
Identification and Counting of European Souslik Burrows from UAV Images by Pixel-Based Image Analysis and Random Forest Classification: A Simple, Semi-Automated, yet Accurate Method for Estimating Population Size

 ,
,  ,
,  ,
, 
Abstract
:
1. Introduction
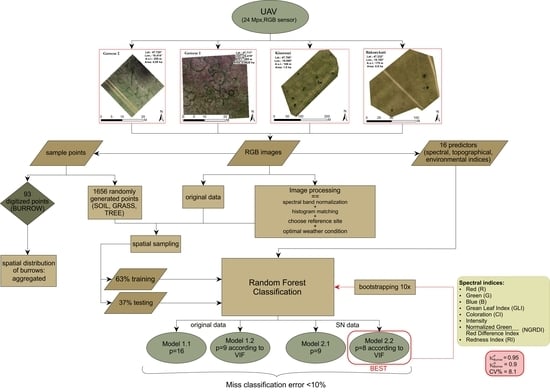
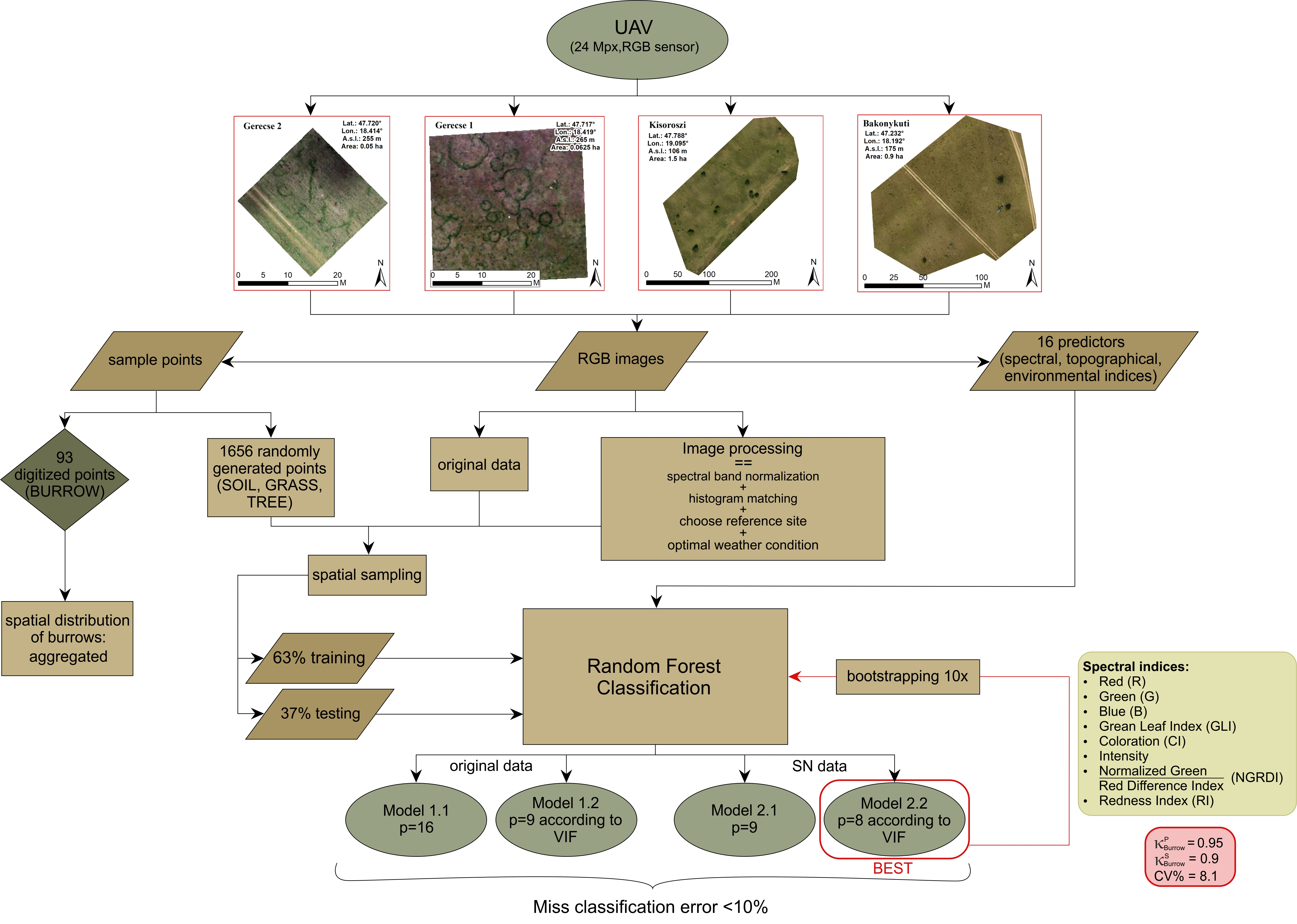
2. Materials and Methods
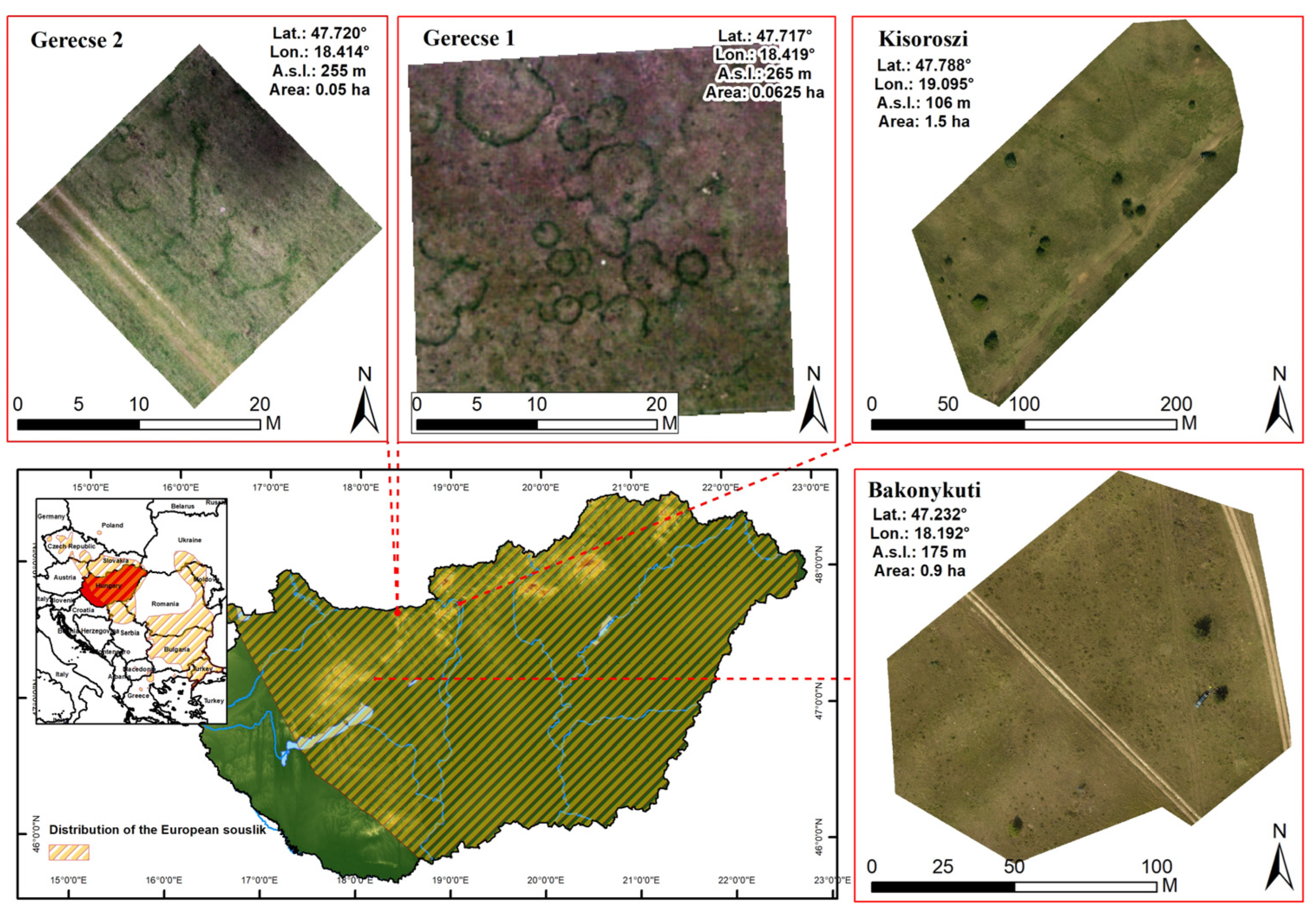
2.1. Field Survey
2.2. Statistical Analyses
2.3. UAV Imagery and Spectral Data Pre-Processing
2.4. Random Forest Classification of Pixels
2.5. Evaluation of RF Classification
2.6. Model Stability
2.7. Spatial Distribution of Burrows (Mounds or/and BOs)
3. Results
3.1. Evaluation of RF Classification
3.2. Model Stability
3.3. Spatial Pattern of BOs
4. Discussion
Technical Issues
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Feldhamer, G.; Drickamer, L.; Vessey, S.; Merritt, J. Mammalogy: Adaptation, Diversity, Ecology; 3rd ed.; JHU Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Gomes Rodrigues, H.; Šumbera, R.; Hautier, L. Life in Burrows Channelled the Morphological Evolution of the Skull in Rodents: The Case of African Mole-Rats (Bathyergidae, Rodentia). J. Mamm. Evol. 2016, 23, 175–189. [Google Scholar] [CrossRef] [Green Version]
- Meadows, P.S.; Meadows, A. The Environmental Impact of Burrowing Animals and Animal Burrows. In Proceedings of the Symposium, London, UK, 3–4 May 1990, 1st ed.; Published for the Zoological Society of London by Clarendon Press: London, UK, 1991; ISBN 0198546807. [Google Scholar]
- Haussmann, N.S. Soil Movement by Burrowing Mammals: A Review Comparing Excavation Size and Rate to Body Mass of Excavators. Prog. Phys. Geogr. 2017, 41, 29–45. [Google Scholar] [CrossRef] [Green Version]
- Hansell, M.H. The Ecological Impact of Animal Nests and Burrows. Funct. Ecol. 1993, 7, 5. [Google Scholar] [CrossRef]
- Whitford, W.G.; Kay, F.R. Biopedturbation by Mammals in Deserts: A Review. J. Arid Environ. 1999, 41, 203–230. [Google Scholar] [CrossRef]
- Sandifer, P.A.; Sutton-Grier, A.E.; Ward, B.P. Exploring Connections among Nature, Biodiversity, Ecosystem Services, and Human Health and Well-Being: Opportunities to Enhance Health and Biodiversity Conservation. Ecosyst. Serv. 2015, 12, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Brevik, E.C.; Slaughter, L.; Singh, B.R.; Steffan, J.J.; Collier, D.; Barnhart, P.; Pereira, P. Soil and Human Health: Current Status and Future Needs. Air Soil Water Res. 2020, 13, 1178622120934441. [Google Scholar] [CrossRef]
- Davidson, A.D.; Lightfoot, D.C. Interactive Effects of Keystone Rodents on the Structure of Desert Grassland Arthropod Communities. Ecography 2007, 30, 515–525. [Google Scholar] [CrossRef]
- Ewacha, M.V.A.; Kaapehi, C.; Waterman, J.M.; Roth, J.D. Cape Ground Squirrels as Ecosystem Engineers: Modifying Habitat for Plants, Small Mammals and Beetles in Namib Desert Grasslands. Afr. J. Ecol. 2016, 54, 68–75. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; Montagne, J.P.; Lenihan, C.M.; Wisinski, C.L.; Nordstrom, L.A.; Shier, D.M. Capturing Pests and Releasing Ecosystem Engineers: Translocation of Common but Diminished Species to Re-Establish Ecological Roles. Anim. Conserv. 2019, 22, 600–610. [Google Scholar] [CrossRef] [Green Version]
- Lindtner, P.; Gömöryová, E.; Gömöry, D.; Stašiov, S.; Kubovčík, V. Development of Physico-Chemical and Biological Soil Properties on the European Ground Squirrel Mounds. Geoderma 2019, 339, 85–93. [Google Scholar] [CrossRef]
- Butler, D.R. Zoogeomorphology: Animals as Geomorphic Agents; Cambridge University Press: Cambridge, UK, 1995. [Google Scholar] [CrossRef]
- Johnson, D.L. Biomantle Evolution and the Redistribution of Earth Materials and Artifacts. Soil Sci. 1990, 149, 84–102. [Google Scholar] [CrossRef]
- Hegyeli, Z. Spermophilus citellus. The IUCN Red List of Threatened Species 2020: E.T20472A91282380. Available online: https://dx.doi.org/10.2305/IUCN.UK.2020-2.RLTS.T20472A91282380.en (accessed on 4 March 2022).
- Ramos-Lara, N.; Koprowski, J.L.; Kryštufek, B.; Hoffmann, I.E. Spermophilus Citellus (Rodentia: Sciuridae). Mamm. Species 2014, 913, 71–87. [Google Scholar] [CrossRef] [Green Version]
- Erdos, L.; Tölgyesi, C.; Horzse, M.; Tolnay, D.; Hurton, Á.; Schulcz, N.; Körmöczi, L.; Lengyel, A.; Bátori, Z. Habitat Complexity of the Pannonian Forest-Steppe Zone and Its Nature Conservation Implications. Ecol. Complex. 2014, 17, 107–118. [Google Scholar] [CrossRef]
- Győri-Koósz, B.; Faragó, S. Az Ürge (Spermophilus Citellus) Tápláléknövényei, Mint Potenciális Elterjedési Tényezők, Ökológiai Értékelésük Alapján. Magy. Apróvad Közlemények 2017, 13, 161–175. [Google Scholar] [CrossRef] [Green Version]
- Šefferová, S.; Janák, M.; Vajda, Z. MANAGEMENT of Natura 2000 Habitats: Pannonic Sand Steppes; European Commission: Brussels, Belgium, 2008. [Google Scholar]
- Ruzic, A. Citellus Citellus (Linaeus, 1766)—Der Oder Das Europäische Ziesel. In Handbuch der Säugetiere Europas, Bd. 1, Nagetiere I; Akad Verlagsgesellschaft: Wiesbaden, Germany, 1978; pp. 123–144. [Google Scholar]
- Gedeon, C.I.; Boross, G.; Németh, A.; Altbäcker, V. Release Site Manipulation to Favour European Ground Squirrel Spermophilus Citellus Translocations: Translocation and Habitat Manipulation. Wildl. Biol. 2012, 18, 97–104. [Google Scholar] [CrossRef] [Green Version]
- Mcdonald, B.L.L.; Stanley, T.R.; Otis, D.L.; Biggins, D.E.; Stevens, D.; Koprowski, J.L.; Ballard, W. Recommended Methods for Range-Wide Monitoring of Prairie Dogs in the United States; US Department of the Interior, US Geological Survey: Richmond, VA, USA, 2011; p. 36. [Google Scholar]
- Willcox, D.; Nash, H.C.; Trageser, S.; Kim, H.J.; Hywood, L.; Connelly, E.; Ichu Ichu, G.; Kambale Nyumu, J.; Mousset Moumbolou, C.L.; Ingram, D.J.; et al. Evaluating Methods for Detecting and Monitoring Pangolin (Pholidata: Manidae) Populations. Glob. Ecol. Conserv. 2019, 17, e00539. [Google Scholar] [CrossRef]
- Biggins, D.E.; Miller, B.J.; Hanebury, L.R.; Oakleaf, B.; Farmer, A.H.; Crete, R.; Dood, A. A Technique for Evaluating Black-Footed Ferret Habitat. In Management of Prairie Dog Complexes for the Reintroduction of the Black-Footed Ferret; US Fish and Wildlife Service Biological Report: Washington, DC, USA, 1993; Volume 13, pp. 73–88. [Google Scholar]
- Harper, S.J.; Batzli, G.O. Effects of Predators on Structure of the Burrows of Voles. J. Mammal. 1996, 77, 1114–1121. [Google Scholar] [CrossRef] [Green Version]
- Hubbs, A.H.; Karels, T.; Boonstra, R. Indices of Population Size for Burrowing Mammals. J. Wildl. Manage. 2000, 64, 296. [Google Scholar] [CrossRef] [Green Version]
- Janák, M.; Marhoul, P.; Mateju, J. Action Plan for the Conservation of the European Ground Squirrel Spermophilus Citellus in the European Union List of Contributors; European Commission: Brussels, Belgium, 2013. [Google Scholar]
- Csorba, G.; Pecsenye, K. Nemzeti Biodiverzitás-Monitorozó Rendszer X. Eml”osök És a Genetikai Sokféleség Monitorozása. Available online: https://scholar.google.com/scholar?hl=en&as_sdt=0%2C5&q=A+Nemzeti+Biodiverzitás-monitorozó+Rendszer+X.+Emlősök+és+a+genetikai+sokféleség+monitorozása&btnG= (accessed on 17 July 2021).
- Váczi, O.; Varga, I.; Bakó, B. A Nemzeti Biodiverzitás-Monitorozó Rendszer Eredményei II—Gerinces Állatok; Körös-Maros Nemzeti Park Igazgatóság: Szarvas, Hungary, 2019. [Google Scholar]
- Gedeon, C.I.; Hoffmann, I.E.; Váczi, O.; Knauer, F.; Ben Slimen, H.; Stefanović, M.; Lehoczky, É.; Laborczi, A.; Suchentrunk, F. The Role of Landscape History in Determining Allelic Richness of European Ground Squirrels (Spermophilus Citellus) in Central Europe. Hystrix 2017, 28, 240–246. [Google Scholar] [CrossRef]
- Cepáková, E.; Hulová, S. Current Distribution of the European Souslik (Spermophilus Citellus) in the Czech Republic. Lynx 2002, 33, 89–103. [Google Scholar]
- Stoeva, E.; Ivanov, I.; Stoev, I.; Yankov, L.; Mechev, A.; Koshev, Y. Successful Reinforcement of the European Souslik by Green Balkans NGO in “Sinite Kamani” Nature Park. In Annuaire de l’Université de Sofia “St. Kliment Ohridski” Faculte de Biologie; University Press: Sofia, Bulgaria, 2016; pp. 153–165. [Google Scholar]
- Hoogland, J. (Ed.) Conservation of the Black-Tailed Prairie Dog: Saving North America’s Western Grasslands; Island Press: Chicago, IL, USA, 2013; Volume 88, ISBN 1559634979. [Google Scholar]
- Hut, R.A.; Scharff, A. Endoscopie Observations on Tunnel Blocking Behaviour in the European Ground Squirrel (Spermophilus Citellus). Z. Fur Saugetierkd. 1998, 63, 377–380. [Google Scholar]
- Katona, K.; Váczi, O.; Altbäcker, V. Topographic Distribution and Daily Activity of the European Ground Squirrel Population in Bugacpuszta, Hungary. Acta Theriol. 2002, 47, 45–54. [Google Scholar] [CrossRef]
- Johson, D.L.; Burnham, J.L.H. (Eds.) Introduction: Overview of concepts, definitions, and principles of soil mound studies. In Mima Mounds: The Case for Polygenesis and Bioturbation: Geological Society of America Special Paper; 2012; Volume 490, pp. 1–99. [Google Scholar] [CrossRef]
- Šumbera, R.; Mazoch, V.; Patzenhauerová, H.; Lövy, M.; Šklíba, J.; Bryja, J.; Burda, H. Burrow Architecture, Family Composition and Habitat Characteristics of the Largest Social African Mole-Rat: The Giant Mole-Rat Constructs Really Giant Burrow Systems. Acta Theriol. 2012, 57, 121–130. [Google Scholar] [CrossRef]
- Velasco, M. A Quickbird’s Eye View on Marmots; International Institute for Geo-Information Science and Earth Observation: Enschede, The Netherlands, 2009. [Google Scholar]
- Pettorelli, N.; Laurance, W.F.; O’Brien, T.G.; Wegmann, M.; Nagendra, H.; Turner, W. Satellite Remote Sensing for Applied Ecologists: Opportunities and Challenges. J. Appl. Ecol. 2014, 51, 839–848. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, F.; Kung, H.; Johnson, V.C. New Methods for Improving the Remote Sensing Estimation of Soil Organic Matter Content (SOMC) in the Ebinur Lake Wetland National Nature Reserve (ELWNNR) in Northwest China. Remote Sens. Environ. 2018, 218, 104–118. [Google Scholar] [CrossRef]
- Swinbourne, M.J.; Taggart, D.A.; Swinbourne, A.M.; Lewis, M.; Ostendorf, B. Using Satellite Imagery to Assess the Distribution and Abundance of Southern Hairy-Nosed Wombats (Lasiorhinus Latifrons). Remote Sens. Environ. 2018, 211, 196–203. [Google Scholar] [CrossRef]
- Wilschut, L.I.; Addink, E.A.; Heesterbeek, J.A.P.; Dubyanskiy, V.M.; Davis, S.A.; Laudisoit, A.; Begon, M.; Burdelov, L.A.; Atshabar, B.B.; de Jong, S.M. Mapping the Distribution of the Main Host for Plague in a Complex Landscape in Kazakhstan: An Object-Based Approach Using SPOT-5 XS, Landsat 7 ETM+, SRTM and Multiple Random Forests. Int. J. Appl. Earth Obs. Geoinf. 2013, 23, 81–94. [Google Scholar] [CrossRef] [Green Version]
- Wilschut, L.I.; Heesterbeek, J.A.P.; Begon, M.; de Jong, S.M.; Ageyev, V.; Laudisoit, A.; Addink, E.A. Detecting Plague-Host Abundance from Space: Using a Spectral Vegetation Index to Identify Occupancy of Great Gerbil Burrows. Int. J. Appl. Earth Obs. Geoinf. 2018, 64, 249–255. [Google Scholar] [CrossRef]
- Boyaci, D.; Erdoğan, M.; Yildiz, F. Pixel-versus Object-Based Classification of Forest and Agricultural Areas from Multiresolution Satellite Images. Turk. J. Electr. Eng. Comput. Sci. 2017, 25, 365–375. [Google Scholar] [CrossRef]
- Vlachopoulos, O.; Leblon, B.; Wang, J.; Haddadi, A.; LaRocque, A.; Patterson, G. Delineation of Crop Field Areas and Boundaries from UAS Imagery Using PBIA and GEOBIA with Random Forest Classification. Remote Sens. 2020, 12, 2640. [Google Scholar] [CrossRef]
- Vlachopoulos, O.; Leblon, B.; Wang, J.; Haddadi, A.; LaRocque, A.; Patterson, G. Delineation of Bare Soil Field Areas from Unmanned Aircraft System Imagery with the Mean Shift Unsupervised Clustering and the Random Forest Supervised Classification. Can. J. Remote Sens. 2020, 46, 489–500. [Google Scholar] [CrossRef]
- Berhane, T.M.; Lane, C.R.; Wu, Q.; Anenkhonov, O.A.; Chepinoga, V.V.; Autrey, B.C.; Liu, H. Comparing Pixel- and Object-Based Approaches in Effectively Classifying Wetland-Dominated Landscapes. Remote Sens. 2018, 10, 46. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qu, L.; Chen, Z.; Li, M.; Zhi, J.; Wang, H. Accuracy Improvements to Pixel-Based and Object-Based LULC Classification with Auxiliary Datasets from Google Earth Engine. Remote Sens. 2021, 13, 453. [Google Scholar] [CrossRef]
- Duro, D.C.; Franklin, S.E.; Dubé, M.G. A Comparison of Pixel-Based and Object-Based Image Analysis with Selected Machine Learning Algorithms for the Classification of Agricultural Landscapes Using SPOT-5 HRG Imagery. Remote Sens. Environ. 2012, 118, 259–272. [Google Scholar] [CrossRef]
- Rokach, L.; Maimon, O. Ensemble Methods for Classifiers. In Data Mining and Knowledge Discovery Handbook; Springer-Verlag: Boston, MA, USA, 2006; pp. 957–980. [Google Scholar]
- Berhane, T.M.; Lane, C.R.; Wu, Q.; Autrey, B.C.; Anenkhonov, O.A.; Chepinoga, V.V.; Liu, H. Decision-Tree, Rule-Based, and Random Forest Classification of High-Resolution Multispectral Imagery for Wetland Mapping and Inventory. Remote Sens. 2018, 10, 580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormann, C.F.; Elith, J.; Bacher, S.; Buchmann, C.; Carl, G.; Carré, G.; Marquéz, J.R.G.; Gruber, B.; Lafourcade, B.; Leitão, P.J.; et al. Collinearity: A Review of Methods to Deal with It and a Simulation Study Evaluating Their Performance. Ecography 2013, 36, 27–46. [Google Scholar] [CrossRef]
- Lillesand, T.; Kiefer, W.R.; Chipman, J. Remote Sensing and Image Interpretation, 7th ed.; John Wiley & Sons: Hoboken, NJ, USA, 2015; ISBN 0471026093. [Google Scholar]
- Janowski, L.; Tylmann, K.; Trzcinska, K.; Rudowski, S.; Tegowski, J. Exploration of Glacial Landforms by Object-Based Image Analysis and Spectral Parameters of Digital Elevation Model. IEEE Trans. Geosci. Remote Sens. 2022, 60, 1–17. [Google Scholar] [CrossRef]
- Breiman, L. Random Forests. Mach. Learn. 2001, 45, 5–32. [Google Scholar] [CrossRef] [Green Version]
- Biau, G.; Scornet, E. A Random Forest Guided Tour. Test 2016, 25, 197–227. [Google Scholar] [CrossRef] [Green Version]
- Belgiu, M.; Drăgu, L. Random Forest in Remote Sensing: A Review of Applications and Future Directions. ISPRS J. Photogramm. Remote Sens. 2016, 114, 24–31. [Google Scholar] [CrossRef]
- Smalheiser, N.R. Data Literacy: How to Make Your Experiments Robust and Reproducible, 1st ed.; Elsevier, Academic Press: London, UK, 2017; ISBN 9780128113066. [Google Scholar]
- Delincé, J. Handbook on Remote Sensing for Agricultural Statistics. In Handbook on Remote Sensing for Agricultural StatisticsAgricultural Statistics; Delincé, J., Ed.; GSARS Handbook: Rome, Italy, 2017. [Google Scholar]
- Zolyomi, B.; Fekete, G. The Pannonian Loess Steppe: Differentiation in Space and Time. Abstr. Bot. 1994, 18, 29–41. [Google Scholar]
- Team, R.C. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- TIBCO Software Inc. Statistica (Data Analysis Software System), Version 13. 2018. Available online: http://statistica.io (accessed on 4 March 2022).
- Bivand, R.; Gebhardt, A. Implementing Functions for Spatial Statistical Analysis Using the R Language. J. Geogr. Syst. 2000, 2, 307–317. [Google Scholar] [CrossRef]
- Sibaruddin, H.I.; Shafri, H.Z.M.; Pradhan, B.; Haron, N.A. Comparison of Pixel-Based and Object-Based Image Classification Techniques in Extracting Information from UAV Imagery Data. IOP Conf. Ser. Earth Environ. Sci. 2018, 169, 12098. [Google Scholar] [CrossRef]
- Agisoft, L. AgiSoft PhotoScan Professional (Version 1.2.6). 2016. Available online: http://www.agisoft.com/downloads/installer/ (accessed on 4 March 2022).
- Louhaichi, M.; Borman, M.M.; Johnson, D.E. Spatially located platform and aerial photography for documentation of grazing impacts on wheat. Geocarto Int. 2001, 16, 65–70. [Google Scholar] [CrossRef]
- Escadafal, R. Soil Spectral Properties and Their Relationships with Environmental Parameters—Examples from Arid Regions; Springer: Berlin/Heidelberg, Germany, 1994; pp. 71–87. [Google Scholar]
- Zarco-Tejada, P.J.; Miller, J.R.; Mohammed, G.H.; Noland, T.L.; Sampson, P.H. Estimation of chlorophyll fluorescence under natural illumination from hyperspectral data. Int. J. Appl. Earth Obs. Geoinf. 2002, 2001, 321–327. [Google Scholar] [CrossRef] [Green Version]
- Bannari, A.; Morin, D.; Bonn, F.; Huete, A.R. A review of vegetation indices. Remote Sens. Rev. 1995, 13, 95–120. [Google Scholar] [CrossRef]
- Travis, M.R.; Iverson, W.D.; Elsner, G.H.; Johnson, C.G. VIEWIT: Computation of Seen Areas, Slope and Aspect for Land-Use Planning; USDA Forest Service: Berkeley, CA, USA, 1975; Volume 11. [Google Scholar]
- Zevenbergen, L.W.; Thorne, C.R. Quantitative analysis of land surface topography. Earth Surf. Process. Landforms 1987, 12, 47–56. [Google Scholar] [CrossRef]
- Guisan, A.; Weiss, S.B.; Weiss, A.D. GLM versus CCA spatial modeling of plant species distribution. Plant Ecol. 1999, 143, 107–122. [Google Scholar] [CrossRef]
- Riley, S.J.; DeGloria, S.D.; Elliot, R. A Terrain Ruggedness Index that Qauntifies Topographic Heterogeneity. Intermt. J. Sci. 1999, 5, 23–27. [Google Scholar]
- Hengl, T.; Reuter, H.I. (Eds.) Geomorphometry: Concepts, Software, Applications; 1st ed.; Developments in Soil Science, Elsevier: Amsterdam, The Netherlands, 2009; Volume 33. [Google Scholar]
- QGIS Development, T. QGIS Geographic Information System: Open Source Geospatial Foundation Project; 2021. QGIS Association. Available online: http://qgis.osgeo.org (accessed on 4 March 2022).
- Hastie, T.; Tibshirani, R.; Friedman, J. The Elements of Statistical Learning, 2nd ed.; Springer: New York, NY, USA, 2017; Volume 27, ISBN 978-0-387-84857-0. [Google Scholar]
- Breiman, L.; Friedman, J.H.; Olshen, R.A.; Stone, C.J. Classification and Regression Trees, 1st ed.; Routledge: Boca Raton, FL, USA, 2017; ISBN 9781351460491. [Google Scholar]
- Xu, Y.; Goodacre, R. On Splitting Training and Validation Set: A Comparative Study of Cross-Validation, Bootstrap and Systematic Sampling for Estimating the Generalization Performance of Supervised Learning. J. Anal. Test. 2018, 2, 249–262. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J. Weighted Kappa: Nominal Scale Agreement Provision for Scaled Disagreement or Partial Credit. Psychol. Bull. 1968, 70, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Stokes, E.; Johnson, A.; Rao, M. Monitoring Wildlife Populations for Management. Available online: https://www.researchgate.net/profile/Arlyne-Johnson/publication/257363333_Module_7_Monitoring_Wildlife_Populations_for_Management_Background_Presentation_and_Exercises/links/00463525085e5ab4ca000000/Module-7-Monitoring-Wildlife-Populations-for-Management- (accessed on 17 June 2021).
- Stephenson, P.J. Integrating Remote Sensing into Wildlife Monitoring for Conservation. Environ. Conserv. 2019, 46, 181–183. [Google Scholar] [CrossRef]
- Plumptre, A.J. Monitoring Mammal Populations with Line Transect Techniques in African Forests. J. Appl. Ecol. 2000, 37, 356–368. [Google Scholar] [CrossRef]
- Agjee, N.H.; Mutanga, O.; Peerbhay, K.; Ismail, R. The Impact of Simulated Spectral Noise on Random Forest and Oblique Random Forest Classification Performance. J. Spectrosc. 2018, 2018, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Congalton, R.G. Accuracy Assessment and Validation of Remotely Sensed and Other Spatial Information. Int. J. Wildl. Fire 2001, 10, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Assal, T.J.; Lockwood, J.A. Utilizing Remote Sensing and GIS to Detect Prairie Dog Colonies. Rangel. Ecol. Manag. 2007, 60, 45–53. [Google Scholar] [CrossRef]
- Sherman, P.W. Kinship, Demography, and Belding’s Ground Squirrel Nepotism. Behav. Ecol. Sociobiol. 1981, 8, 251–259. [Google Scholar] [CrossRef]
- Weddell, B.J. Dispersion of Columbian Ground Squirrels (Spermophilus Columbianus) in Meadow Steppe and Coniferous Forest. J. Mammal. 1989, 70, 842–845. [Google Scholar] [CrossRef]
- Sequeira, A.M.M.; Bouchet, P.J.; Yates, K.L.; Mengersen, K.; Caley, M.J. Transferring Biodiversity Models for Conservation: Opportunities and Challenges. Methods Ecol. Evol. 2018, 9, 1250–1264. [Google Scholar] [CrossRef] [Green Version]
- Wenger, S.J.; Olden, J.D. Assessing Transferability of Ecological Models: An Underappreciated Aspect of Statistical Validation. Methods Ecol. Evol. 2012, 3, 260–267. [Google Scholar] [CrossRef]
- Jin, S.; Su, Y.; Gao, S.; Hu, T.; Liu, J.; Guo, Q. The Transferability of Random Forest in Canopy Height Estimation from Multi-Source Remote Sensing Data. Remote Sens. 2018, 10, 1183. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Cui, Q.; Sun, Y.; Wang, Q. Photovoltaic Panel Extraction from Very High-Resolution Aerial Imagery Using Region–Line Primitive Association Analysis and Template Matching. ISPRS J. Photogramm. Remote Sens. 2018, 141, 100–111. [Google Scholar] [CrossRef]
- Pontius, R.G.; Millones, M. Death to Kappa: Birth of Quantity Disagreement and Allocation Disagreement for Accuracy Assessment. Int. J. Remote Sens. 2011, 32, 4407–4429. [Google Scholar] [CrossRef]
- Wu, S.S.; Qiu, X.; Usery, E.L.; Wang, L. Using Geometrical, Textural, and Contextual Information of Land Parcels for Classification of Detailed Urban Land Use. Ann. Assoc. Am. Geogr. 2009, 99, 76–98. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Variable Acronym | Variable’s Long Name | Type | Reference or Description | Model |
|---|---|---|---|---|
| R | Red | Spectral | Red colour band | M1.1-2, M2.1-2 |
| G | Green | Spectral | Green colour band | M1.1-2, M2.1-2 |
| B | Blue | Spectral | Blue colour band | M1.1-2, M2.1-2 |
| GLI | Green Leaf Index | Spectral | [66] | M1.1-2, M2.1-2 |
| CI | Coloration Index | Spectral | [67] | M1.1-2, M2.1-2 |
| Intnsty | Intensity | Spectral | [67] | M1.1-2, M2.1-2 |
| NGRDI | Normalized Green/Red Difference Index | Spectral | [68] | M1.1-2, M2.1-2 |
| RI | Redness Index | Spectral | [69] | M1.1-2, M2.1-2 |
| SLP | Slope | Topographical | [70] | M1.1 |
| ASP | Aspect | Topographical | [71] | M1.1 |
| TPI | Topographioc Position Index | Topographical | [72] | M1.1 |
| TRI | Topographic Ruggedness Index | Topographical | [73] | M1.1 |
| Rghness | Roughness | Topographical | [73] | M1.1 |
| DEM | Digital Elevation Model | Topographical | [74] | M1.1, M2.1 |
| Grass height | Grass height | Topographical | Estimated height of surface point | M1.1 |
| Site | Study site | Environmental | Study site | M1.1 |
| Model 1.1 | Model 1.2 | Model 2.1 | Model 2.2 | |
|---|---|---|---|---|
| Overall Accuracy | 92.49 | 95.01 | 92.10 | 91.31 |
| Standard Deviation | 1.16 | 0.97 | 1.18 | 1.24 |
| CV% | 1.30 | 1.00 | 1.30 | 1.40 |
| 95% CI lower | 90.12 | 93.00 | 89.68 | 88.79 |
| 95% CI upper | 94.85 | 97.02 | 95.42 | 93.83 |
| Cohen’s Kappa | 0.76 | 0.84 | 0.76 | 0.74 |
| Standard Deviation | 0.04 | 0.03 | 0.04 | 0.04 |
| CV% | 4.6 | 3.6 | 4.6 | 5.0 |
| 95% CI lower | 0.69 | 0.78 | 0.69 | 0.66 |
| 95% CI upper | 0.83 | 0.90 | 0.83 | 0.81 |
| 0.90 | 1.00 | 1.00 | 0.95 | |
| Standard Deviation | 0.07 | 0.00 | 0.00 | 0.05 |
| CV% | 7.40 | 0.00 | 0.00 | 4.90 |
| 0.75 | 0.59 | 0.91 | 0.90 | |
| Standard Deviation | 0.09 | 0.10 | 0.06 | 0.07 |
| CV% | 11.70 | 16.80 | 6.70 | 8.10 |
| Iteration | Cohen’s Kappa for Precision | SD | CV% | Cohen’s Kappa for Sensitivity | SD | CV% | |
|---|---|---|---|---|---|---|---|
| Per-class (BURROW) | 1 | 0.96 | 0.04 | 4.30 | 0.85 | 0.07 | 8.00 |
| 2 | 0.82 | 0.06 | 7.10 | 0.95 | 0.04 | 3.80 | |
| 3 | 0.79 | 0.06 | 7.90 | 0.97 | 0.03 | 2.90 | |
| 4 | 1.00 | 0.00 | 0.00 | 0.84 | 0.06 | 7.20 | |
| 5 | 0.94 | 0.04 | 4.70 | 0.91 | 0.05 | 5.60 | |
| 6 | 0.97 | 0.03 | 3.30 | 0.94 | 0.04 | 4.50 | |
| 7 | 0.96 | 0.04 | 3.70 | 0.87 | 0.06 | 7.00 | |
| 8 | 0.97 | 0.03 | 3.40 | 0.85 | 0.06 | 7.10 | |
| 9 | 0.85 | 0.06 | 7.30 | 0.82 | 0.07 | 7.90 | |
| 10 | 1.00 | 0.00 | 0.00 | 0.90 | 0.05 | 5.80 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gedeon, C.I.; Árvai, M.; Szatmári, G.; Brevik, E.C.; Takáts, T.; Kovács, Z.A.; Mészáros, J. Identification and Counting of European Souslik Burrows from UAV Images by Pixel-Based Image Analysis and Random Forest Classification: A Simple, Semi-Automated, yet Accurate Method for Estimating Population Size. Remote Sens. 2022, 14, 2025. https://doi.org/10.3390/rs14092025
Gedeon CI, Árvai M, Szatmári G, Brevik EC, Takáts T, Kovács ZA, Mészáros J. Identification and Counting of European Souslik Burrows from UAV Images by Pixel-Based Image Analysis and Random Forest Classification: A Simple, Semi-Automated, yet Accurate Method for Estimating Population Size. Remote Sensing. 2022; 14(9):2025. https://doi.org/10.3390/rs14092025
Chicago/Turabian StyleGedeon, Csongor I., Mátyás Árvai, Gábor Szatmári, Eric C. Brevik, Tünde Takáts, Zsófia A. Kovács, and János Mészáros. 2022. "Identification and Counting of European Souslik Burrows from UAV Images by Pixel-Based Image Analysis and Random Forest Classification: A Simple, Semi-Automated, yet Accurate Method for Estimating Population Size" Remote Sensing 14, no. 9: 2025. https://doi.org/10.3390/rs14092025
APA StyleGedeon, C. I., Árvai, M., Szatmári, G., Brevik, E. C., Takáts, T., Kovács, Z. A., & Mészáros, J. (2022). Identification and Counting of European Souslik Burrows from UAV Images by Pixel-Based Image Analysis and Random Forest Classification: A Simple, Semi-Automated, yet Accurate Method for Estimating Population Size. Remote Sensing, 14(9), 2025. https://doi.org/10.3390/rs14092025