.84 | 0.81 |
| 2 | 0.04 | 0.08 | 0.57 | 0.31 | 0.88 | 0.80 | 0.76 | |
| 3 | 0.04 | 0.03 | 0.54 | 0.38 | 0.90 | 0.92 | 0.87 | |
| Avg. | 0.04 | 0.06 | 0.54 | 0.35 | 0.89 | 0.85 | 0.81 |
| Class | CV | FP | FN | TN | TP | Precision | Recall | Dice |
|---|---|---|---|---|---|---|---|---|
| Vegetated water body | 1 | 0.27 | 0.08 | 0.15 | 0.49 | 0.64 | 0.85 | 0.67 |
| 2 | 0.28 | 0.11 | 0.17 | 0.44 | 0.61 | 0.81 | 0.64 | |
| 3 | 0.08 | 0.14 | 0.24 | 0.53 | 0.81 | 0.72 | 0.70 | |
| Avg. | 0.21 | 0.11 | 0.19 | 0.49 | 0.69 | 0.79 | 0.67 | |
| Tillage | 1 | 0.06 | 0.25 | 0.18 | 0.51 | 0.85 | 0.66 | 0.67 |
| 2 | 0.11 | 0.23 | 0.13 | 0.53 | 0.80 | 0.68 | 0.69 | |
| 3 | 0.03 | 0.55 | 0.27 | 0.15 | 0.77 | 0.20 | 0.27 | |
| Avg. | 0.07 | 0.34 | 0.19 | 0.40 | 0.81 | 0.51 | 0.54 | |
| Roads | 1 | 0.15 | 0.12 | 0.53 | 0.20 | 0.70 | 0.67 | 0.58 |
| 2 | 0.15 | 0.10 | 0.48 | 0.27 | 0.71 | 0.69 | 0.60 | |
| 3 | 0.35 | 0.06 | 0.40 | 0.19 | 0.51 | 0.77 | 0.46 | |
| Avg. | 0.22 | 0.09 | 0.47 | 0.22 | 0.64 | 0.71 | 0.55 | |
| Non-vegetated water body | 1 | 0.10 | 0.58 | 0.30 | 0.02 | 0.36 | 0.03 | 0.04 |
| 2 | 0.06 | 0.10 | 0.02 | 0.82 | 0.91 | 0.88 | 0.85 | |
| 3 | 0.20 | 0.05 | 0.26 | 0.49 | 0.71 | 0.90 | 0.72 | |
| Avg. | 0.12 | 0.24 | 0.19 | 0.44 | 0.66 | 0.60 | 0.54 | |
| Crops | 1 | 0.07 | 0.17 | 0.14 | 0.62 | 0.85 | 0.76 | 0.75 |
| 2 | 0.04 | 0.27 | 0.13 | 0.55 | 0.89 | 0.66 | 0.72 | |
| 3 | 0.16 | 0.04 | 0.05 | 0.75 | 0.81 | 0.94 | 0.84 | |
| Avg. | 0.09 | 0.16 | 0.11 | 0.64 | 0.85 | 0.79 | 0.77 | |
| Building | 1 | 0.03 | 0.06 | 0.56 | 0.36 | 0.91 | 0.84 | 0.85 |
| 2 | 0.03 | 0.07 | 0.53 | 0.37 | 0.91 | 0.82 | 0.83 | |
| 3 | 0.21 | 0.06 | 0.41 | 0.32 | 0.65 | 0.83 | 0.66 | |
| Avg. | 0.09 | 0.06 | 0.50 | 0.35 | 0.82 | 0.83 | 0.78 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Trujillano, F.; Jimenez Garay, G.; Alatrista-Salas, H.; Byrne, I.; Nunez-del-Prado, M.; Chan, K.; Manrique, E.; Johnson, E.; Apollinaire, N.; Kouame Kouakou, P.; et al. Mapping Malaria Vector Habitats in West Africa: Drone Imagery and Deep Learning Analysis for Targeted Vector Surveillance. Remote Sens. 2023, 15, 2775. https://doi.org/10.3390/rs15112775
Trujillano F, Jimenez Garay G, Alatrista-Salas H, Byrne I, Nunez-del-Prado M, Chan K, Manrique E, Johnson E, Apollinaire N, Kouame Kouakou P, et al. Mapping Malaria Vector Habitats in West Africa: Drone Imagery and Deep Learning Analysis for Targeted Vector Surveillance. Remote Sensing. 2023; 15(11):2775. https://doi.org/10.3390/rs15112775
Chicago/Turabian StyleTrujillano, Fedra, Gabriel Jimenez Garay, Hugo Alatrista-Salas, Isabel Byrne, Miguel Nunez-del-Prado, Kallista Chan, Edgar Manrique, Emilia Johnson, Nombre Apollinaire, Pierre Kouame Kouakou, and et al. 2023. "Mapping Malaria Vector Habitats in West Africa: Drone Imagery and Deep Learning Analysis for Targeted Vector Surveillance" Remote Sensing 15, no. 11: 2775. https://doi.org/10.3390/rs15112775
APA StyleTrujillano, F., Jimenez Garay, G., Alatrista-Salas, H., Byrne, I., Nunez-del-Prado, M., Chan, K., Manrique, E., Johnson, E., Apollinaire, N., Kouame Kouakou, P., Oumbouke, W. A., Tiono, A. B., Guelbeogo, M. W., Lines, J., Carrasco-Escobar, G., & Fornace, K. (2023). Mapping Malaria Vector Habitats in West Africa: Drone Imagery and Deep Learning Analysis for Targeted Vector Surveillance. Remote Sensing, 15(11), 2775. https://doi.org/10.3390/rs15112775